
模拟酸雨对台湾桤木光合特性的影响
2012-09-29 07:34:12黄桂芬吴承祯林勇明
亚热带农业研究 2012年4期
黄桂芬,吴承祯,洪 伟,林 晗,洪 滔,林勇明,陈 灿,李 键
(1.漳州市南靖县龙山镇人民政府,福建 南靖 363600;2.福建省高校森林生态系统过程与经营重点实验室,福建 福州 350002;3.福建农林大学林学院,福建 福州 350002)
近年来,随着全球经济的发展,酸雨已成为世界三大环境问题之一。我国大面积的土地已遭受酸雨危害,是全球范围内的第三大酸雨区,仅次于欧洲和北美。随着植物种类、发育阶段以及酸雨pH值的变化,酸雨对植物的光合作用会产生不同的影响[1-3]。酸雨通过降低植物叶绿素含量、组成及破坏同化组织降低植物的光合作用[4],且随着酸度的增加,影响愈来愈严重。周青等[5]、陈美华等[6]研究发现,当酸雨pH为2.5时,腊梅、芒果的光合速率显著降低;单运峰等[7]研究认为,pH 3.0的酸雨明显降低马尾松和杉木的净光合速率,但当pH<3.0时,净光合速率未发生明显变化。Yu et al[8]研究表明,酸雨使得黄瓜幼苗的净光合速率显著下降,主要是由非气孔因素引起,并且可变荧光与最大荧光的比值(Fv/Fm)也随之下降。单运峰[9]研究表明,随着模拟酸雨酸度的降低,杉木、马尾松、水杉、擦木、火力楠、青冈和油茶7种植物的叶绿素含量有所上升。Shna et al[10]认为酸雨导致松树叶绿素含量增加。酸雨通过影响叶绿素含量而影响植物光合作用,随着叶绿素含量的减少和叶绿素a/b的变化,植物的光合作用受到了直接的影响。
台湾桤木(Alnus formosana)为桦木科桤木属落叶大乔木,原产台湾,在原产地分布较为广泛,生态适应性强,生长快,由于具有固氮菌能生物固氮,培肥林地土壤。台湾桤木已在福建、江西、湖南等地引种,前人针对台湾桤木分布及种源和家系选择[11]、地理变异和种源区划[12]、混交林营造[13]、桤木引种[14]、早期适应性[15]、桤木生物量[16]、林分结构规律[17]、水分胁迫的生理响应[18]、水分胁迫的光合响应和抗旱性[19]、细根生长及土壤性质变化特征[20]、蒸腾特性及影响因子[21]等开展了系统研究,但有关酸雨对台湾桤木光合特性的影响未见报道。为此,本研究探讨不同浓度酸雨对台湾桤木幼苗光合荧光特征的影响,以期为台湾桤木的栽培和引种推广提供依据。
1 材料与方法
1.1 试验地概况
试验地位于福建农林大学森林生态研究所田间试验地,北纬25°15'-26°39',东经118°08'-120°37',属亚热带海洋性季风气候,年均气温16-20℃,1月均温10.5℃ ,7月均温28.7℃,年降水量900-2100 mm,平均日照数1700-1980 h,无霜期326 d,平均湿度77%,气候温和,雨量充沛,四季常青。
1.2 试验设计
2010年3月从南平来舟林业试验场取1年生台湾桤木幼苗,移植入直径25 cm的盆中。每盆装园土10 kg,种植1棵台湾桤木幼苗,放入大棚中,防止雨水对其影响。台湾桤木苗高度约为100 cm,常规培养3个月后,选取长势良好且基本一致的幼苗,于2010年6月14日开始喷洒酸溶液,模拟5种不同浓度的酸雨胁迫。用稀 H2SO4配制酸雨,酸雨 pH 值分别设为 2.5、3.0、3.5、4.0、4.5,以蒸馏水为对照(CK),每个处理设5个重复。用小型喷雾器将不同浓度的酸液均匀喷于整株台湾桤木幼苗上,每3天喷洒1次,每株幼苗每次喷洒50 mL。气温过高时可喷点水,以保证台湾桤木幼苗的正常生长。酸雨处理30 d后,取主茎上的成熟叶进行各光合指标的测定,每隔30 d测定1次。所得数据用EXCEL软件和DPS软件处理。
1.3 光合参数的测定
从2010年7月中旬开始观测,于7月14日、8月11日、9月8日和10月6日上午9:00-11:00,采用便携式光合仪对台湾桤木主茎上成熟叶片进行光合指标测定,每个处理重复3次。测定时保持外界正常的温度、湿度和CO2浓度。测试指标包括:叶片的净光合速率(Pn)、蒸腾速率(Tr)、细胞间隙CO2浓度(Ci)、气孔导度(Gs)等。气孔限制值(Ls)采用Berry et al[22]的方法进行计算。WUE为水分利用效率,Ca为空气中CO2浓度。

2 结果与分析
2.1 酸雨对台湾桤木叶片Pn的影响
不同酸雨酸度胁迫1个月后(7月),台湾桤木叶片的Pn随酸雨酸度的降低而升高。pH 2.5时,叶片的Pn达到最低点,比CK降低28.6%;pH 4.5时,Pn达到最高点,比CK高出10.2%。台湾桤木叶片Pn随胁迫时间的延长呈先降低后升高的趋势,说明台湾桤木对酸雨胁迫的反应比较敏感,使得Pn降低,但是随着对外界环境的逐渐适应,受伤害的细胞逐渐得到恢复,叶片Pn呈现上升的趋势(图1)。
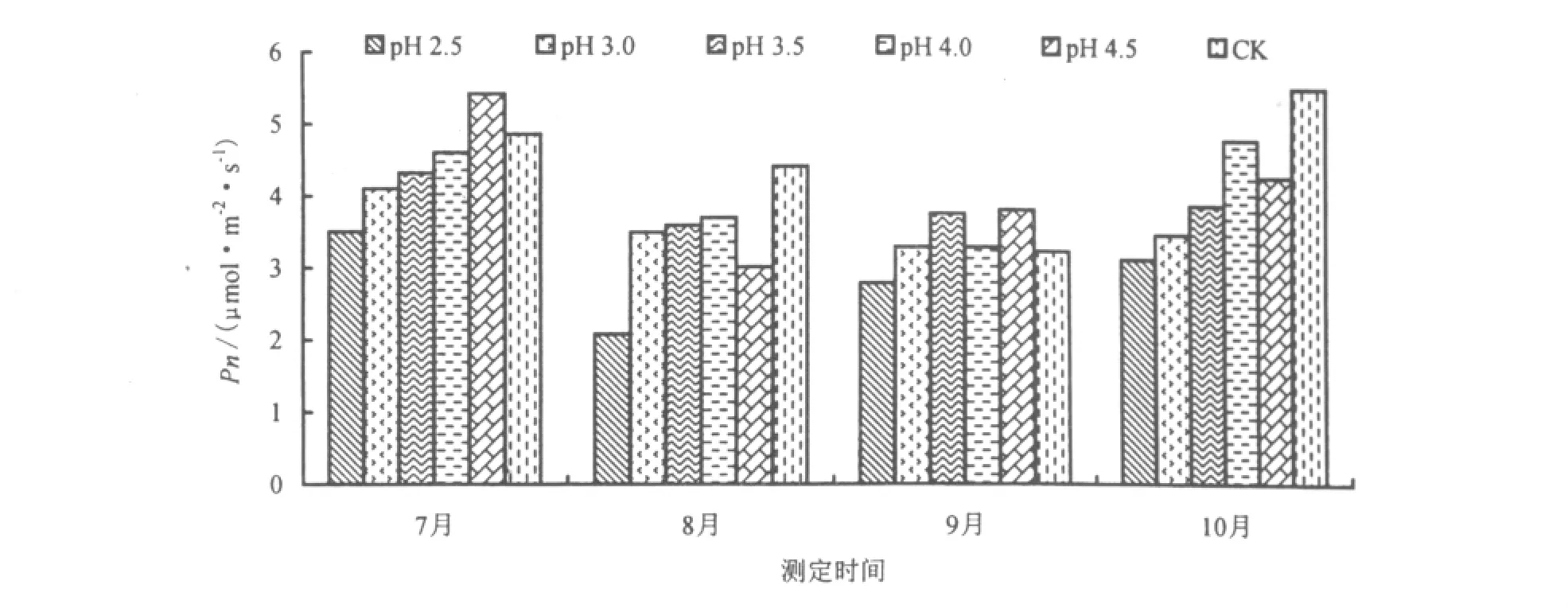
图1 酸雨胁迫对Pn的影响Fig.1 Effect of acid rain stress on the net photosynthetic rate
酸雨的影响使得植物的叶绿素合成受到阻碍,叶绿素含量和叶绿素a/b含量降低,从而影响植物的光合作用,降低了植物的 Pn。吕均良等[23]研究表明,经过 pH 3.5、2.5、2.0 的酸雨处理后,桃梨叶片的 Pn 降低,与本研究的结论相符。
2.2 酸雨对台湾桤木叶片Tr及WUE的影响
酸雨胁迫1个月后(7月),台湾桤木叶片的Tr和WUE均随酸雨酸度的降低而升高。pH 2.5时,叶片的 Tr达到最低点,比 CK 降低 59.5%,而 pH 3.0、3.5、4.0、4.5 分别比 CK 降低 57.4%、53.1%、50.1% 和34.7%(图2)。随着酸雨胁迫时间的延长,台湾桤木叶片Tr呈先升高后降低的趋势。经过pH 2.5、3.0、3.5、4.0 及 4.5 处理的台湾桤木叶片 WUE 比 CK 分别降低 25.9%、26.5%、25.4%、16.6% 和 32.6%。酸雨降低了台湾桤木叶片的WUE,与红花木莲、乐东拟单性木兰、金叶含笑等植物趋势一致[24]。随着酸雨胁迫时间的延长,台湾桤木叶片WUE呈先降低后升高的趋势(图3)。
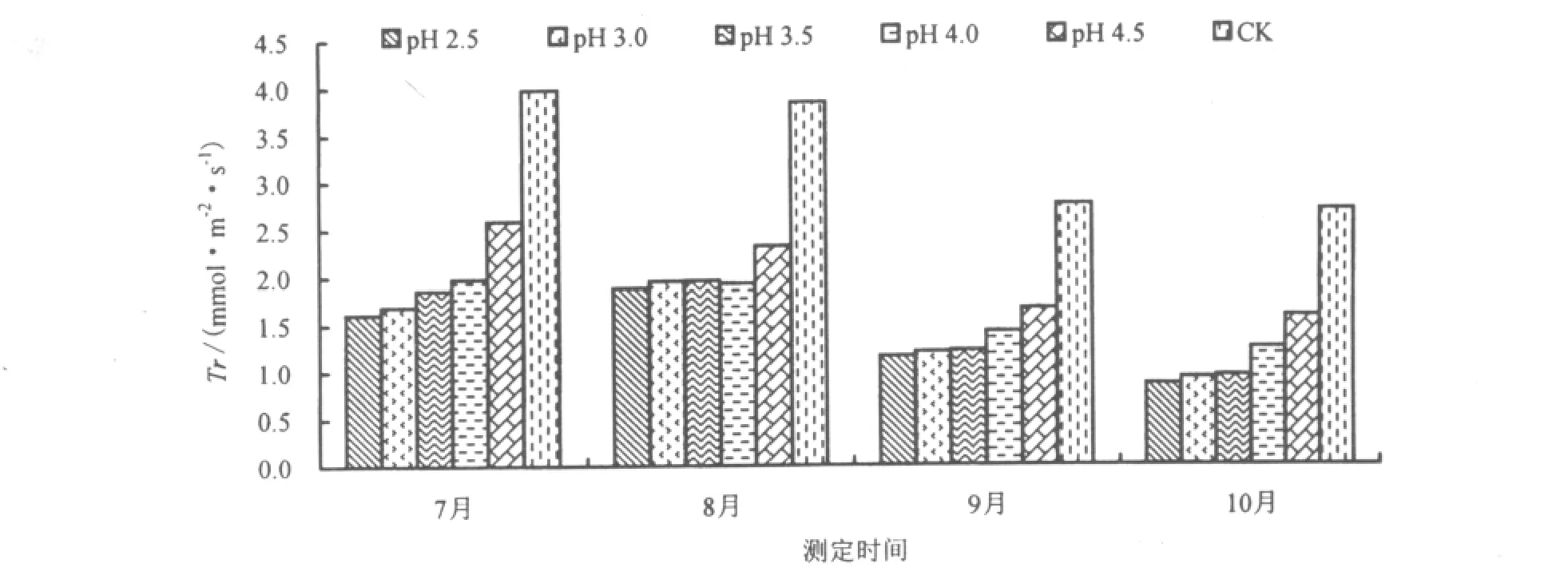
图2 酸雨胁迫对Tr的影响Fig.2 Effect of acid rain stress on the transpiration rate
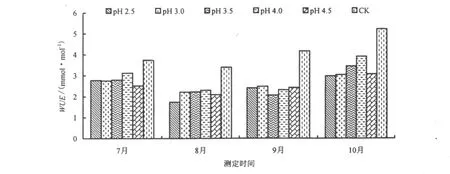
图3 酸雨胁迫对WUE的影响Fig.3 Effect of acid rain stress on the water use efficiency
福州7-8月气温比较高,水、热条件充沛,生态系统内外潜热交换强烈,植物处于旺盛生长阶段,Tr很大。9-10月,随着气温的降低和酸雨胁迫的持续,Tr逐渐降低。Tr受到光合有效辐射、气温、相对湿度、土壤水势、Gs、Ci以及叶面水汽压亏损等多种因子的影响而发生变化[25,26],这些因子对台湾桤木Tr的影响程度存在差异。朱万泽等[27]研究表明,光合有效辐射、Gs、空气温差和叶面水汽压亏缺是影响台湾桤木叶片蒸腾作用的主要生理生态因子。酸雨减少光合有效辐射、降低Gs而导致Tr下降。
2.3 酸雨对台湾桤木叶片Ci及Gs的影响
酸雨胁迫1个月后(7月),台湾桤木叶片的Ci随酸雨酸度的降低而降低。pH 2.5时,叶片的Ci达到最高点,比CK高出4.1%,pH 4.0时,叶片的Ci达到最低点,比CK降低0.9%,其他处理的Ci都比CK高(图4)。由此可见,酸雨一定程度上提高了台湾桤木Ci,与李志国等[24]研究红花木莲等6种园林植物趋势相一致。随着酸雨胁迫时间的延长,台湾桤木叶片Ci呈前期降低,至9月达最低,10月缓慢升高的趋势。
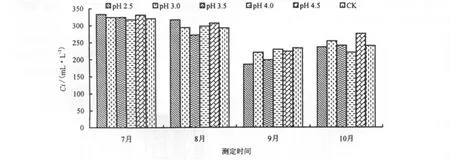
图4 酸雨胁迫对Ci的影响Fig.4 Effect of acid rain stress on the intercellular CO2concentration
植物气孔可以防止植物因为蒸腾作用所引起的水份过分流失,从而保证植物能够获得足够的CO2气体进行光合作用。酸雨胁迫1个月后(7月),台湾桤木叶片的Gs随着酸雨酸度的降低而升高,这与李志国等[24]研究红花木莲等6种园林植物对酸雨的响应规律一致。在各处理中,pH 2.5时,叶片的Gs达到最低点,比CK降低22.1%;pH 4.5时,叶片的Gs达到了最高点,比CK高出6.87%(图5)。在酸雨的作用下,台湾桤木Gs降低是为了适应酸雨胁迫环境,降低蒸腾,从而减少水份的损失。随着酸雨胁迫时间的变化,台湾桤木叶片Gs呈先降低后升高的趋势。
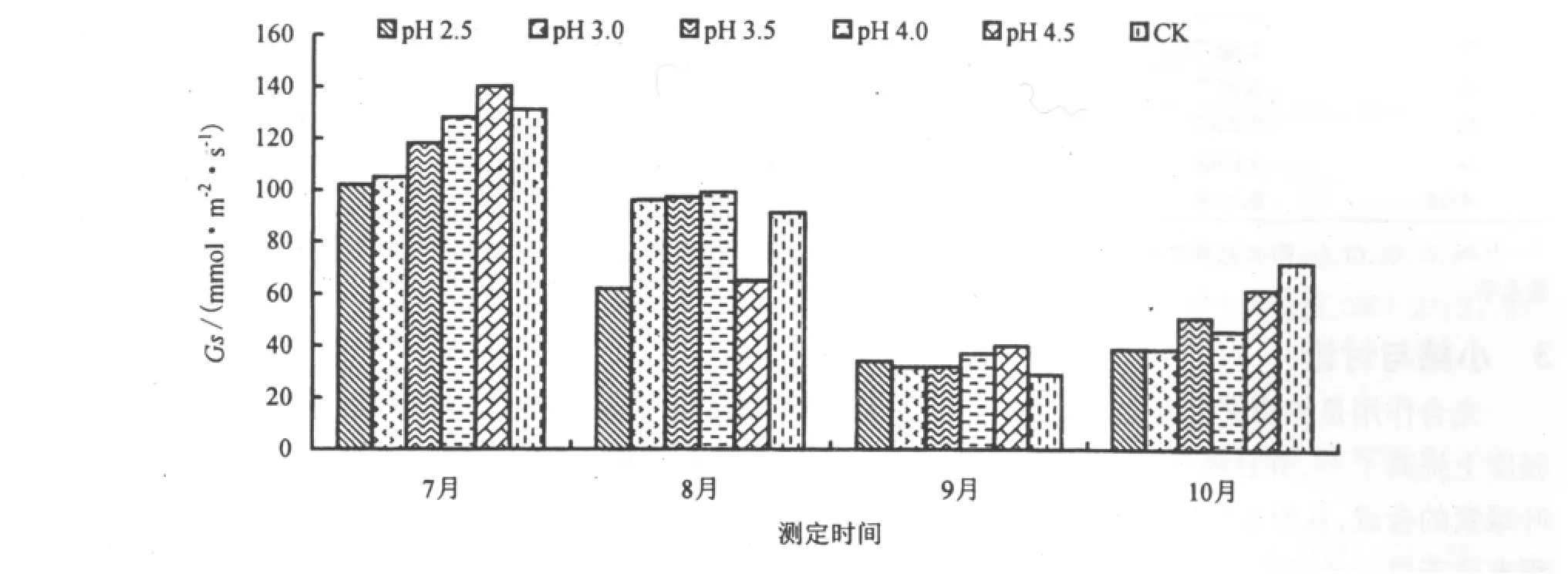
图5 酸雨胁迫对Gs的影响Fig.5 Effect of acid rain stress on the stomatal conductance
2.4 酸雨对台湾桤木叶片Ls的影响
酸雨胁迫1个月后(7月),台湾桤木叶片的Ls随酸雨酸度的降低而升高,各处理的Ls均低于CK,与李志国等[24]研究红花木莲等6种园林植物叶片Ls对酸雨的响应规律一致。pH 4.5时,叶片的Ls最低,比CK 降低 37.1%。pH 2.5、3.0、3.5、4.0 的 Ls分别比 CK 降低 25%、26.5%、26.9% 和 15.2%(图6)。随着酸雨胁迫时间的延长,台湾桤木叶片Ls呈先升高后降低的趋势。酸雨降低台湾桤木叶片Ls,Farquhar et al[28]认为,当叶内Ci减少、Ls增大时,气孔的部分关闭成为光合下降的主要原因,而当叶内Ci增加、Ls减少时,非气孔限制值成为光合下降的主要原因。本研究中由于酸雨的影响,Ci增大,而Ls减少,表明非气孔限制值是台湾桤木光合下降的主要原因。
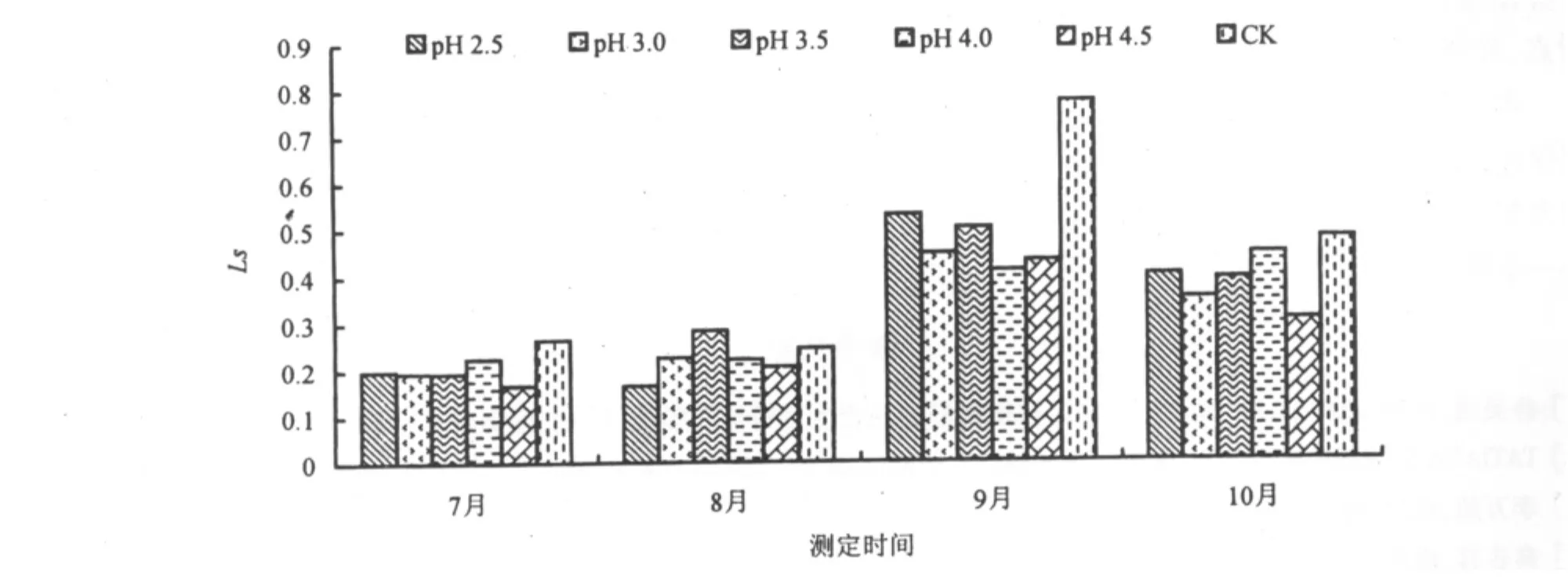
图6 酸雨胁迫对Ls的影响Fig.6 Effect of acid rain stress on the stomatal limitation
2.5 酸雨胁迫下台湾桤木叶片各光合指标间的相关性分析
台湾桤木幼苗叶片Pn、Tr、Gs 3个指标之间呈正相关并达到极显著水平,而Ci和Ls及WUE则表现为极显著的负相关(表1)。
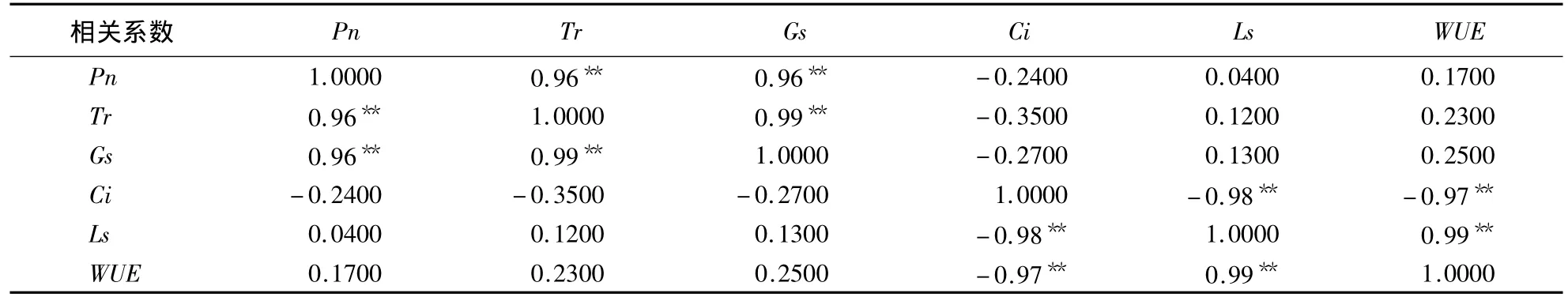
表1 台湾桤木受酸雨胁迫1个月后叶片各光合指标间的相关性分析1)Table1 Analysis of correlation among various photosynthetic indices in the leaves of A.formosana under the stress by simulated acid rain for 30 days
3 小结与讨论
光合作用是植物进行生长的基础。本研究表明,酸雨降低了台湾桤木的Pn、Tr、Gs、Ls和WUE,但一定程度上提高了Ci,并且随着时间的持续,表现出一定的适应性。酸雨中的H+浓度达到一定值时,将阻碍叶绿素的合成,从而直接影响植物的光合作用[25-26]。台湾桤木在酸雨胁迫下光合作用的抑制机理及其过程有待于进一步研究。
酸雨是一个重要的环境影响因素,前人对山核桃[1]、青冈[3]、木荷[3]、腊梅[5]、芒果[6]、马尾松[7]及杉木[7]等植物对酸雨的响应进行了研究,但鉴于研究材料年龄、叶龄、模拟酸度及测定时间等不一致,在可比性上存在一定的模糊性。但酸雨对植物存在影响是共性的问题,且酸雨酸度越高,对植物的伤害就越严重,伤害的程度与酸雨的酸性强度呈正比[29-30],本研究获得相似的结论。
酸雨对台湾桤木光合作用的影响,与酸雨酸度之间存在一定的关系,建立危害程度与酸雨胁迫强度、胁迫时间之间的关系,并优化确定台湾桤木忍耐酸雨危害的阈值对指导生产和应对酸雨危害具有重要的指导意义。因此,深入研究酸雨对台湾桤木危害的敏感器官、敏感时期并科学设计酸雨模拟模式是建立台湾桤木对酸雨影响的响应模型的基础性研究和关键技术。
植物光合特性受诸多因素的影响,如温度、湿度、CO2浓度及叶片状况、测定时间等。尽管本研究中对外界温度、湿度和CO2浓度进行设定,从而讨论不同酸雨酸度下台湾桤木叶片光合作用的差异,但季节、时点、前期天气等仍是影响植物光合特性的重要方面,其影响程度及效果还需深入研究与讨论。
本试验以模拟酸雨为胁迫方式,研究其对台湾桤木叶片光合特性的影响,但酸雨对台湾桤木的影响不仅仅停留在叶片上,必将对其植株生长、生物量等形态指标产生影响。在酸雨持续影响下其响应力与恢复能力如何,可否通过激发体内的信号传导物质或合成相关次生代谢物避免并恢复酸雨造成的伤害,有待于进一步研究和佐证。
[1]鲁美娟,江洪,余树全,等.模拟酸雨对山核桃和杨梅光合生理特征的影响[J].生态学杂志,2009,28(8):1476-1481.
[2]TATIANA S,JOHN M,ANTHONY L.Soybean electrophysiology:Effects of acid rain[J].Plant Science,2002,162:723 -731.
[3]李万超,江洪,曾波,等.模拟酸雨对青冈和木荷幼苗光合响应特性的影响[J].西南大学学报,2008,30(7):99-104.
[4]高吉喜,潘凤云,周兴宝.二氧化硫对植物新陈代谢的影响(Ⅱ)——对光合、呼吸与物质代谢的影响[J].环境科学研究,1997,10(6):5 -9.
[5]周青,黄晓华,王冬燕,等.稀土元素La对酸雨损伤腊梅的影响[J].生态学杂志,1997,16(6):59-61.
[6]陈美华,欧世金,蒋德书.模拟酸雨对芒果生长及土壤的影响[J].广西农业大学学报,1995,14(4):300-304.
[7]单运峰,冯宗炜.模拟酸雨对马尾松和杉木幼树的影响[J].环境科学学报,1988,8(3):307-315.
[8]YU J,YE S,HUANG L.Effects of simulated acid precipitation on photosynthesis,chlorophyll fluorescence,and antloxidative enzymes in Cucumis sativus L[J].Photosynthetica,2002,40(3):331 -335.
[9]单运峰.酸雨、大气污染与植物[M].北京:中国环境科学出版社,1994.
[10]SHAN Y,FENG Z,IZUTA T,et al.The individual and combined effects of ozone and simulated acid rain on chlorophyll contents,carbon allocation and biomass accumulation of armand pine seedlings[J].Water,Air & Soil Pollution,1995,85(3):1399-1404.
[11]王军辉,顾万春,李斌,等.桤木优良种源/家系的选择研究[J].林业科学,2000,36(3):58-60.
[12]王军辉,顾万春,夏良放,等.桤木种源的地理变异和种源区划[J].浙江林学院学报,2005,22(5):502-506.
[13]郑兆飞.桤木杉木混交林水源涵养能力研究[J].西南林学院学报,2008,28(3):8-10.
[14]麻文礼.桤木在武夷山引种造林对比试验初报[J].林业科技开发,2003(17):27-30.
[15]陈益泰,卓仁英,吴天林.桤木属植物的引种和早期适应性[J].林业科学研究,2004,17(2):139-146.
[16]刘贤词,文仕知,冯汉华,等.四川桤木人工林不同年龄段生物量的研究[J].中南林业科技大学学报,2007,27(2):83-86.
[17]林家财.四川桤木人工林林分结构规律研究[J].武夷科学,2007,23(2):115-119.
[18]朱万泽,王金锡,薛建辉.台湾桤木和四川桤木种源苗木对水分胁迫的生理响应[J].西北植物学报,2005,25(10):1969-1975.
[19]朱万泽,薛建辉,王金锡.台湾桤木种源对水份胁迫的光合响应及其抗旱性[J].水土保持学报,2004,18(4):171-181.
[20]罗正华,李贤伟,袁渭阳,等.台湾桤木幼林地细根生长及林地土壤抗蚀性分析[J].四川农业大学学报,2008,26(3):245-250.
[21]朱万泽,王金锡.引种台湾桤木的蒸腾特性及其影响因子[J].四川林业科技,2005,26(1):10-14.
[22]BERRY J,DOWNTON W.Environmental regulation of photosynthesis[M].New York:Academic Press,1982.
[23]吕均良,李三玉,黄寿波.模拟酸雨对桃梨叶片和果实的影响[J].浙江农业大学学报,1998,24(6):603-607.
[24]李志国,姜卫兵,翁忙玲,等.常绿阔叶园林6树种(品种)对模拟酸雨的生理响应及敏感性[J].园艺学报,2011,38(3):512-518.
[25]周海燕,黄子琛.不同时期毛乌素沙区主要植物种光合作用和蒸腾作用的变化[J].植物生态学报,1996,20(2):120-131.
[26]杜占池,杨宗贵.不同土壤型羊草光合和蒸腾作用特性的比较研究[J].植物学报,1995,37(1):66-73.
[27]朱万泽,王金锡.引种台湾桤木的蒸腾特性及其影响因子[J].四川林业科技,2005,26(1):10-15.
[28]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annu Rev Plant Physiol,1982,33:317 -345.
[29]杨振德,陈锐亮,程玉兰.几种园林树种对酸雨抗性的研究[J].广西大学学报,1999,3(2):29.
[30]邱栋梁,刘星辉.酸雨对园艺植物危害的研究进展[J].福建农业大学学报,1999,28(1):28-32.
猜你喜欢
中国书法(2023年9期)2023-12-26 18:03:59
云南化工(2020年11期)2021-01-14 00:50:56
河北地质(2017年2期)2017-08-16 03:17:15
陕西气象(2017年4期)2017-08-02 01:41:14
环境保护与循环经济(2017年8期)2017-03-22 01:29:06
中南林业科技大学学报(2016年1期)2016-12-20 08:09:31
广西林业科学(2016年3期)2016-03-16 05:43:36
福建质量管理(2015年9期)2015-04-17 18:28:24
装备环境工程(2015年4期)2015-02-28 01:20:05
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:56
