
马鞍列岛海洋特别保护区岩相潮间带底栖生物初步研究
2012-11-06 09:31金敬林蔡丽萍吴盈子
海洋开发与管理 2012年11期
金敬林,蔡丽萍,吴盈子
(国家海洋局舟山海洋工作站 舟山 316021)
马鞍列岛海洋特别保护区岩相潮间带底栖生物初步研究
金敬林,蔡丽萍,吴盈子
(国家海洋局舟山海洋工作站 舟山 316021)
2010年9月的大潮期间在马鞍列岛海洋特别保护区岩相潮间带共设两个断面,进行底栖生物调查。共鉴定出潮间带底栖动、植物21种,其中软体动物11种,甲壳动物5种,藻类4种,环节动物1种。用多样性指数、均匀度指数、丰富度指数及ABC曲线对岩相潮间带底栖生物群落进行分析,结果显示:潮间带底栖生物多样性指数较低,两断面的底栖生物未受到干扰。
岩相;潮间带;底栖生物
潮间带是位于高低潮之间的地带,即高潮时被水淹没,低潮时露出水面的浅水地带。潮间带为湿地生态系统的一个重要类型,生物资源丰富,底栖生物是潮间带生态系统的重要组成部分。近几年由于人类生产开发活动频繁,且海陆理化因子交替作用下环境复杂多变,对潮间带生态学的研究得到了国内外学者的广泛关注。许多学者对舟山海域潮间带底栖生物有了一定的研究[1-6],但对马鞍列岛海洋特别保护区岩相潮间带底栖生物未做调查,本研究希望通过对马鞍列岛海洋特别保护区岩相潮间带底栖生物分布的类型、数量等特征进行分析,为马鞍列岛海洋特别保护区开发利用总体规划的制定、综合管理、环境保护和合理利用等提供基础数据和科学依据,实现马鞍列岛海洋特别保护区资源保护与开发的和谐统一。
1 材料与方法
1.1 调查区域概况
2005年6月,马鞍列岛海洋特别保护区正式批准建立。马鞍列岛海洋特别保护区位于长江口外东南,泗礁岛东北部,保护区范围呈方形,总面积549 km2,其中岛陆面积19 km2,海域面积530 km2。主要保护对象为重要经济鱼类、贝藻类以及无居民海岛岛礁资源。
1.2 采样站位和方法
本次调查时间为2010年9月,分别在马鞍列岛海洋特别保护区的嵊山岛和西绿华岛各设1处断面(图1)。

图1 潮间带采样断面
每条断面各设3个点,对高潮区、中潮区和低潮区分别进行定性和定量采集。所布设的断面为岩礁开放型,远离人为及污染源的地方。高潮区因生物分布不均,选用25 cm×25 cm的定量框取样,中潮区和低潮区的生物量比较密集,故选用10 cm×10 cm的定量框取样。在采集定量样品的同时,将其附近的所有底栖生物种类收入,作为定性样品,并分别用甲醛进行现场固定,带回实验室分析鉴定。
采集操作过程、室内样品的计数、称重、分析计算、资料整理均严格依据《海洋监测规范》中第七部分近海污染生态调查和生物监测的规定进行[7]。
1.3 数据处理与分析
按照《海洋监测规范》中第七部分近海污染生态调查和生物监测的规定[7],对调查的岩相潮间带底栖生物评价分析。计算公式如下:
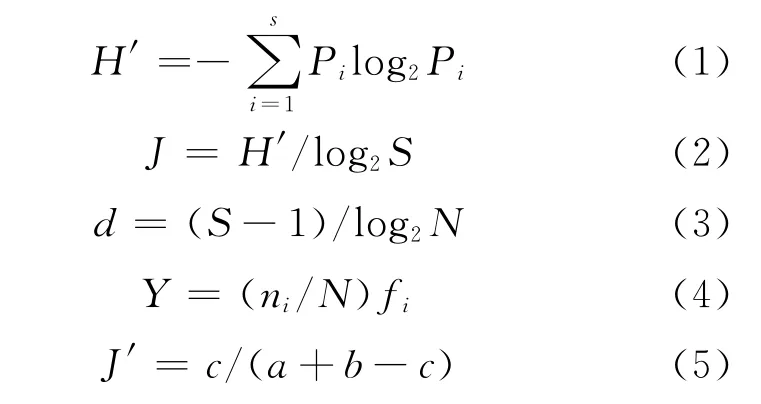
式中:H′为香农-韦弗(Shannon-Weaver)多样性指数;J为皮诺(Pielou)均匀度指数;d为马卡列夫(Margalef)丰富度指数;Y为优势度;J′为Jaccar群落系数;N为样品中的生物总个体数;S为样品中的种类总数;Pi为种i的个体数占总个体数的比例;fi为该种出现的站位数占总站位数之比的百分比;ni为第i种的个体数;a、b分别为两条断面的种类数,c为两条断面的共有种数。并用丰富度/生物量比较曲线,即ABC曲线(abundance and biomass curves)来监测环境污染对潮间带底栖动物群落的扰动。
2 结果与分析
2.1 种类与组成
此次调查出现的岩相潮间带底栖生物共鉴定出21种,隶属4门,两断面均以软体动物为主。主要有粒结节滨螺、短滨螺、日本笠藤壶、厚壳贻贝、鳞笠藤壶等。其中软体动物11种,占52.4%;甲壳动物5种,占23.8%;藻类4种,占19.0%;环节动物1种,占4.8%。
西绿华岛断面共鉴定出岩相潮间带底栖生物17种,其中软体动物9种,占52.9%;甲壳动物4种,占23.5%;藻类3种,占17.7%;环节动物1种,占5.9%。
嵊山岛断面共鉴定出岩相潮间带底栖生物16种,其中软体动物9种,占56.3%;甲壳动物5种,占31.2%;藻类2种,占12.5%。
2.1.1 生物种类水平分布
西绿华岛和嵊山岛这两断面岩相潮间带底栖生物种类水平分布存在一定的差异(图2)。西绿华岛共鉴定出17种,嵊山岛共鉴定出16种。在西绿华岛断面出现了环节动物。
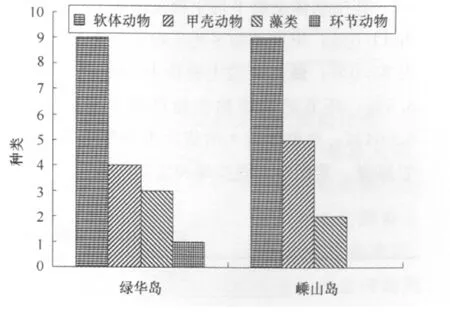
图2 底栖生物不同断面的物种组成
2.1.2 生物种类垂直分布
西绿华岛和嵊山岛岩相潮间带底栖生物垂直分布明显,集中分布在中潮区和低潮区,高潮区生物分布较少(表1)。各潮区分带明显,高潮区为滨螺带,生物有短滨螺、粒结节滨螺;中潮区为藤壶带,生物以鳞笠藤壶和厚壳贻贝为主;低潮区为藤壶-藻类带,生物有鳞笠藤壶和鼠尾藻等。
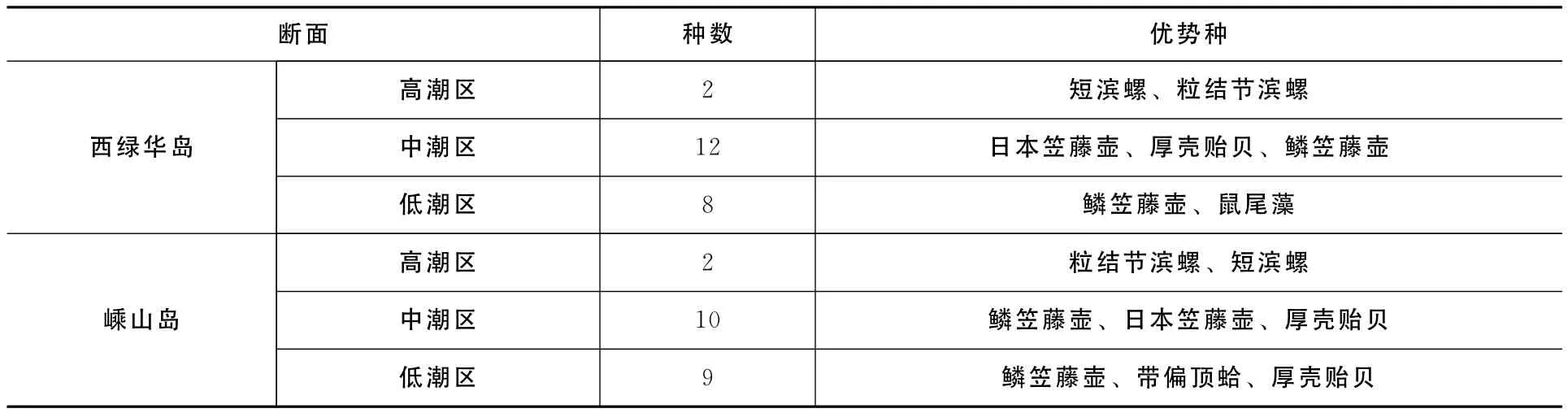
表1 西绿华岛和嵊山岛两断面不同潮区底栖生物的优势种分布
2.2 潮间带底栖生物密度和生物量
2.2.1 潮间带底栖生物平均栖息密度和生物量的水平分布
西绿华岛断面的平均密度为4 033个/m2,其中软体动物平均密度为2 283个/m2,占56.6%;甲壳动物平均密度为1 733个/m2,占43.0%;环节动物平均密度为17个/m2,占0.4%;西绿华岛潮间带底栖生物主要以软体动物为主,其次为甲壳动物;栖息密度较大的依次为粒结节滨螺、鳞笠藤壶、短滨螺、日本笠藤壶和疣荔枝螺。
西绿华岛断面的平均生物量为11 062.1 g/m2,其中软体动物平均生物量为1 275.8 g/m2,占11.5%;甲壳动物平均生物量为9 399 g/m2,占85.0%;藻类平均生物量为386.8 g/m2,占3.5%;环节动物平均生物量为0.5 g/m2,占0.004%;生物量较大的依次为鳞笠藤壶、日本笠藤壶、紫贻贝、短滨螺和疣荔枝螺。
嵊山岛断面的平均密度为8 217个/m2,其中软体动物平均密度为5 117个/m2,占62.3%;甲壳动物平均密度为3 100个/m2,占37.7%。嵊山岛潮间带底栖生物也主要以软体动物为主,其中栖息密度较大的依次为粒结节滨螺、日本笠藤壶、紫贻贝、鳞笠藤壶和厚壳贻贝。
嵊山岛断面的平均生物量为16 856 g/m2,其中软体动物平均生物量为7 126 g/m2,占42.3%;甲壳动物平均生物量为9 678 g/m2,占57.4%;藻类平均生物量为52 g/m2,占0.3%。生物量较大的依次为日本笠藤壶、鳞笠藤壶、厚壳贻贝、紫贻贝和带偏顶蛤。
2.2.2 潮间带底栖生物平均栖息密度和生物量的垂直分布
潮间带底栖生物平均栖息密度和平均生物量垂直分布见表2。由表2可知,平均栖息密度和平均生物量在中潮区和低潮区比较大,由大到小依次为中潮区、低潮区、高潮区。

表2 潮间带底栖生物平均栖息密度和平均生物垂直分布
2.3 物种丰富度、均匀度和群落多样性
通过调查,对马鞍列岛海洋特别保护区潮间带底栖生物的物种丰富度指数(d)、均匀度指数(J)和香农-韦弗多样性指数(H′)进行分析(表3)。从表3可以看出,西绿华岛和嵊山岛这两个断面的岩相潮间带底栖生物物种丰富度、均匀度和多样性指数差别较大,且多样性指数均较低。
种类组成是群落最基本的特征,可以反映生物群落和环境的相互关系,不同生境栖息的生物种类和组成各不相同。西绿华岛和嵊山岛这两个断面的共有物种数为12种,用Jaccard群落系数J′得出,这两个断面的群落系数J′=0.571 4,种类构成差别较大,说明这两个断面生境存在一定的不同。
2.4 ABC曲线分析
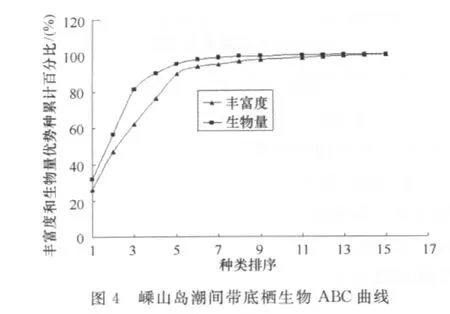
海洋大型底栖生物生活相对稳定(固定底内或短距离活动),对海洋环境污染反应敏感,具有污染效应指示性。Warwick提出用丰度/生物量比较曲线(ABC曲线)法来监测环境污染对大型底栖生物群落的扰动,并对不同海域、不同生境中的大型底栖生物群落进行了验证,结果表明:对于任何物理性、生物性以及污染扰动引起的大型底栖生物群落变化,ABC曲线都能灵敏地作出反应[8]。根据调查这两个断面底栖生物的丰富度和生物量的数据做ABC曲线(图3和图4)。由图3和图4可知,两断面潮间带底栖生物的ABC曲线中生物量曲线位于丰富度曲线之上。

表3 西绿华岛和嵊山岛岩相潮间带底栖生物生态学参数

3 讨论
3.1 岩相潮间带底栖生物平均生物量和密度分析
通过对马鞍列岛海洋特别保护区岩相潮间带底栖生物种类平均生物量和密度的分析,与其他海区的调查数据比较(表4),结果表明:马鞍列岛海洋特别保护区岩相潮间带底栖生物的平均生物量大于其他海区,平均密度除略小于普陀中街山保护区外,也均大于其他海区。由此可以看出,嵊泗马鞍列岛保护区岩相潮间带底栖生物保护较好。
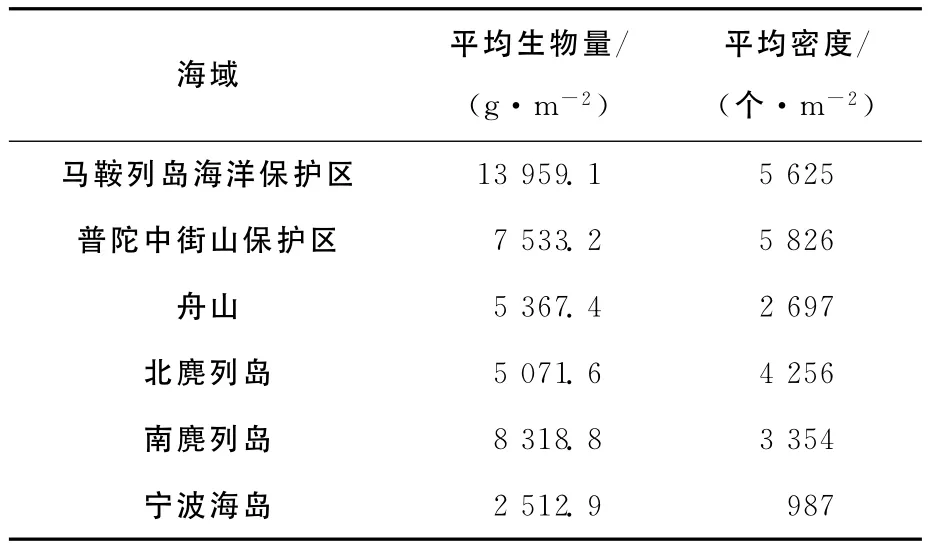
表4 马鞍列岛保护区岩相潮间带与其他地区的平均生物量和密度比较[2,9-12]
3.2 环境污染问题
对马鞍列岛海洋特别保护区岩相潮间带底栖生物进行ABC曲线分析表明,这两个断面的岩相潮间带底栖生物未受到干扰。但是2010年该保护区水质为劣Ⅳ类海水水质标准,主要超标因子为无机氮[13],这应当引起相关部门的注意。
3.3 物种南移
在定性样品调查中,发现了翡翠贻贝。该物种是低潮区至潮下带17 m范围内的养殖品种,主要分布在福建、广东和海南[14],翡翠贻贝在马鞍列岛海洋特别保护区的出现,可能由全球气候变暖引起,也可能是由外来船舶带到该海域。
4 结论与建议
通过调查,马鞍列岛海洋特别保护区岩相潮间带底栖生物的平均生物量和平均密度较大,但是底栖生物种类单一,主要以软体动物和甲壳动物为主,且潮间带经济软体贝类不断衰退,生物个体呈小型化,而一些不能食用或者可食性较小的生物种群却得到充分生长繁衍,从而在群落中占了较大的比例,如粒结节滨螺和短滨螺。建议当地政府加大对马鞍列岛保护区的保护力度,合理开发海洋资源,促进海岛经济和自然环境和谐可持续发展。
[1] 廖一波,曾江宁,陈全震,等.嵊泗海岛不同底质潮间带春秋季大型底栖动物的群落格局[J].动物学报,2007,53(6):1000-1010.
[2] 朱四喜,章飞军,顾沈明,等.中街山列岛岩相潮间带大型底栖动物的群落格局[J].生态科学,2009,28(5):391-397.
[3] 章飞军,丁宏印,邱树萍,等.浙江秀山岛潮间带大型底栖动物群落组成及其生物多样性[J].浙江海洋学院学报:自然科学版,2010,29(1):9-14.
[4] 杨万喜,陈永寿.嵊泗列岛潮间带群落生态学研究Ⅱ:岩相潮间带底栖动物的群落结构[J].应用生态学报,1998,9(1):75-78.
[5] 宋翔,朱四喜,杨红丽,等.浙江岱山岛潮间带大型底栖动物的群落结构[J].浙江海洋学院学报:自然科学版,2009,28(2):214-218.
[6] 寿鹿,高爱根,曾江宁,等.底质环境对浙江衢山岛潮间带大型底栖动物分布的影响[J].动物学杂志,2007,42(3):79-83.
[7] 中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会.GB 17378.7—2007海洋监测规范(近海污染生态调查和生物监测)[S].北京:中国标准出版社,2008.
[8] 顾晓英,陶磊,尤仲杰,等.象山港大型底栖动物群落特征[J].海洋与湖沼,2010,41(2):208-213.
[9] 蔡如星,郑锋,王彝豪,等.舟山潮间带生态学研究II:数量组成与分布[J].东海海洋,1990,9(3):58-72.
[10]张永普,应雪萍,吴海龙,等.北麂列岛岩相潮间带底栖生物群落的组成特征[J].海洋湖沼通报,2000(4):23-33.
[11]彭欣,谢起浪,陈少波,等.南麂列岛潮间带底栖生物时空分布及其对人类活动的响应[J].海洋与湖沼,2009,40(5):584-589.
[12]卢建平,蔡如星,胡建云.宁波海岛潮间带生态学研究II:数量组成与分布[J].东海海洋,1996,14(4):57-66.
[13]2010年舟山市海洋环境公报[R].舟山市:舟山市海洋渔业局,2011.
[14]黄宗国.中国海洋生物种类与分布[M].北京:海洋出版社,2008.
猜你喜欢
海洋信息技术与应用(2020年4期)2021-01-18
海洋通报(2020年3期)2020-10-20
——全球首台3000吨超级浆态床锻焊加氢反应器成功登陆鱼山岛
大陆桥视野(2020年6期)2020-07-02
东坡赤壁诗词(2019年4期)2019-09-12
石油化工建设(2018年1期)2018-07-10
快乐语文(2018年13期)2018-06-11
东南文化(2016年4期)2016-08-22
应用海洋学学报(2015年1期)2015-11-22
中学生天地(A版)(2015年7期)2015-08-06
读者(乡土人文版)(2013年10期)2013-04-12
