
特种野猪ESR基因、FSHβ基因的多态性分析
2012-09-22 07:35:06何若钢刘丁健刘桂武张家富李斯丽涂兴强
饲料工业 2012年9期
何若钢 刘 谨 刘丁健 刘桂武 张家富 李斯丽 涂兴强 言 稳
(广西大学动物科技学院,广西南宁 530005)
特种野猪也称为杂优野猪,它是以纯种野公猪作为父本、优良瘦肉型猪或者本地猪为母本进行杂交,杂交后代再与纯种野公猪进行回交,以提高特种野猪的含野血缘,这样形成的一种野猪称为特种野猪。特种野猪具有浓郁的野味以及较高的瘦肉率,其肉质鲜美柔嫩香醇,脂肪含量低,营养丰富全面,含有很多种氨基酸以及很多种微量矿物质元素,渐渐成为当代新型的绿色食品。目前在市场上十分畅销,尤其是香港、广州、北京、上海等发达都市需求量相当大,特种野猪肉50元/kg仍然供不应求。
特种野猪是优良山林野公猪与优良瘦肉型母猪进行多次杂交后,经人工多次驯化的杂优野猪,其基因稳定,不易产生变异,后代可以长期做种繁殖而基因保持稳定。由于特种野猪是野猪与家猪的杂交产物,表现出很好的杂交优势,家猪、野猪的优点于一身,它既保持了纯种野猪肉质鲜嫩、风味鲜美、瘦肉率高、抗病力强、耐粗饲能力强、对环境的适应性好的优势,又保持了家猪生长速度快、饲料转化率高、繁殖力强的特点,并且克服了纯种野猪生长慢、体形偏小、季节性繁殖的缺点。所以养殖特种野猪已变成养殖业中的热门话题,属于国家科技部星火计划推广项目之一。从分子遗传学角度对特种野猪的优异特性进行全面研究非常必要。本研究收集了72头含50%野猪血缘的特种野母猪耳组织样品,对影响繁殖性状的主要功能基因(ESR、FSHβ)的多态性进行检测和分析,通过分子育种的手段提高特种野猪的产仔数,提高特种野猪的繁殖性能,为特种野猪遗传资源的保护和开发利用提供基础资料及科学依据。
1 材料和方法
1.1 样本采集和基因组提取
本研究共采集了72头含50%野猪血缘的特种野母猪耳组织样品。样品来自上思县殴达畜牧业有限责任公司旗下的特种野猪驯养场。耳组织样用装有70%乙醇的试管带回实验室,置于-20℃的冰箱保存备用。盐析法提取基因组DNA。
1.2 仪器和材料
蛋白酶 K,Sigma 公司;MarkerDL2000:takara 公司;2Xtaq Master Mix:北京天根生物有限公司;Taq酶、dNTP、MgCl2,MgSO4、PCRbuffer,Promega 公司;GelRed核酸凝胶染色剂:美国Biotium公司;无水乙醇、异丙醇等常规国产试剂:成都市成龙化工厂;琼脂糖:西班牙Biowest原装;琼脂糖胶回收试剂盒:北京天根生物有限公司。
1.3 引物设计
ESR和FSHβ基因的引物如表1所示。
1.4 PCR扩增
1.4.1 反应体系
反应总体积分别为 50 μl和 25 μl,包括:①ESR PCR 反应体系:2Xtaq Mastermix,25 μl;ESR 上游引物(20 μmol/l)1 μl;ESR 下游引物 (20 μmol/l)1 μl;DNA模板(20 ng/μl)1 μl;灭菌 ddH2O 22 μl。②FSHβPCR反应体系:2Xtaq Mastermix,12.5 μl;FSHβ 上游引物(20 μmol/l)0.5 μl;FSHβ 下游引物(20 μmol/l)0.5 μl;DNA 模板(20 ng/μl)0.5 μl;灭菌 ddH2O 11 μl。

表1 ESR和FSHβ基因的引物序列
1.4.2 反应程序
ESR PCR反应程序为:95℃预变性3 min;94℃变性 1 min;56℃退火 30 s,72℃延伸 30 s;返回第2步,进行35个循环;72℃再延伸5 min;4℃保温。
FSHβ PCR反应程序为:95℃预变性3 min;94℃变性1 min;55℃退火45 s,72℃延伸45 s;返回第2步,进行35个循环;72℃再延伸5 min;4℃保温。
1.5 琼脂糖凝胶电泳及ESR片段的胶回收
扩增产物用1.5%琼脂糖凝胶电泳(5V/CM),在微波炉加热后加入适量的核酸染料GelRed,电泳缓冲液为1×TAE,在伯乐凝胶成像系统中成像观察并切胶、回收特异性目的片段(ESR),回收方法按照天根生化公司的胶回收试剂盒进行。将回收片段样品进行酶切分析。
1.6 PCR产物酶切
PCR酶切方法是首先取5 μl PCR产物到PCR管中,然后加入1.2 μl内切酶PvuⅡ,同时加入1 μl 10×buffer的内切酶PvuⅡ稀释液,加入12.8 μl灭菌超纯水,总共是20 μl体积,放在37℃水浴锅中水浴8 h,最后于4%琼脂糖凝胶电泳检测。
1.7 ESR、FSHβ基因型分析
ESR基因型分析:ESR基因中B等位基因一个碱基对发生点突变,导致ESR基因座位等位基因表现出差异。这种差异产生了一个限制性内切酶PvuⅡ的酶切位点,ESR基因的PCR产物经限制性内切酶PvuⅡ酶切后呈现出多态性,根据其条带的数目和位置,确定有AA、AB、BB 3种基因型,其等位基因A的片段为121 bp,等位基因B的片段为(56+65)bp。据此,AA基因型为一条带,为121 bp,AB基因型为三条带,为 121、65、56 bp,BB 基因型为两条带,为 65 bp和56 bp。
FSHβ基因型分析:FSHβ基因中A等位基因在其第1内含子中存在1个292 bp的逆转座子插入突变,导致A、B等位基因的长度出现差异,这造成FSHβ基因座位的等位基因差异,这种差异在电泳中产生不同图谱,其等位基因A的扩增片段为500 bp,等位基因B的扩增片段为220 bp。据此也可将FSHβ基因座位分为AA,AB和BB 3种基因型,若扩增结果出现500 bp的单条带则为AA型,若出现500 bp和220 bp两条带则为AB型,若出现220 bp一条带则为BB型。
1.8 数据处理
分析所有基因图谱,整理基因型数据,计算所测特种野猪群ESR基因以及FSHβ基因中不同基因型的基因型频率、各等位基因在整个群体中出现的基因频率,用SPSS分析各基因型产仔数的差异显著性,数据用平均数±标准差表示。
2 结果与分析
2.1 ESR-PCR扩增以及酶切结果
2.1.1 ESR-PCR扩增结果
PCR扩增产物电泳图谱见图1。
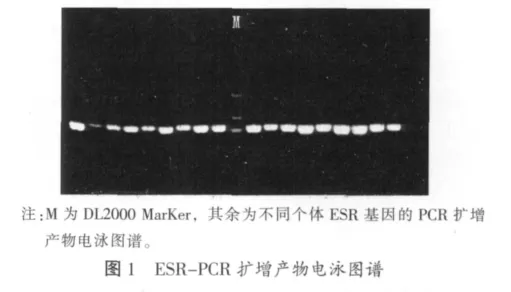
2.1.2 ESR酶切结果
扩增产物经限制性内切酶Pvu Il酶切后出现多态,在图中根据条带的数目和位置,我们可以确定有AA、AB 两种基因型,分别为(AA 121 bp);AB(3条带,65+56+121 bp)
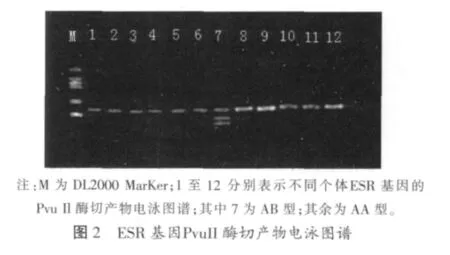
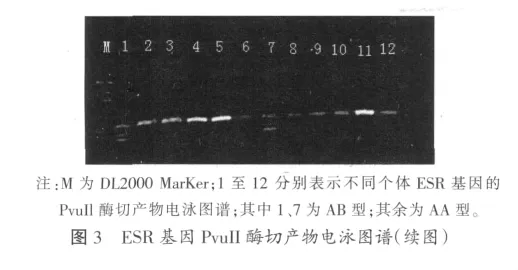
2.2 FSHβ-PCR扩增结果
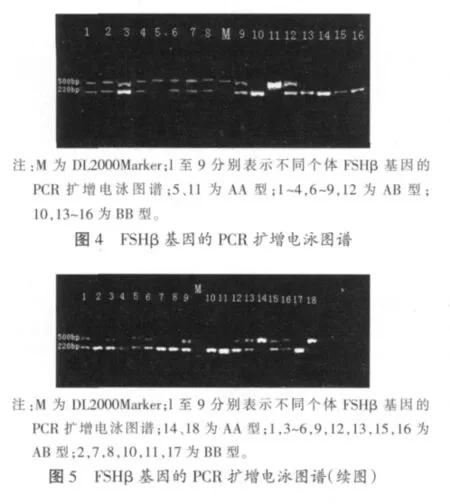
FSHβ经PCR扩增后其扩增产物于1%琼脂糖凝胶检测,电泳图谱如图4和图5。基因只表现出AA和AB两种基因型。其频率分别为91.67%和8.33%,等位基因A和B的频率分别为95.83%和4.17%,AA型以及等位基因A的频率均超过90%,在群体中占据明显的优势地位。FSHβ亚基基因表现出三种基因型AA、AB和BB,其频率分别为11.11%、55.56%以及33.33%,AB型为优势基因型。等位基因A和B的频率分别为38.89%和61.11%,等位基因B在群体中占据优势。
2.3.2 ESR、FSHβ不同基因型与产仔数关系
在被检特种野猪群体中ESR基因缺少BB型,ESR基因、FSHβ亚基基因不同基因型之间与总产仔数、产活仔数以及初生重的关系见表3。
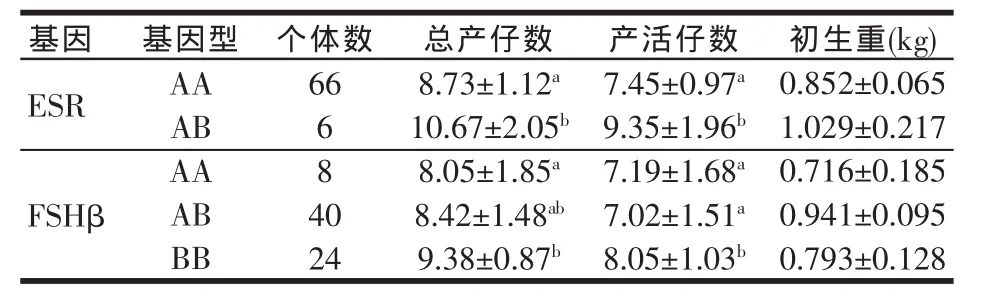
表3 特种野猪ESR、FSHβ不同基因型与产仔数关系
2.3 统计分析结果
2.3.1 ESR、FSHβ亚基因多态在特种野猪中的分布及作用
在所检测的72头特种野猪中ESR和FSHβ亚基基因不同,基因型分布(头数)、基因型频率和基因频率见表2。
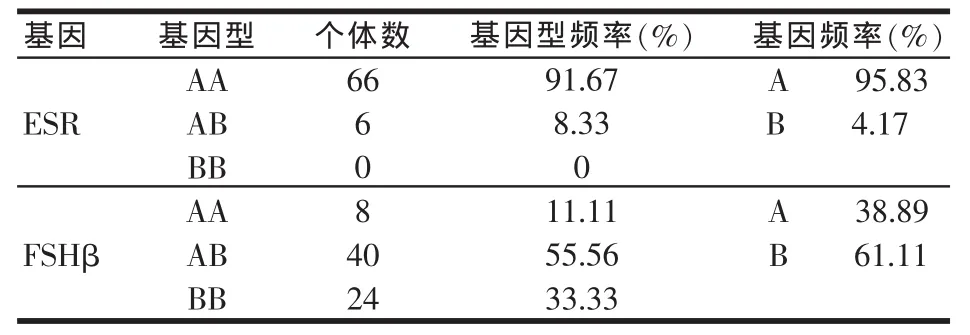
表2 特种野猪ESR和FSHβ基因的基因型频率及基因频率
从表2可以看出,在所检测的特种野猪中,ESR
由表3可知,在所检测的特种野猪群中,ESR基因中AA和AB两种基因型对应的猪总产仔数、产活仔数存在显著差异(P<0.05),且AB>AA基因型,AB型比AA型的猪总产仔数、产活仔数分别高1.94头以及1.9头。不同基因型对仔猪出生重的影响差异不显著(P>0.05)。FSHβ基因AA、AB和BB三种基因型对应的猪总产仔数和产活仔数均以BB型最高,显示出BB>AB>AA基因型的趋势。其中BB型对应的猪总产仔数显著高于AA型(P<0.05),比AA型高1.33头,BB型产活仔数显著高于AA型以及AB型(P<0.05),分别高0.85头和1.03头。
3 讨论
3.1 ESR基因多态性及其与产仔数的关联分析
在所检测的72头特种野猪中,ESR基因不同基因型猪总产仔数、产活仔数均存在AB基因型>AA基因型,且AB型猪总产仔数、产活仔数均显著高于AA型,这表明ESR基因是控制产仔数的主基因或与产仔数主基因存在紧密连锁,而B等位基因是增加产仔数的优势基因。这与赵西彪等、丁家桐等研究结果一致。
李凤娥等、Natoloczna-Kotara对不同猪品种ESR基因进行了多态性分析,统计了ESR基因B等位基因在不同品种中的频率,结果表明:大白×梅山(0.59)、梅山猪(0.50)、中国瘦肉猪新品系 DIV2(0.68)、清平猪(0.90)、通城猪(0.67),这表明繁殖性能高的中国本地猪种含有较高优势基因B。而在本试验中特种野猪的ESR等位基因A的频率为0.958 3,在群体中占绝对优势地位,这可能是导致特种野猪产仔数偏低的主要原因。
根据试验结论推断,ESR为BB基因型的母猪产仔数应最多,但在本试验所检测的特种野猪群中ESR基因缺少BB基因型,这可能是因为等位基因B在群体中所占的基因频率太低,不容易被检测到所致。另外,样本量偏小可能也是导致没有检测到ESR基因BB基因型的一个因素。统计结果发现仔猪初生重在不同基因型之间没有显著影响,这和国内外的多数报道一致,这说明ESR对仔猪初生重不产生显著影响,只能提高母猪的产仔数。
3.2 FSHβ基因多态性及其与产仔数的关联分析
本次在特种野猪群体中对FSHβ基因多态性的检测共检测到AA、AB、BB三种基因型,虽然AB基因型总产仔数与AA、BB基因型差异未达到显著水平,AB基因型产活仔数与AA基因型差异也未达显著水平,但三种基因型总产仔数和产活仔数呈现BB>AB>AA的趋势,这表明对于FSHβ基因而言,等位基因B是影响产仔数的优势基因,鲁绍雄等在撤坝猪ESR和FSHβ基因多态性及其与产仔数的关联分析中得到FSHβ基因对撒坝猪产仔数的影响是FSHβ亚基基因BB型为猪产仔数增效基因型,等位基因B为产仔数增效基因;周胜花等在ESR和FSHβ基因与晋阳白猪繁殖性状关系研究中得到FSHβ基因为BB基因型的母猪产仔数最多,FSHβ基因的有利基因B频率较高,晋阳白猪高繁殖性能可能与有利基因B的频率较高有关。本研究与以上研究结果一致。
FSHβ亚基基因的多态性分析中发现国内地方猪种如莱芜黑猪、香猪、二花脸猪、民猪、撒坝猪等群体中A等位基因占优势,而在国外著名猪种中如约克夏、长白等群体中等位基因B占优势。本研究中特种野猪FSHβ亚基B基因频率占优势,为0.611 1,这与报道过的其它中国地方猪种的分布频率有很大的偏差,这可能与特种野猪在培育的过程中引入外血有关。
4 结论
在所检测的特种野猪群中,ESR基因,AB型比AA型的猪总产仔数、产活仔数分别高1.94头以及1.9头。FSHβ基因BB基因型的猪总产仔数和产活仔数最高,显示出BB>AB>AA基因型的趋势,其中BB型比AA型高1.33头,BB型产活仔数分别比AA、AB高0.85头和1.03头。ESR、FSHβ不同基因型对仔猪初生重的影响差异不显著。
从本次检测结果来看,被检测的特种野猪ESR基因和FSHβ基因不同基因型能对产仔数产生显著影响,因此FSHβ基因和ESR基因可以作为控制猪高繁殖率的候选基因。ESR基因能增加产仔数的优势基因B频率相当低,为0.041 7;而FSHβ基因中能增加产仔数的优势基因B的频率较高,为0.611 1。因此,对该群体来说,注意提高ESR基因中等位基因B的频率,将能显著提高猪群的繁殖性能。在以后的育种工作中,可以把具有ESR基因的B等位基因的个体予以保留,通过标记辅助选择(MAS)来改良猪产仔数性状,从而带来巨大的潜在经济效益。
[1]熊沈学,沈丽玲.浅议特种野猪饲养管理[J].经济动物,2009(3):105-106.
[2]马锦屏.特种野猪的饲养管理[J].畜牧兽医科技信息,2008(1):86.
[3]洪学.特种野猪养殖的优势与效益分析[J].中国养猪,2009(6):35.
[4]SHORT T H,ROTHCHILD M F,SOUTHWOOD O I,et al.Effect of the estrogen receptor locus on reproduction traits in four commercial pig lines[J].jounal of animal science,1997,75:3138-3142.
[5]赵要风,李宁,肖潞,等.猪FSHβ亚基基因结构区逆转座子插入突变及其与猪产仔数关系的研究[J].中国科学(C辑),1999,29(1):81-86.
[6]赵西彪,于钦秀,等.ESR和FSHβ基因与梅山猪产仔数关系研究[J].猪业科学,2008(2):28-29.
[7]丁家桐,葛红山,等.猪雌激素受体基因与产仔数和乳头数的关系研究[J].南京农业大学学报,2002,25(2):79-81.
[8]李凤娥,熊远著,等.猪品种间ESR基因PCR RFLP的初步研究[J].华中农业大学学报,2000,19(1):37-39.
[9]Natoloczna-Kotara A,Jasek S1 Association between polymorphism in ESR gene and litter size of Polish Landrace and Polish Large White sows-preliminary results[J].Annals of Animal Science,2001,21(1):153-157.
[10]王继英,郭建凤,郝小静,等.山东省八个猪种繁殖、肉质、抗病基因多态性分析[J].华北农业学报,2009,24(6)227-234.
[11]鲁绍雄,胡晓湘,等.撤坝猪ESR和FSHβ基因多态性及其与产仔数的关联分析[J].云南农业大学学报,2009,24(3):389-393.
[12]周胜花,贺东昌,等.ESR和FSHβ基因与晋阳白猪繁殖性状关系研究[J].中国畜牧兽医,2009,36(12):97-99.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23 06:21:42
小天使·一年级语数英综合(2022年3期)2022-04-16 05:07:32
现代畜牧科技(2021年6期)2021-07-16 05:50:08
中国畜牧杂志(2020年8期)2020-08-16 23:52:44
国外畜牧学·猪与禽(2018年4期)2018-05-14 11:14:19
小学阅读指南·低年级版(2017年6期)2017-06-12 07:38:57
猪业科学(2016年2期)2016-04-25 07:02:13
读写算(下)(2015年6期)2015-08-22 05:57:56
中国动物保健(2015年7期)2015-02-05 10:59:38
中国饲料(2015年18期)2015-01-25 12:09:03
