
帽儿山地区杂木林更新演替趋势分析1)
2012-09-18 01:27张命军王庆成薛思雷刘忠玲吴文娟
东北林业大学学报 2012年3期
张命军 王庆成 薛思雷 刘忠玲 吴文娟
(东北林业大学,哈尔滨,150040)
杂木林中树种组成复杂,各树种种子库储量是否充足,种子能否形成幼苗,幼苗能否形成幼树进入演替层,决定着森林更新动态。森林土壤种子库是植被天然更新的物质基础[1],与植被演替动态和天然更新有密切关系[2-3],很大程度上决定了植被发展的进程和方向[4-5]。地表的枯枝落叶、地被物等阻碍种子与土壤接触,影响土壤种子库的建成和结构[6-7],导致不同层的种子储量和萌发情况不同[8-9],最终将影响森林天然更新的效果[10-11]。天然更新除需有充足的、有活力的种子作为基础外,还必须有促进幼苗生长存活、幼树生长的种群结构[12-15]。种群结构对分析和预测种群动态具有重要意义。进展型种群标志着森林演替的趋势,稳定型种群将是重要的伴生树种,衰退型种群将在未来的群落中被淘汰[16]。以往主要通过种群结构研究群落更新与演替[17-20],而通过土壤种子库与种群结构相结合研究更新与演替的报道较少,本文通过对东北林业大学帽儿山实验林场主要森林类型——杂木林林地土壤种子库及分层株数调查,分析林分天然更新状况和演替趋势,为东北东部地区杂木林的经营管理提供理论依据。
1 研究地概况
试验地设立在东北林业大学帽儿山实验林场尖砬沟森林培育实验站(E127°30'~127°34',N45°21'~45°25')的杂木林中。该区为低山丘陵地带,属大陆性季风气候,四季分明。降水量600~800 mm,集中于7、8月份,年蒸发量1093 mm。年平均温度2.7℃,1月平均温度-19.6℃,7月平均温度20.9℃,全年大于或等于10℃的积温为2638℃。早霜一般出现在8月末,晚霜一般在5月末止,无霜期120~140 d。平均海拔300 m,地带性土壤为暗棕色森林土。该区植被属于长白植被区系,原地带性植被为阔叶红松林,历经大规模破坏(采伐、火烧、开垦等),现为次生演替成的天然次生林相,是东北东部山区天然次生林的典型林型。主要乔木有水曲柳(Fraxinus mandshurica)、蒙古栎(Quercus mongolica)、白桦 (Betula platyphylla)、胡桃楸 (Juglans mandshurica)、黄波椤(Phellodendron amurense)、山杨(Populus davidiana)、紫椴(Tilia amurensis)、枫桦(Betula costata)和春榆(Ulmus propinqua)等。灌木树种主要有暴马丁香(Syringa amurensis)、毛榛(Corylus mandshurica)等。草本植物主要有木贼(Equisetum ramosissimum)、细叶苔草(Carex rigescens)、蚊子草(Filipendula palmata)和毛缘苔草(Carex campylorhina)等。
研究的杂木林位于中坡位,坡度12°,坡向为西坡。主林层树种组成为:2山+1蒙+1胡+1五+1水+1椴+1黄+1榆+1白,主林层林分密度为763株·hm-2,郁闭度为 0.8。
2 研究方法
2.1 土壤种子库
2010年9月,在杂木林内随机设置12块6.0 m×6.0 m的大样方,将其进一步划分为36个网格状的1.0 m×1.0 m的小样方。在36个小样方中随机选取9个进行土壤种子库的取样。用直径为10 cm的圆筒形取样器,分别取未分解有机层(Oi)、分解有机层(Oe)、腐殖质层(A,0~5 cm)样品,每层取样108个。样品分别装入自封袋中,标号,带回实验室。手工挑拣把种子从各样品中分离出来,记录形态完整、形态不完整(破损、瘪粒、虫害和动物取食)的种子数量。计算各层种子不同形态的百分含量,公式为:
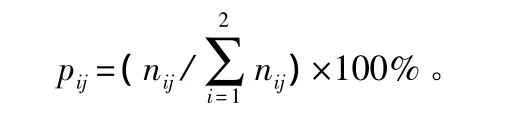
式中:i为种子形态(完整、不完整);j为土壤层次(Oi层,Oe层和A层);p为各层不同形态种子比例;n为种子储量。
2.2 种群结构
在林内设置3块30.0 m×30.0 m的样地,将每块样地划分为9个10.0 m×10.0 m的样方,总计27个。在每个样方中对主林层(林冠层)、演替层(2.0 m<树高<林冠层高度)[19-20]的乔木进行每木检尺,记录树种、树高、胸径。在每个10.0 m×10.0 m 的样方内沿对角线设置3个1.0 m×1.0 m的小样方,总计81个。在每个小样方中对更新层(树高<2.0 m)的幼苗、幼树进行调查,记录树种、苗高、地径。根据调查结果,对各层不同树种进行分层株数分析。
分层株数计算公式:
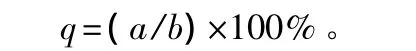
式中:q为分层数量;a为某树种在某层次中的数量;b为该树种在所有层次中的总数。
分层频度计算公式:
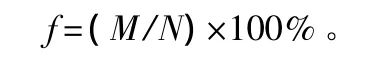
式中:f为分层频度;M为某种出现在某层样方中的数量;N为样方总数。
2.3 更新幼苗树龄分布
调查结束后,在林分内依不同郁闭度选取不同高度的水曲柳幼苗58株,蒙古栎幼苗36株,胡桃楸幼苗14株(更新苗较少)。测定幼苗高和地径,然后挖取幼苗,整株带回实验室,通过幼苗根茎处年轮确定幼苗树龄。
2.4 数据分析
应用SPASS软件(SPSS公司,13.0)对种子储量进行单因子方差分析和最小显著差数法(LSD)检验。
3 结果与分析
3.1 杂木林土壤种子库主要树种种子储量
在调查的林地土壤种子库中,水曲柳种子的总储量平均为600.4粒·m-2(表1),占4种种子总储量的86.3%;五角槭、蒙古栎总储量较少,分别占7.3%、4.1%;胡桃楸种子最少,为 2.4%。
土壤种子库中除胡桃楸外,其它3个树种的种子储量在Oe层储量最高,在A层储量最低(表1)。水曲柳、五角槭种子在Oe层的储量显著高于Oi层和A层(P<0.05),Oi和A层之间无显著差异。蒙古栎种子在Oe层的储量显著高于A层(P<0.05),Oi层和A层之间无显著差异。胡桃楸种子在各层的储量无显著差异(表1)。
形态完整的水曲柳、五角槭种子随着土层深度的增加而减少。形态完整的水曲柳种子在Oi层、Oe层、A 层的比例分别为 94.3%、79.2%和 63.9%,形态完整的五角槭种子在Oi层、Oe层、A层的比例分别为75%、23.5%和0(表1);留存的蒙古栎种子,有75.0%被动物取食过,20.8%为瘪粒;胡桃楸种子中有85.7%存在动物取食痕迹。蒙古栎和胡桃楸种子在样地中分布较少,其中蒙古栎种子只在12个样方(有2个样方两层有种子)出现过,且在Oi层和Oe层未发现完整种子,只在A层发现1粒发芽的完整种子。胡桃楸也只在12个样方(有2个样方两层有种子)中出现过,A层未发现完整种子,只在Oi层和Oe层分别发现1粒完整种子(表1)。
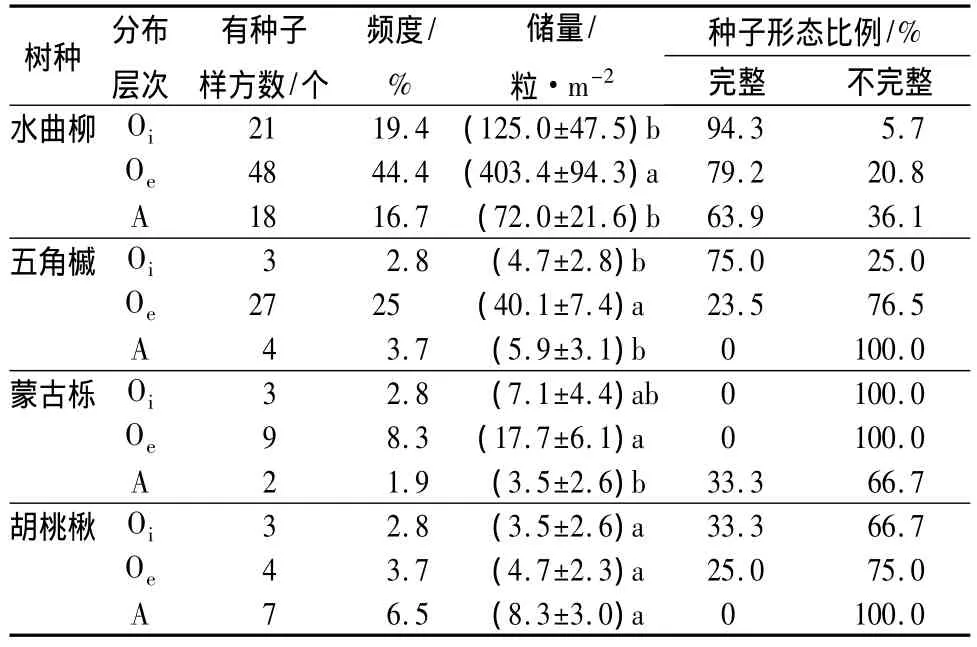
表1 帽儿山杂木林土壤种子库中不同树种各形态种子分层储量
3.2 杂木林种群结构分析
3.2.1 杂木林树种组成
杂木林主林层、演替层、更新层平均树高分别为16.2、6.1、0.34 m,平均胸径(或地径)分别为 18.3、4.6、0.35 cm。主林层树种组成为:2山1蒙1胡1五1水1椴1黄1榆1白,以山杨为主,同时有蒙古栎、五角槭、水曲柳、胡桃楸、紫椴等树种。演替层树种组成为:6五1蒙1榆1水+紫+山-胡-白,以五角槭为主,同时有蒙古栎、春榆、水曲柳等树种。更新层树种组成为:4五4山2水+紫-蒙,以五角槭、山杨、水曲柳为主(表2)。
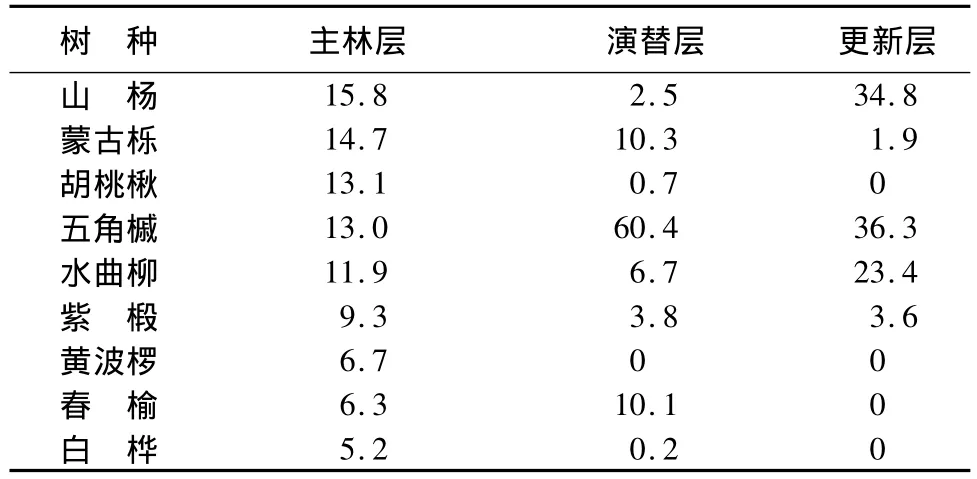
表2 帽儿山杂木林各林层树种组成 %
3.2.2 杂木林不同光照条件下更新层幼苗的生长
平均树龄相同的水曲柳幼苗,其平均高和地径均随郁闭度降低而增加。水曲柳幼苗平均树龄4 a,最大树龄可达9 a(表3)。蒙古栎幼苗在郁闭度较小时,幼苗平均高、平均地径和平均树龄较高,随着郁闭度增加,幼苗平均高、地径和树龄有减小的趋势,郁闭度增加时幼苗最大树龄减小(表3)。随着郁闭度的增加,胡桃楸幼苗的平均树龄减少(表3)。
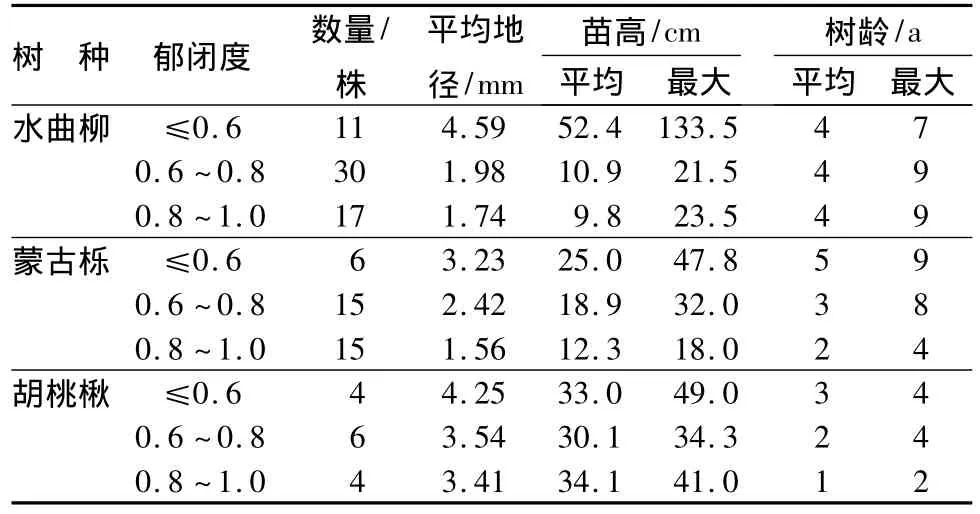
表3 不同光照条件下更新层的幼苗生长量
水曲柳、蒙古栎和胡桃楸幼苗高生长主要发生在第1年,2年生苗木的苗高与1年生相比差异不明显(表4)。
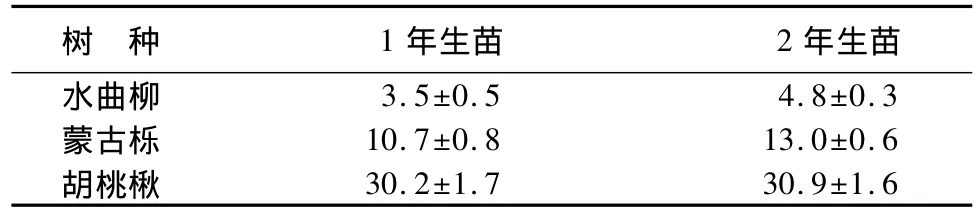
表4 不同树种的1年生、2年生更新苗的平均高 cm
3.2.3 杂木林种群动态分析
杂木林中,水曲柳、蒙古栎、紫椴、山杨分层株数比为:更新层>主林层>演替层(表5)。水曲柳主林层与演替层分层株数比相近,更新层相对较大,密度达3704株·hm-2,明显高于其它树种。五角槭分层株数比为:更新层>演替层>主林层(表5)。五角槭在更新层中的密度仅次于水曲柳,但五角槭在主林层和演替层的密度均最高,分别为119和852株·hm-2。五角槭在演替层的分层频度达到100%,在主林层和更新层也有55.6%和16.0%的分布。
春榆分层株数比和分布频度均为:演替层>主林层>更新层(表5),在演替层的密度和分布频度仅低于五角枫。
黄波椤、胡桃楸和白桦主林层分层株树比和分布频度远高于演替层,呈倒塔形分布。稠李(Padus racemosa)、白牛槭(Acer mandshuricum)、枫桦、红松(pinus koraiensis)在林分中密度较小,仅在主林层和演替层有零星分布(表5)。
4 结论与讨论
杂木林下种子库中水曲柳种子储量最大(表1),虽然水曲柳种子主要集中在Oe层,但各层均有较大储量,水曲柳种子能较顺利向幼苗转化,更新层幼苗密度达3704株·hm-2(表5)。调查发现,演替层中没有2~5 m高的幼树。刘足根等[21]的研究也发现,次生林中树高小于2 m的水曲柳幼苗较多,没有树高超过2 m、胸径小于7.5 cm的幼树。更新层水曲柳幼苗平均树龄为4 a,最大树龄9 a(表3),说明水曲柳幼苗可以在林下存活并能适应较长时间的遮光环境,只是其生长受到限制,不能顺利向演替层发展。霍常富等[22]的研究认为,水曲柳对弱光环境具有一定的适应能力,但遮光会使苗木叶片的光合速率显著下降而抑制植株的生长。所以水曲柳天然更新受限的主要原因是幼苗难以转化为幼树。
蒙古栎各层储量均较小(表1),原因是啮齿类动物取食量较大[23-24],此外,大部分成熟脱落的种子有昆虫幼虫寄居。程积民等[25]认为,缺少种子是影响蒙古栎天然更新的主要原因之一。蒙古栎幼苗第1年高生长较大,第2年高生长降低(表4),其主要原因是蒙古栎是大粒种子,种子中储存的干物质较多,可以满足第1年高生长的需要,而后其高生长逐年下降[26]。随着杂木林郁闭度的增加,其林下蒙古栎幼苗的平均树龄降低(表3),说明蒙古栎在庇荫环境中幼苗存活时间缩短。许中旗等[26]的研究认为,林冠下蒙古栎幼苗可通过增加比叶面积在一定程度上缓解由于光照不足所导致的碳收入不足,但只能在一定程度上延长其在林下的存活时间。如果林分的光照条件不能得到改善,蒙古栎幼苗将趋于死亡。所以蒙古栎幼苗天然更新的另一个限制是幼苗向幼树转化困难。
胡桃楸土壤种子库中各层储量均很少(表1),原因是人为采摘以及种子凋落后啮齿类动物取食所致。种子的极度缺乏是限制胡桃楸天然更新的原因之一。随着杂木林郁闭度的增加,林下胡桃楸幼苗存活率降低,当杂木林郁闭度较大时,胡桃楸平均树龄只有1 a,最大树龄为2 a(表3)。第1年幼苗有较大的高生长,第2年高生长降低(表4),可见胡桃楸幼苗在遮光的条件下,生长受到极大的限制。王凯等[27]的研究表明,随着生长环境光强的降低,胡桃楸幼苗的相对生长率显著下降。马万里等[28]的研究发现,胡桃楸幼苗和幼树个体比例较小,种群在幼年期死亡率较高。因而,种子极度缺乏以及幼苗在林冠下存活率低,是限制胡桃楸天然更新的主要原因,在未来的演替中可能衰退。
五角槭土壤种子库中Oe层储量丰富,更新层幼苗较多,可以顺利地向演替层生长(演替层幼苗密度852株·hm-2,表5)。大量种子以及幼苗、幼树的存在为五角槭的更新演替提供了良好的基础,而且五角槭在演替层和主林层的分布密度也较大(表5),将成为该林分的优势种。
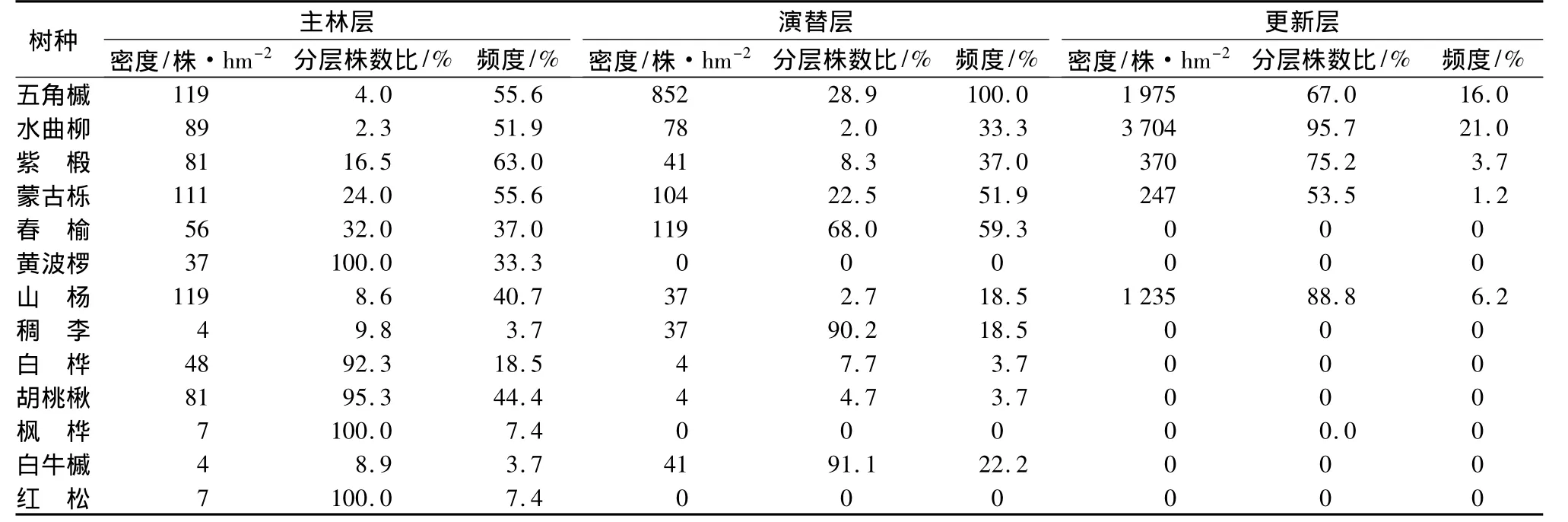
表5 帽儿山杂木林各树种在不同林层内的密度、分层株数比及频度
根据林地种子库的调查以及各林层天然更新演替的分析可以认为,如果不加人工干预,该地区杂木林珍贵树种水曲柳、胡桃楸和蒙古栎更新困难,将演替为五角槭占优势的林分。本文只是对杂木林更新演替进行了初步的研究与探讨,还有待进一步的研究,尤其是更新层幼苗的树龄结构还需进一步深入的探讨。
[1]Moles A T,Drake D R.Potential contribution of the seed rain and seed bank regeneration of native forest under plantation pine in New Zealand[J].New Zealand Journal of Botany,1999,37(1):83-93.
[2]李根柱,王贺新,朱书全,等.东北次生林区枯落物对天然更新的障碍作用[J].辽宁工程技术大学学报:自然科学版,2008,27(2):295-298.
[3]唐勇,曹敏,张建侯,等.西双版纳热带森林土壤种子库与地上植被的关系[J].应用生态学报,1999,10(3):279-282.
[4]Adel J,Behnam H,Younes A,et al.Soil seed banks in the Arasbaran Protected Area of Iran and their significance for conservation management[J].Biological Conservation,2003,109(3):425-431.
[5]Falińska K.Seed bank dynamics in abandoned meadows during a 20-year period in the Bialowieza National Park[J].Journal of E-cology,1999,87(3):461-475.
[6]刘旭,程瑞梅,肖文发.土壤种子库研究进展[J].世界林业研究,2008,21(1):27-33.
[7]Facelli J M,Pickett S T A.Plant litter:Its dynamics and effects on plant community structure[J].The Botanical Review,1991,57(1):1-32.
[8]羊留冬,杨燕,王根绪.森林凋落物对种子萌发与幼苗生长的影响[J].生态学杂志,2010,29(9):1820-1826.
[9]Pierson E A,Mack R N.The population biology of Bromus tectorum in forests:Effect of disturbance,grazing,and litter on seedling establishment and reproduction[J].Oecologia,1990,84(4):526-533.
[10]Gillman L N,Ogden J,Wright S D,et al.The influence of macro-litterfall and forest structure on litterfall damage to seedlings[J].Austral Ecology,2004,29(3):305-312.
[11]徐振邦,代力民,陈吉泉,等.长白山红松阔叶混交林森林天然更新条件的研究[J].生态学报,2001,21(9):1413-1420.
[12]刘庆,尹华军,程新颖,等.中国人工林生态系统的可持续更新问题与对策[J].世界林业研究,2010,23(1):71-75.
[13]朱教君,刘足根,王贺新.辽东山区长白落叶松人工林天然更新障碍分析[J].应用生态学报,2008,19(4):695-703.
[14]Zhu Jiaojun,Matsuzaki T,Lee Fengqin,et al.Effects of gap size created by thinning on seedling emergency,survival and establishment in a coastal pine forest[J].Forest Ecology and Management,2003,182(1/3):339-354.
[15]Gong Y L,Swaine M D,Miller H G.Effects of fencing and ground preparation on natural regeneration of native pinewood over 12 years in Glen Tanar,Aberdeenshire[J].Forestry,1991,64(2):157-168.
[16]李博.生态学[M].北京:高等教育出版社,2000.
[17]达光文.旱泉沟流域次生林演替过程中种群格局动态[J].草业科学,2009,26(6):41-46.
[18]钟晓青,张宏达,方炜.广东封开黑石顶一种亚热带常绿阔叶林群落演替动态研究[J].林业科学,1996,32(4):305-310.
[19]柴一新,祝宁.哈尔滨市城市森林的天然更新[J].东北林业大学学报,2010,38(3):8-10.
[20]佟静秋,牟长城,赖富丽.哈尔滨城市人工林自然演替趋势[J].东北林业大学学报,2009,37(3):24-25.
[21]刘足根,朱教君,袁小兰,等.辽东山区次生林主要树种种群结构和格局[J].北京林业大学学报,2007,29(1):12-18.
[22]霍常富,孙海龙,王政权,等.光照和氮营养对水曲柳苗木光合特性的影响[J].生态学杂志,2008,27(8):1255-1261.
[23]王学,肖治术,张知彬,等.昆虫种子捕食与蒙古栎种子产量和种子大小的关系[J].昆虫学报,2008,51(2):161-165.
[24]闫兴富.辽东栎的繁殖体及其与种群更新的关系[J].西北林学院学报,2008,23(5):103-107.
[25]程积民,赵凌平,程杰.子午岭60年辽东栎林种子质量与森林更新[J].北京林业大学学报,2009,31(2):10-16.
[26]许中旗,黄选瑞,徐成立,等.光照条件对蒙古栎幼苗生长及形态特征的影响[J].生态学报,2009,29(3):1121-1128.
[27]王凯,朱教君,于立忠,等.光环境对胡桃楸幼苗生长与光合作用的影响[J].应用生态学报,2010,21(4):821-826.
[28]马万里,罗菊春,荆涛,等.长白山林区核桃楸种群数量动态变化的研究[J].植物研究,2007,27(2):249-253.
猜你喜欢
林业科技情报(2021年1期)2021-04-14
果树资源学报(2020年3期)2020-05-13
新疆农业科学(2019年11期)2019-12-31
疯狂英语·初中天地(2019年10期)2019-10-23
农民致富之友(2019年11期)2019-05-23
森林工程(2018年4期)2018-08-04
农民致富之友(2017年23期)2018-01-02
学与玩(2017年4期)2017-02-16
天然产物研究与开发(2016年11期)2016-06-15
小天使·四年级语数英综合(2015年6期)2015-07-17
