
黔南晚石炭世生物灰泥丘生长发育特征研究
2012-11-14 02:38屠锡艳刘永华张晓光
中国非金属矿工业导刊 2012年6期
屠锡艳,李 明,刘永华,李 芃,张晓光
(中国建筑材料工业地质勘查中心辽宁总队,辽宁 沈阳 110004)
1 引言
生物礁研究一直是地质科学领域中十分重要的课题。尽管关于生物礁的概念说法不一,但总体认识是:生物礁是由各种生物建造而成的碳酸盐岩隆,具有原地形成的各种生物结构,代表着生物原始的生存位置,反映生物原始的生态格局[1-3]。正是由于生物礁的这一独特的生物建造特点,让我们可以借助对生物礁的研究,来恢复其地史时期中生物群落组成、结构、生长发育过程及古生态特征。生物礁是开展沉积学、古生物学、古生态学等学科研究的理想场所。生物礁作为独特的生物建造,其礁内各种孔隙、空洞非常发育,是石油和天然气理想的储集场所。自上世纪20年代,由于在生物礁中发现了大量的油气,更加大了人们对生物礁的研究力度,另外生物礁与层控矿床也有密切关系,因此,生物礁研究得到普遍关注和重视。
从各地史时期生物礁的研究现状来看,石炭纪生物礁的研究一直比较薄弱,特别是我国更是如此。多数学者认为石炭纪是地史时期的造礁低潮期,其礁体数量少,类型单一。但是,近几年以巩恩普教授为代表的学者对于石炭纪生物礁的研究取得了突破,证实了石炭纪存在大量不同类型的生物礁。
通过对工作区的野外和室内工作,基本查明了黔南晚石炭世生物灰泥丘生长发育特征,通过系统总结和分析,并且对比Waulsortian相研究,笔者认为晚石炭世生物灰泥丘并不是Waulsortian相,这对中国其他地区广泛开展石炭纪生物灰泥丘研究具有重要指导意义,有助于石炭纪生物礁研究工作的深入。
2 区域地质特征
黔南地区大地构造位置归属于滇黔桂盆地,又称南盘江盆地(图1)。该盆地作为扬子板块西南被动陆缘的一部分,从泥盆纪开始出现裂谷化成因的裂陷槽,因大陆拉伸,在南盘江盆地中出现堑垒相间的构造格局[1]。石炭纪时,该盆地与特提斯海盆邻接,基底活动性强。由于深部热膨胀引起以南北方向为主的引张力作用,造成地壳微型扩张,形成一系列隆、凹相间构造格局。在扩张过程中,处于固结较早、刚性较强的构造高位发育成碳酸盐台地,而拉张下陷部位则演化成台间深水盆地。随裂陷加深,构成了浅水补偿型碳酸盐台地与半深水非补偿型台沟相间的古构造和岩相展布格局[2]。
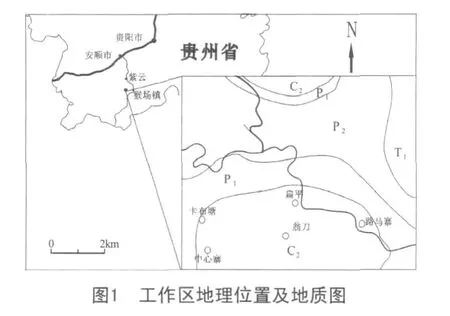
黔南晚石炭世海域进一步扩大,浅水碳酸盐台地范围扩大并连成一片,台盆相已收缩成孤立的狭长状[3]。沉积岩石类型为独山—威宁分区浅水台地相浅色碳酸盐岩,主要有浅灰色—灰色中厚层至块状生物碎屑灰岩、泥晶灰岩、微晶灰岩和各种礁灰岩,含有丰富的底栖生物。
3 泥丘相地层生物地层学研究
3.1 泥丘相剖面
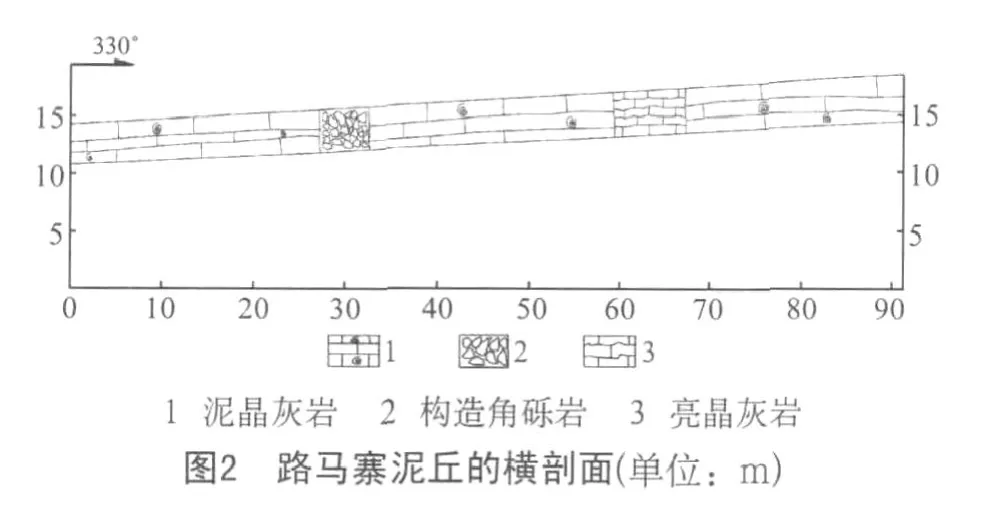
路马寨泥丘横剖面(见图2)长90m,主要为泥晶胶结的粒泥灰岩。地层中生物量很少,偶见海百合茎及有孔虫,但蜓碎屑相对较发育。在泥丘的横剖面可以看出,路马寨灰泥丘在横向上岩性变化小,只发育一段构造角砾岩和一段亮晶灰岩,这两段岩性差异主要来源于构造运动以及成岩后期的亮晶方解石充填孔隙、重结晶作用。
(1)泥丘下伏岩层——蜓生物碎屑灰岩。该层灰岩中发育大量的蜓碎屑,偶见海百合茎、有孔虫、单体珊瑚以及少量腕足动物碎片。
(2)生物灰泥丘,外貌均呈穹形隆起。丘核均为泥晶灰岩,不显层理,均呈块状,且藻丝发育,偶见团块状粒状体(推测为藻颗粒)。丘顶与丘核无明显差异。丘两翼均为生物碎屑灰岩,主要为蜓碎屑灰岩。
(3)灰泥丘的上覆岩层——含蜓的亮晶生物碎屑灰岩。该层灰岩中发育大量的蜓碎屑、海百合茎、腕足动物、单体珊瑚以及其他生物。岩石以亮晶胶结为主,属高能环境产物。
3.2 礁相地层生物群特征
从上述各礁相地层剖面看,工作区广泛发育一套灰色碳酸盐岩,其中礁、丘、滩分布普遍,代表了黔南晚石炭世礁相地层典型的岩性特点。在地层中,生物繁盛,其中不乏多种造礁生物,为黔南晚石炭世生物礁的形成和发展奠定了基础。
蜓是石炭系和二叠系划分、对比地层的重要标准化石,在本工作区所代表的黔南晚石炭世地层中,它是地层划分、对比的主要标准。
工作区产出生物灰泥丘地层中,蜓类化石相对其它生物化石,则较为丰富,从总的蜓类生物群特征来看,发育灰泥丘的地层应该是晚石炭世晚期地层。
3.3 泥丘中的其他生物
工作区泥丘相地层中除上述生物外,还发育其他一些生物。它们有的具有造丘作用,有的具有附丘作用,还有的具有毁丘作用,以下对于有关各种生物的特征及其在泥丘中的作用进行简单论述。
(1)有孔虫。有孔虫在灰泥丘中非常少见,且种属单一,不起造丘作用,呈单体存在。
(2)海百合茎。泥丘相地层中分布一些海百合茎,个体较小,呈小的群体产出。
(3)藻类。泥丘相地层中存在的藻类较单一,具体属种鉴定比较困难,绝大多数缺少内部结构特征。经观察研究推测其应为蠕孔藻。
(4)蓝细菌。在礁相地层中起粘结作用,形成粘结岩,也是形成规模不同生物泥丘的造丘生物。
4 生物灰泥丘特征
黔南晚石炭世生物灰泥丘为较深水的,通过微生物粘结作用、造岩作用形成的碳酸盐建隆,其基本特征见图3。
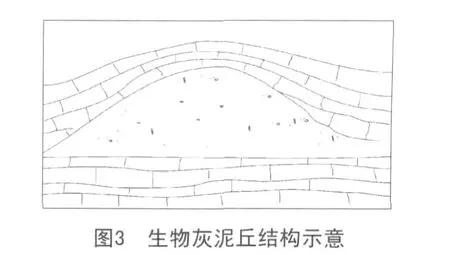
(1)组构特征。外貌均呈穹形隆起,抗浪构造明显而典型。丘核均为泥晶灰岩,不显层理,且藻丝发育,偶见团块状粒状体(推测为藻颗粒)。丘顶与丘核无明显差异。丘两翼均为生物碎屑灰岩,主要为蜓碎屑灰岩。野外露头和显微镜下观察均可见晶洞,孔内为亮晶方解石充填。
(2)岩石构成。其岩石构成主要为粒泥灰岩,及少量藻凝块岩,但骨骸颗粒较低而泥晶基质含量较高,表明为较深水的,水动力条件较弱的沉积环境。
(3)古生物化石及其古生态。同层位的滩相沉积富含多种生物,腕足类、蜓、非蜓有孔虫、海百合、苔藓虫以及珊瑚类,与此形成鲜明对比的是,灰泥丘的生物丰度很低且种属单调。偶见单体珊瑚化石,薄片中见少量海百合、有孔虫以及蜓类化石。
丘翼主要由亮晶胶结生物碎屑灰岩组成,砂砾屑成分为泥晶方解石,普遍富含藻丝;重结晶作用强,镜下常见重结晶方解石呈嵌晶状,将砂屑、生物屑镶嵌其中。亮晶方解石明显可分为两期:第一期为沿孔壁生长的马牙状方解石;第二期呈中一粗粒方解石充填孔隙中,在粒状方解石晶间孔中偶见残余沥青。由于强烈的胶结作用和重结晶作用,残余原生孔隙基本全被两期方解石胶结物填充。次生孔隙,以砂屑,生物碎屑粒内溶孔和粒间溶孔、方解石胶结物晶间溶孔为主,孔隙度很低。在灰泥丘丘顶与丘核分布有藻丝。丘顶部与围岩接触界限明确。
灰泥丘的生长发育过程:由于凝块石灰泥丘内无宏观的造架生物,因而长期以来对其成因有多种认识。灰泥丘夹于灰色—灰黑色含化石丰富的灰岩之中。经过观察以及较为深入的研究,将工作区灰泥丘的生长发育过程分为三个阶段。
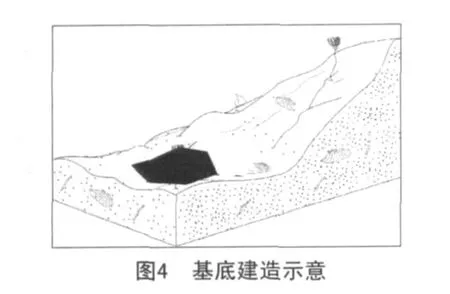
第一阶段:基底的建立(发育期)。
当水深、水温、水动力条件等控制灰泥丘生长的因素达到适合其生长的标准时,蓝细菌等微生物开始在较为平坦的碎屑滩上大量繁殖,在生长过程中一方面粘结灰泥,另一方面钙化的微生物参与造岩,同时也是灰泥的提供者。在各种微生物的共同作用下,形成了具早期固结特点简单的基底建造(见图4)。
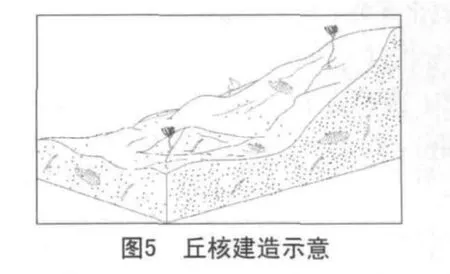
第二阶段:丘核的建立(繁盛期)。
在简单的基底建造之上,蓝细菌等微生物进一步繁殖生长,形成了生物灰泥丘的次级生长构造。在开放的格架孔中,可充填灰泥和细小生物碎屑等沉积物,而在遮闭的格架孔中,空洞可被亮晶方解石充填,或者可继续发育成为凝块石。蓝细菌群落在这种比较安静的水体中生活,灰泥含量较高。群落内蓝细菌是优势生物,多为丝状体,在其生长过程中,它以粘结作用和粘结包覆作用来粘结或吸附灰泥颗粒一同沉积下来,进而完成泥丘建造(见图5)。
第三阶段:丘顶的建立(衰退期)。
灰泥丘的终结有两点原因(见图6)。
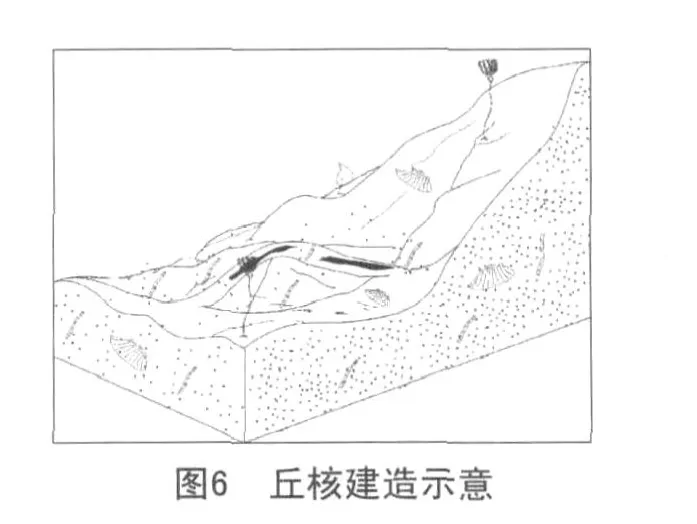
(1)随着海平面的下降,水动力条件的增强,大量的造礁生物大量繁殖,汲取了大部分的养料,微生物在这种动荡的沉积环境中很难生存,钙化停止进而被在水动力作用较强的条件下沉淀的颗粒质灰岩掩埋—灰泥丘终结。
(2)随着海平面的上升,水动力条件的减弱,蓝细菌赖以生存的阳光很难到达海底,水温也降低。由于缺少阳光以及温度的影响,蓝细菌(自养)不能完成正常的新陈代谢而走向死亡—灰泥丘终结。
综上所述,微生物的沉积、造岩造丘作用明显,灰泥丘的发育主要是微生物生命活动的结果。
5 黔南生物灰泥丘与世界Waulortian丘对比研究
(1)规模对比。就丘体的规模而言,黔南生物灰泥丘的厚度和直径从几米到几十米且成群出现,一般规模较小;而Waulsortian丘的厚度和直径从一米到上百米,成群出现或横向联结形成上千米长的大泥滩,一般规模较大。
(2)岩相对比。就岩相方面比较,黔南生物灰泥丘的岩相组成为粒泥灰岩,属弱水动力条件的沉积环境;Waulsortian丘的岩相组成粒泥灰岩、泥粒灰岩、颗粒灰岩、漂浮岩,岩石颗粒大于黔南生物灰泥丘,说明Waulsortian丘生成环境的水动力条件略强于黔南生物灰泥丘的生成水动力条件。
(3)造丘生物对比。黔南生物灰泥丘的造丘生物为自养细菌,其生活环境应为富氧、透光的环境;Waulsortian丘的造丘生物为异养细菌,其生活环境应为富养、不透光或亚透光的环境。说明Waulsortian丘的生成水深要大于黔南生物灰泥丘的生成水深。
(4)丘内常见生物对比。黔南生物灰泥丘内常见的有孔虫、海百合茎、苔藓虫以及蜓不起造丘作用,只是悬浮于灰泥基质中的一种附属品。且这些化石的个体都很小,呈小的单体,内部结构简单;Waulsortian丘的常见的珊瑚、钙质海绵、有孔虫、腕足类、投足类、海百合茎以及苔藓虫不起造丘作用,只是悬浮于灰泥基质中的一种附属品。大部分珊瑚和海绵呈指状或层状,藉以适应泥质环境。此外,这些珊瑚化石的个体都很小,呈小的单体,隔壁发育,间室构造简单,地层跨度长。但Waulsortian丘这些生物量很大,远大于在黔南灰泥丘中的含量。在通过对比说明,黔南生物灰泥丘和Waulsortian丘的生存环境均不适合其它生物的生长。
(5)成丘地层对比。黔南生物灰泥丘通过蜓类鉴定,确定其生成地层为晚石炭世地层;而Waulsortian丘的生成地层为早石炭世(杜内期—韦宪期)的地层。Waulsortian丘早于黔南生物灰泥丘形成。
(6)晶洞对比。黔南生物灰泥丘和Waulsortian丘都遍存在洞穴(层状和条带状孔洞),代表软泥中的流体排泄构造,部分或全部充以微亮晶或亮晶方解石胶结物。
(7)丘体结构对比。黔南生物灰泥丘和Waulsortian丘都具备丘顶、两翼、丘核的结构,但Waulsortian丘的三结构具有明显的岩相差异。①海百合的停机坪构造(海百合丘顶):该停机坪有4个优势相,即颗粒灰岩、粒泥灰岩、泥粒灰岩及漂浮岩相,孔隙被方解石充填。该相缺乏其他类型的浅成灰岩,整个构造近乎水平;②苔藓虫丘核(丘核):岩石为粗集粒灰岩和漂浮岩,其中含有具有窗孔构造的苔藓虫及少量的海百合茎。丘核主要为泥晶胶结,含有海百合茎、窗孔苔藓虫、有孔虫、腕足类等生物,生物种属单一,个体小且数量少,地层跨度长;③两翼(丘翼):其岩石为粒泥灰岩、泥粒灰岩及漂浮岩,含有少量的海百合及介形虫,具有纤细的泥质迭片结构,坡角为30~50°;而对于黔南生物灰泥丘的三结构不具备明显的岩相差异,丘顶和丘核主要为泥晶胶结的粒泥灰岩和微晶灰岩,两翼为泥晶胶结的蜓碎屑灰岩。
(8)成岩作用对比。Waulsortian丘的成岩作用包括:方解石胶结、机械压实作用、白云岩化、硅化以及碳氢岩化。丘核岩相包括水底束状的方解石、叶片状/凌柱状方解石团块具有明亮的光谱特征,孔隙被亮晶方解石充填。海百合停机坪构造包含共轴胶结物与海百合有序胶结,即4种白云岩化的类型被认知:①局部置换;②化学压实;③白云岩浸透;④氨基转换。
对于黔南生物灰泥丘的成岩作用较为简单,主要为重结晶、胶结作用。白云岩化、硅化作用在泥丘成岩过程中并未起主要作用,只是在后期的构造裂隙中偶然发生且不强烈。证明泥丘被埋藏后受热液影响较小。
(9)沉积环境对比。根据以上分析确定黔南生物灰泥丘的沉积环境为较深水、富氧、弱水动力条件的透光带斜坡上;而Waulsortian丘的沉积环境深水、贫氧、弱水动力条件的亚透光带或不透光斜坡上。
综上所述,黔南地区所发育的灰泥丘与Waulsortian丘具有明显的差异,因此确定黔南地区所发育的灰泥丘并非Waulsortian丘,而是一种特殊的石炭纪灰泥建造。
6 结论
(1)灰泥丘的类型单一,规模小。
(2)造礁生物为自养的蓝细菌微生物。
(3)丘内生物种类单一,内部结构简单。
(4)查明了生物灰泥丘的建造过程。
(5)确定生物灰泥丘的生成环境为较深水、富氧、弱水动力条件的透光带斜坡上。
(6)确定了生物灰泥丘的产丘层位为上石炭统上部地层,即马平组下部地层。
(7)与Waulsortian相对比研究表明,黔南地区所发育的灰泥丘与Waulsortian丘具有明显的差异,因此确定黔南地区所发育的灰泥丘并非Waulsortian相。
中国南方晚石炭世生物灰泥丘研究并不多,除本工作区之外,其他地区还没有开展相关研究。所以生物灰泥丘的研究对全面认识晚石炭世生物礁的基本发育特征具有重要意义。
[1]赵焕庭,宋朝景,朱袁智.南沙群岛“危险地带”腹地珊瑚礁的地貌与现代沉积特征[J].第四纪研究,1992(4):368-377.
[2]赵美霞,余克服,张乔民.珊瑚礁区的生物多样性及其生态功能[J].生态学报,2006,26(1):186-194.
[3]杨惠民,刘炳温,邓宗淮.滇黔桂海相碳酸盐岩地区最佳油气保存单元的评价与选择[M].贵阳:贵州科技出版社,1999,44-47.
猜你喜欢
山东冶金(2022年2期)2022-08-08
矿产综合利用(2021年3期)2021-07-14
同煤科技(2021年1期)2021-03-25
中国海上油气(2020年5期)2020-10-20
矿产综合利用(2020年1期)2020-07-24
科学导报·学术(2019年21期)2019-09-10
——黔南示范小城镇集锦(之二)
当代贵州(2017年50期)2017-12-19
夜郎文学(2017年2期)2017-09-26
当代贵州(2017年16期)2017-06-07
当代贵州(2016年29期)2016-11-03
