
果蝇Mef2基因多克隆抗体的制备及检测
2012-11-22 01:32:46徐腊梅盛立翔吴秀山莫小阳
湖南师范大学自然科学学报 2012年5期
朱 玲,徐腊梅,盛立翔,吴秀山,莫小阳
(湖南师范大学蛋白质化学及鱼类发育生物学教育部重点实验室,心脏发育研究中心,中国 长沙 410081)
Mef2属于MADS家族转录因子基因,MADS家族是指其蛋白质结构域含有高度保守的MADS结构域的一类转录因子的总称[1].MADS结构域是一种DNA结合结构域,由55~63个氨基酸组成,所识别并结合的DNA保守序列为CArG框[CC(A/T)6GG].
Mef2位于果蝇2号染色体上,mRNA全长5 990 bp,编码501个氨基酸.脊椎动物有4种MEF2蛋白(MEF2A-D),而果蝇中只有1种MEF2蛋白(D-MEF2)[2].不管是脊椎动物还是果蝇中的MEF2蛋白都含有一个高度保守的MADS和MEF2结构域,并且由它们来介导DNA结合、二聚作用、蛋白质-蛋白质的相互作用[3].
Mef2最初是在分化的肌肉细胞中作为一个肌肉特异结合的活性因子被鉴定的,1997 年却发现Mef2基因在心脏发育的过程中起重要作用[4].在果蝇中,Mef2首先在原肠胚形成时在整个中胚层中表达,随着胚胎的发育,它的表达逐渐限制在体节、内脏和心肌细胞中[5-6].鉴于Mef2基因在果蝇心脏发育的过程中起着重要的作用[7],所以制备一个特异性好的多克隆抗体尤其重要,对进一步以果蝇为模型研究Mef2基因具有重要的意义.
1 材料与方法
1.1 实验材料和试剂
大肠杆菌Rosseta菌种,pET-28a菌种以及coliDH5α菌种为本实验室提供;限制性内切酶EcoRⅠ和XhoⅠ,DNA、蛋白质相对分子质量Marker,Tag DNA聚合酶购自深圳晶美公司;pMD18-T载体和连接酶购自大连TaKaRa公司;UNIQ-10柱式DNA 胶回收纯化试剂盒购自上海生工公司;质粒提取试剂盒购自OMEGA公司;Glutathione SepharoseTM4B蛋白纯化试剂盒购自PIERCE公司;弗氏佐剂购自Sigma公司;丙烯酰胺、甲叉双丙烯酰胺、IPTG(异丙基-β-D-硫代半乳糖苷)等购自上海Sangon公司;新西兰大白兔购自中南大学实验动物学部.
1.2 引物设计与合成
从NCBI数据库(http://www.ncbi.nlm.nih.gov)检索出果蝇Mef2基因的序列,根据巢式PCR的原理利用Primer Premier 5.0 软件设计两对引物:第一对:D-Mef2-S1:5′CGCCAGCTGCCGGAAATCCA 3′;D-Mef2-A1:5′GCGTCTGCAGCGTGACCACT 3′.第二对(分别加入EcoRⅠ和XhoⅠ酶切位点):D-Mef2-S2:5′GAGAATTCATGGGCCGCAAA AAAATTCA 3′EcoRⅠ;D-Mef2-A2:5′GGCTCGAGGTGGACTGGCCTGCAGCAGG 3′XhoⅠ.
1.3 基因克隆和载体构建
选取30只野生型果蝇,液氮冷冻30 min,用Trizol法提取总RNA[8],反转录PCR扩增,获得first-chain cDNA.然后以first-chain cDNA为模板用第一对引物进行PCR扩增,然后以所得产物为模板,用第二对引物进行PCR扩增,得到目的片段[9].目的片段经纯化后克隆入T载体,转化入DH5α感受态细胞中,酶切检测筛选出阳性单克隆,经测序鉴定后将DNA片段从pMD18-T-Mef2质粒上切下,连入pET-28a载体(EcoRⅠ和XhoⅠ酶切线性化),得到阳性克隆,经EcoRⅠ和XhoⅠ双酶切鉴定后,得到重组表达质粒pET-28a-Mef2.
1.4 Mef2融合蛋白诱导表达及纯化
pET-28a-Mef2重组表达质粒转化入大肠杆菌菌株Rosseta,筛选出阳性菌落,然后接种于LB 培养基(含100 mg/L 氯霉素,卡那霉素)37 ℃ 培养12 h左右,以1∶50转接扩大培养至OD600达0.6,按0.1 mol/L 加IPTG,25 ℃诱导,4、5、6、7 h各取1 mL菌液,确定最佳诱导时间.
收集诱导后大肠杆菌,PBS漂洗后超声裂解,离心取上清.4 ℃下与经Binding Buffer漂洗活化后的Ni-IDA凝胶柱结合,Washing Buffer洗去杂蛋白,Elution Buffer洗脱目的蛋白,获得纯化的His-Mef2融合蛋白,-80 ℃保存备用.
1.5 Mef2多克隆抗体制备
将纯化后的His-Mef2蛋白与弗氏完全佐剂按体积1∶1在注射器中推成乳剂,分散约12个点对同一新西兰大白兔进行背部皮下免疫注射.在第一次注射后14、21、28 d,将His-Mef2蛋白按体积比1∶1与弗氏不完全佐剂在注射器中推成乳剂,进行多次免疫.第35天主动脉取血,静置过夜(4 ℃),3 000 r/min离心处理10 min,取上清分装保存(-80 ℃).
1.6 效价测定
获得免疫兔血清后,用诱导的总蛋白进行Western blot实验检测抗体效价.将含pET-28a-Mef2重组子的菌株在LB培养液中37 ℃培养,将5 mL接种于200 mL新鲜培养基中,用IPTG在25 ℃条件下诱导5 h后,收集细菌,PBS重悬,然后用SDS-PAGE电泳分离.将Mef2抗体分别以1∶100、1∶500、1∶1 000和1∶2 000的体积比稀释后,以免疫前兔血清作为对照,采用Western blotting方法检测抗体的特异性[10].
2 结果与分析
2.1 重组质粒pET-28a-Mef2的构建与鉴定
将纯化后的果蝇Mef2基因片段克隆到pMD18-T载体中,经测序鉴定与基因理论序列100%匹配,然后用EcoRⅠ和XhoⅠ双酶切,连入pET-28a载体(EcoRⅠ和XhoⅠ酶切线性化),获得重组质粒pET-28a-Mef2.经酶切和测序鉴定重组质粒正确(图1).
2.2 Mef2 融合蛋白诱导表达及纯化
将重组子转入Rosseta感受态细胞中,以0.1 mmol/L IPTG进行诱导,30 ℃条件下进行时间梯度诱导1,2,3,4,5,6 h分别取样检测蛋白的表达,结果(如图2)显示Mef2蛋白在诱导1 h后就开始表达,随着时间推移表达量增加,在3 h后趋于稳定.
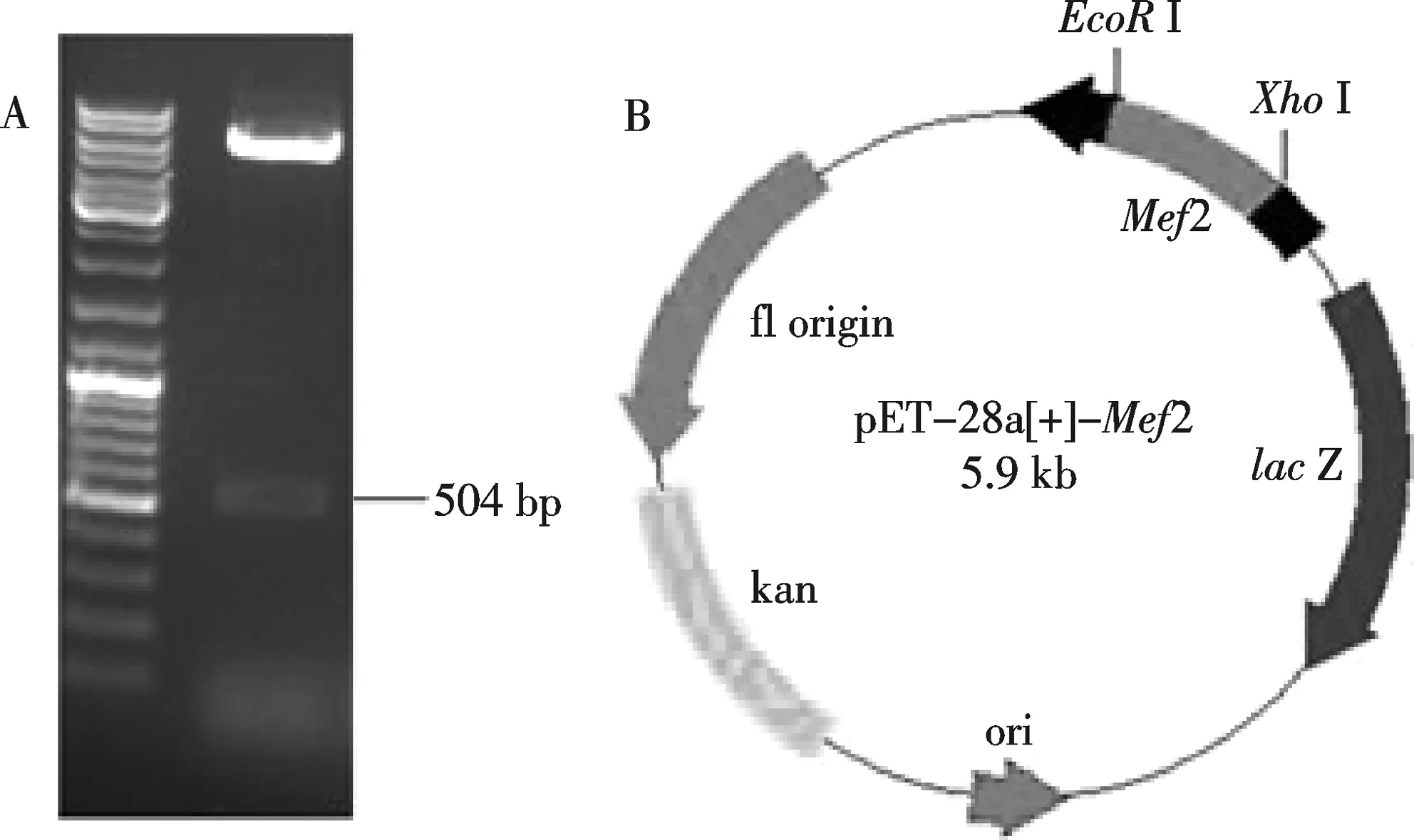
A: pET-28a-Mef2质粒双酶切鉴定,B:pET-28a-Mef2质粒示意图图1 pET-28a-Mef2质粒酶切鉴定图谱和示意图

His-Mef2融合蛋白时间梯度诱导, IPTG 0.1 mmol/L, 温度30 ℃. 1:Marker; 2:pET-28a-Mef2诱导前;3~8:0.1 mmol/L IPTG诱导pET-28a-Mef2 蛋白表达1~6 h.图2 His-Mef2 融合蛋白的表达
选定最佳诱导条件(30 ℃、0.1 mmol/L IPTG 浓度、3 h),大量诱导获得浓度较高的蛋白,超声裂解后经Ni-IDA凝胶柱亲和纯化,获得比较纯的蛋白(图3),可以进行蛋白免疫[11].
2.3 抗血清的制备与效价检测
用纯化的目的蛋白免疫新西兰大白兔获得血清,用诱导的总蛋白进行效价测定,抗体稀释体积比分别为1∶100、1∶500、1∶1 000和1∶2 000,免疫前兔血清(control)为对照.结果发现抗体的效价较好,满足实验所需 (图4).

图3 His-Mef2 融合蛋白的纯化
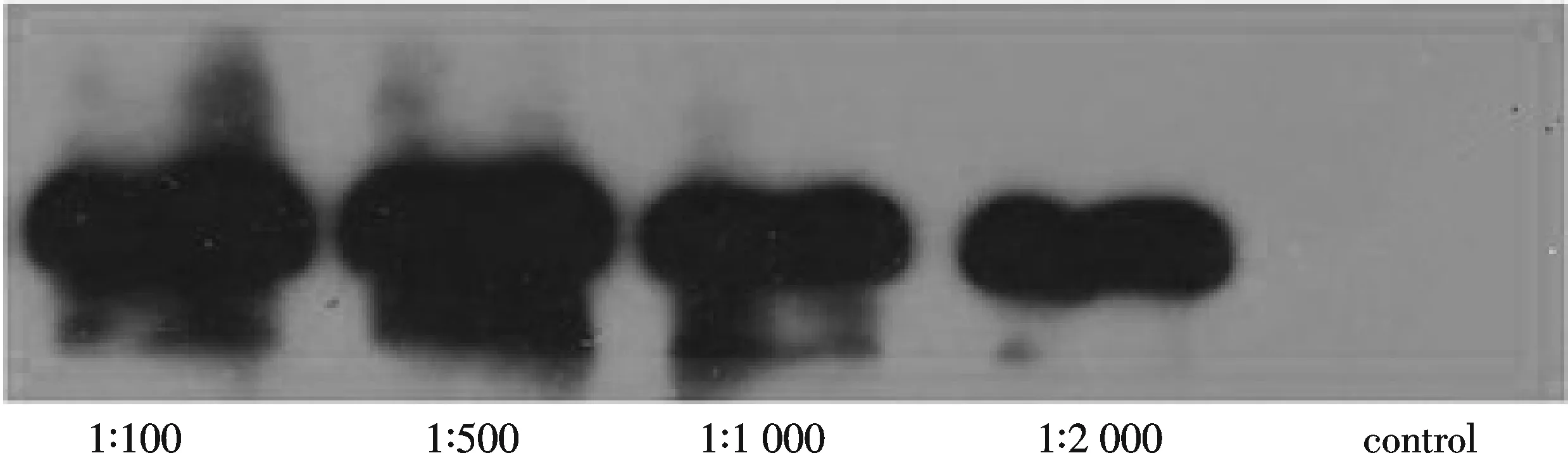
图4 Mef2 抗体效价测定
3 讨论
为了以果蝇为模型进一步研究Mef2基因的功能,本文通过试验获得了Mef2原核表达融合蛋白,制备了高效价和特异性好的兔抗Mef2血清.在获得融合蛋白的过程中,先通过软件分析选取了亲水性好而又含结构域少的一段序列,为获得特异性好的抗体提供了前提条件[12].在诱导蛋白的过程中,IPTG 的终浓度选取的是0.1 mmol/L ,蛋白表达量相对高,又不至于因IPTG 浓度太高对细胞的生长产生毒性作用,试验相对容易成功.
pET-28a(+)载体是一种常用的原核表达载体,具有Kan抗性,质粒大小为5.3 kb.载体的N端携带His标签、凝血酶基因及T7启动子标签序列;在C端具有His标签序列, pET-28a(+)载体可在BL21(DE3)、Rosstea等表达菌株中高表达融合有His标签的蛋白,便于下游蛋白纯化.pET-28a这种载体没有本身溶解性高的多肽融合蛋白,也没有催化二硫键形成的酶融合蛋白,而且不含信号肽序列,所以同一种蛋白用这个载体,形成包涵体的可能性更大些[13].
本试验中,作者通过构建pET-28a-Mef2重组质粒、诱导表达His-Mef2融合蛋白获得了高纯度的蛋白并免疫新西兰大白兔制备了Mef2抗体.通过效价检测证明制备的Mef2多克隆抗体具有较好的特异性,能满足试验要求. 因此,它为将来运用染色质免疫沉淀、免疫共沉淀、免疫组化等手段深入研究Mef2基因的功能奠定了基础.
参考文献:
[1] SOLER C, HAN J, TAYLOR M V,etal.The conserved transcription factor Mef2 has multiple roles in adult Drosophila musculature formation[J]. Development, 2012,139(7):1270-1275.
[2] MILLIGAN J N, JOLLY E R. Identification and characterization of a Mef2 transcriptional activator in schistosome parasites.[J]. PloS Negl Trop Dis, 2012,6(1):e1443.
[3] FLAVELL S W, COWAN C W, KIM T K,etal. Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number[J]. Science, 2006,311(5763):1008-1012.
[4] BLANCHARD F J, COLLINS B, CYRAN S A,etal. The transcription factor Mef2 is required for normal circadian behavior in Drosophila[J]. J Neurosci, 2010,30(17):5855-5865.
[5] 吴秀山,袁婺洲,王跃群,等.心脏发育研究[M].长沙:湖南科学技术出版社, 2004:214-302.
[6] 吴秀山.心脏发育概论[M]. 北京:科学出版社, 2006:86-103.
[7] WESSELLS R J, BODMER R. Age-related cardiac deterioration: insights from drosophila [J]. Biosci, 2007,12:39-48.
[8] WU X, GOLDEN K, BODMER R,etal. Heart development in drosophila requires the segement polarity genewingless [J]. Dev Biol, 1996,177:104-116.
[9] LI S, ZHOU D Y, LU M M,etal. Advanced cardiac morphogenesis does not require heart tube fusion[J].Science, 2004,305(5690):1619-1622.
[10] 龚 琳,吴秀山,李永青,等.果蝇Nulp1特异性多克隆抗体的制备及检测[J].湖南师范大学自然科学学报, 2011,34(4):69-73.
[11] 闵 璐, 周云雷, 周 煌,等.lrrc10蛋白的表达、纯化及多克隆抗体的制备[J].湖南师范大学自然科学学报, 2011,34(6):68-71.
[12] 刘华友,朱 玲,李 帆,等.斑马鱼Foxp4基因多克隆抗体的制备及检测[J].激光生物学报, 2011,20(4):556-559.
[13] 梁 艳,宋怀婷,李竞超,等.myh6蛋白的表达、纯化及多克隆抗体制备[J].激光生物学报, 2010,19(5):63-67.
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
大自然探索(2023年11期)2023-03-01 09:04:36
环球时报(2022-09-20)2022-09-20 15:18:57
学苑创造·A版(2022年3期)2022-03-29 23:32:16
今日农业(2020年24期)2020-12-15 16:16:00
学苑创造·A版(2019年6期)2019-07-11 01:07:39
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
兽医导刊(2016年12期)2016-05-17 03:51:50
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
