
光照强度对毛竹竹笋到幼竹生长过程的影响1)
2012-09-18 01:27方飞燕白尚斌周国模王懿祥谢一鸣
东北林业大学学报 2012年3期
方飞燕 白尚斌 周国模 王懿祥 谢一鸣
(浙江农林大学,临安,311300)
光是一切生命的能量来源,光因子对植物的生长发育和分布产生重要的影响。绿色植物依赖叶绿素进行光合作用,将辐射能转化成具丰富能量的糖。光合作用与光照强度密切相关,在不同植物种中,植物光合能力对光照强度的反应是有差异的[1]。当生境中光照强度超过光补偿点(光合作用吸收CO2与呼吸作用放出CO2相等时的光照强度),植物才有净光合效率,才能正常的生长,反之,植物因有机物质无法得到有效的补充而不能正常生长。不同植物的光补偿点是不一样的。因此,环境中光照强度的变化不仅影响植物的生长发育,而且还影响植物群落的发育和演替[2-3]。
毛竹(Phyllostachys pubescens),又名楠竹(湖南、贵州)、孟宗竹(台湾),禾本科竹亚科刚竹属植物[4],是我国竹类植物中分布最广、产量最大、用途最多的一个优良竹种。由于其重要的经济价值,中国南方地区近年来大力发展竹业,使得毛竹林面积逐年增加,毛竹纯林化过程表现明显,蚕食了大量壳斗科、樟科和松科等乔木树种,灌木层和草本层的物种组成也发生着显著变化[5]。这一过程可能与毛竹蔓延引起林内光环境的变化有关[6-7]。毛竹自身独特的繁殖特性,即由竹笋长成幼竹,在这个过程中很大程度上是不直接需光的[8],且毛竹幼竹在林内的弱光条件也能很好地进行光合作用[9],生长极其迅速,短短的时间内就能达到林冠层。因此,为了解毛竹幼竹成竹过程中对光的需求,探讨毛竹成功扩张到周边森林群落的机制,文中采用人工遮光的方法来研究不同光照强度对毛竹幼竹生长的影响。
1 试验地概况
试验地位于浙江安吉递铺镇东侧高坞岭村,E119°22'~119°23',N30°36'~ 30°37°,属天目山山脉,海拔为 192 ~558 m,东西高,中部低[10],属亚热带季风气候,年均气温13.4℃,年降水量1100~1900 mm[11],无霜期 250 d,雨日 170 d,年平均日照1900 h,光照充足、气候温和、雨量充沛、四季分明。区域面积10.2 km2,其中毛竹林面积 1.1 hm2,是个山多田少的纯山丘。现植被多为毛竹林、松林及常绿阔叶次生林,保存良好,土壤大部分为山地红壤,土层较厚。
2 材料和方法
在浙江安吉高坞岭村,经踏查选择有代表性的毛竹林分3个(分别记为毛竹林A、毛竹林B、毛竹林C,基本概况见表1),各设置50 m×50 m的样地1个,于2010年3月末竹笋破土10~20 cm时,在每个样地内选取45株竹笋,分成3组,即每15株成一组作为试验对象,而所选竹笋的胸径、生长情况都要大致相同。对每个样地内的各组竹笋分别进行遮光处理试验。遮光处理分为全遮光和笋尖透光遮光,以自然条件做对照。全遮光是先把遮阳布做成锥台型,上底面直径为5 cm,下底面为30 cm,高为15 m,上底面为封闭,下底面为敞开。为了防止笋苗被压坏,采用3根12 m左右的杆子撑起遮阳布,以竹笋为中心距竹笋8 cm处对称地插上几根木头用以支开锥台内保持空气的流通;笋尖透光遮光是把遮阳布做成锥台型,上底面直径为5 cm,下底面为30 cm,高为15 m,上底面为不封闭并在上底面用铁丝扎成一小环(直径为5 cm),下底面为敞开,处理过程中,笋尖露出2 cm,其余操作与全遮光相同。
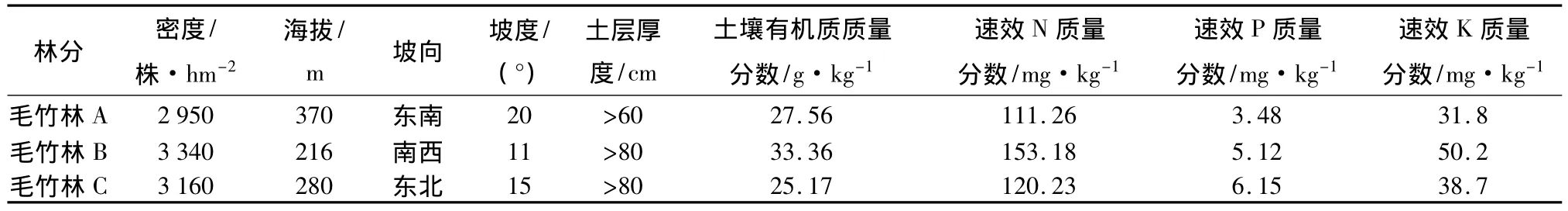
表1 安吉高坞岭村毛竹林分立地基本概况
2010年3月末遮光处理后到6月份(毛竹生长基本结束),每隔1 d进行各项指标(包括株高、胸径等)观测、测定。
为了对比遮光处理对幼竹成竹过程的影响,数据采用SPSS17.0中ANOVA进行差异分析,不同林分之间的对比采用EXCELL中描述统计中均值±标准误进行。绘图采用SigmaPlot 10.0软件进行。
2 结果与分析
2.1 遮光处理对毛竹竹高的影响
将不同遮光处理下的毛竹竹高按时间序列绘成图1。由图1可知,各样地中,从3月27日至4月4日,毛竹在全遮光处理下和在其他处理下一样,毛竹的竹高均在增长,但从4月4日至4月11日,毛竹的竹高不再增长,之后,毛竹停止生长,直至萎蔫死亡。而从其他处理看来,在3月末到4月末,毛竹的竹高增长速度相对缓慢,各处理和各样地间差别不明显;从5月份开始,竹高增长明显加快,5月下旬至6月初基本稳定,说明高生长过程基本结束。各处理和各样地之间的差异变化明显,这个阶段毛竹竹高由高到低的顺序均为:尖透、对照。这说明适当的遮光处理有利于毛竹竹高的增高,而过度的遮光如前面所说的全遮光处理,就对毛竹的竹高生长有抑制作用,导致毛竹死亡。而从方差分析表明,在不同样地中,在不同处理下,毛竹竹高有显著差异(P<0.05),尖透处理下比对照高12.99%;在不同样地中,毛竹竹高在尖透处理下比对照分别高16.07%、8.73%和 14.16%。
2.2 遮光处理对毛竹胸径的影响
由图2可知,在3月末到4月末,各处理和各样地间均无胸径,即幼竹的高度还未超过1.3 m,故未测得胸径数据;从4月底开始,胸径增长明显加快,到5月中旬达到稳定,不再增加,各处理之间的差异变化明显,这个阶段毛竹胸径由大到小的顺序均为:尖透、对照。这同样说明适当的遮光处理有利于毛竹胸径的提早生长,而全遮光处理下,毛竹未长到1.3 m就死亡,故未测定胸径数据。在毛竹林A、B和C 3块样地中,毛竹胸径在尖透处理下比对照分别高2.56%、2.94%和 1.78%,尖透处理下比对照高均值为2.43%。而从方差分析表明,在2种不同处理下,毛竹胸径差异不显著(P<0.05),这说明胸径的生长几乎不受光照强度的影响,而取决于竹笋的大小。但其生长过程受到光照强度的影响,适当遮光有利于提前进入胸径的生长。
2.3 遮光处理对毛竹径高比的影响
毛竹的径高比,即毛竹的胸径与竹高的比值,反映毛竹横向和纵向生长的关系。由表2可知,毛竹的径高比变化动态与毛竹的胸径变化动态相似,即在3月份到4月下旬,各处理和各样地间均无径高比;从4月底开始,径高比增长明显加快,样地A、C中尖透处理下,在5月8日到达顶峰,之后有所下降且达到固定值;而样地B中,在5月1日最大,尖透处理和对照的径高比分别为2.36和2.51,之后开始下降,到5月中旬后稳定不变。方差分析表明,在不同的样地中,各处理间的毛竹径高比无显著差异(P>0.05),说明不同处理方法对毛竹的径高比影响不明显。
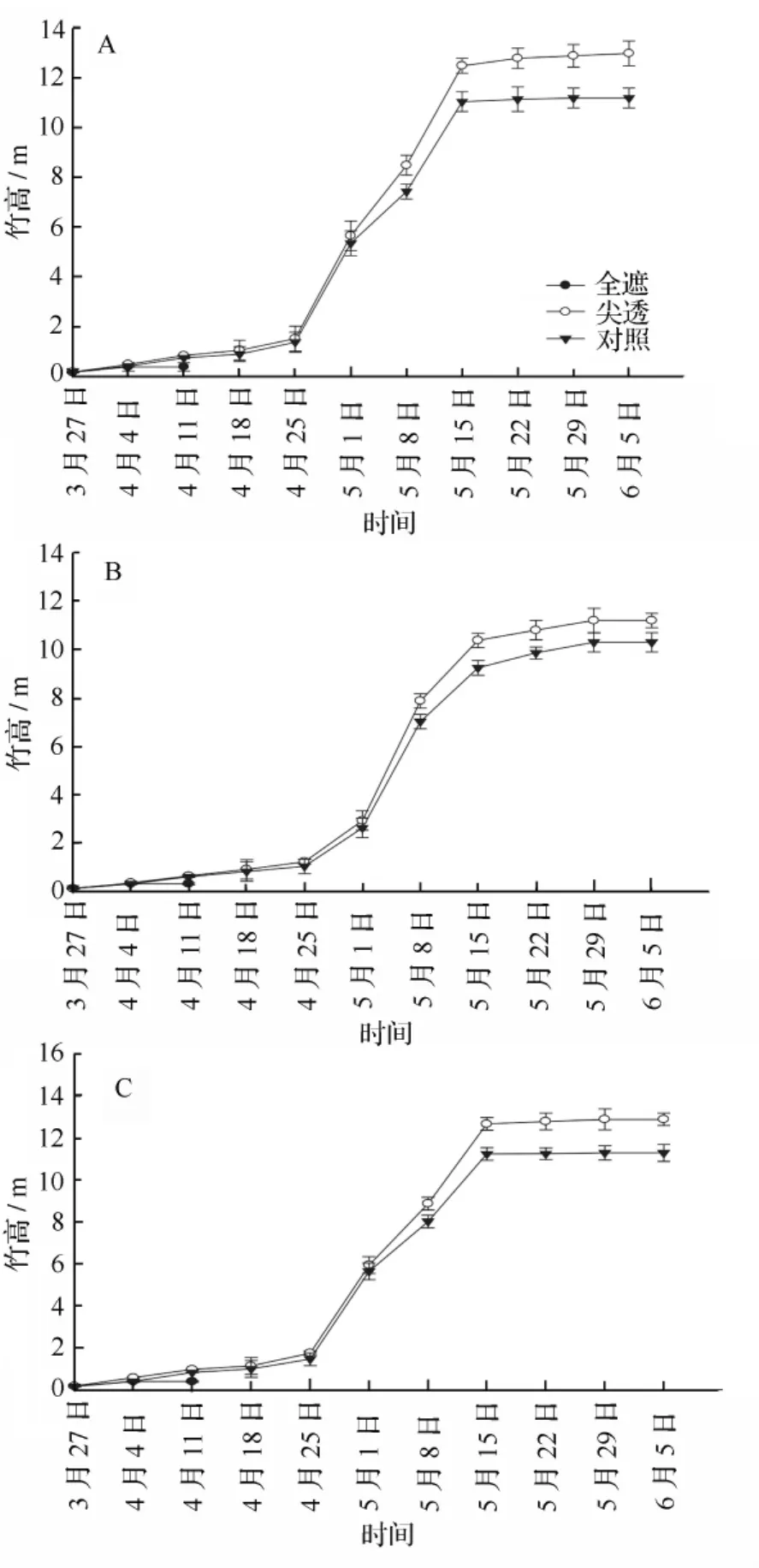
图1 不同遮光处理下不同样地毛竹的竹高动态变化(A、B、C为样地)
3 结论与讨论
植物生长状况与其所处的光环境密切相关[12]。在影响植物结构特性的生态因子(光、温度、水分及营养等)中,光照强度的作用最为显著[13],对植株的生长发育有重要意义。遮光影响光照强度大小,遮光度越高,光照强度越低。随着光照强度的降低,植物从生长上会产生一定的适应性,植物可以通过形态和生长调节,如增加株高、扩大叶面积、调整平面在空间上的方向等,主动适应环境,但当遮光严重时,会造成气孔部分关闭及生理功能减弱[14],生长发育就会受到严重阻碍,表现出黄花、瘦弱及死亡现象[15]。株高和胸径是毛竹两个重要生长指标。对毛竹进行全遮光处理,刚开始几天竹高在增长,但到达一定时候,缺少阳光导致毛竹死亡,说明由竹笋过渡到幼竹前尽管未展枝放叶也是离不开光的。而毛竹透尖遮光处理下,其竹高、胸径均大于自然条件下的毛竹竹高和胸径,特别是高生长有显著的差异,适当地遮光处理(笋尖透光处理)对毛竹的生长发育有更大的促进作用,竹高和胸径增长快。这也说明了毛竹从竹笋到幼竹这个生长过程中很大程度上是不直接需强光的[8],能够在弱光条件下很好地生长,这与杨迪蝶等[9]的研究结果相似。这个阶段与毛竹成竹生长需要较强的光照强度[16]是不同的,苗期毛竹叶片随着光合有效辐射(PA,R)逐渐增强(0~1800 μmol·m-2·s-1),其叶片叶绿素荧光参数非光化学猝灭也随之增加,表观光合电子传递速率则呈现先上升,后下降的趋势[17]。
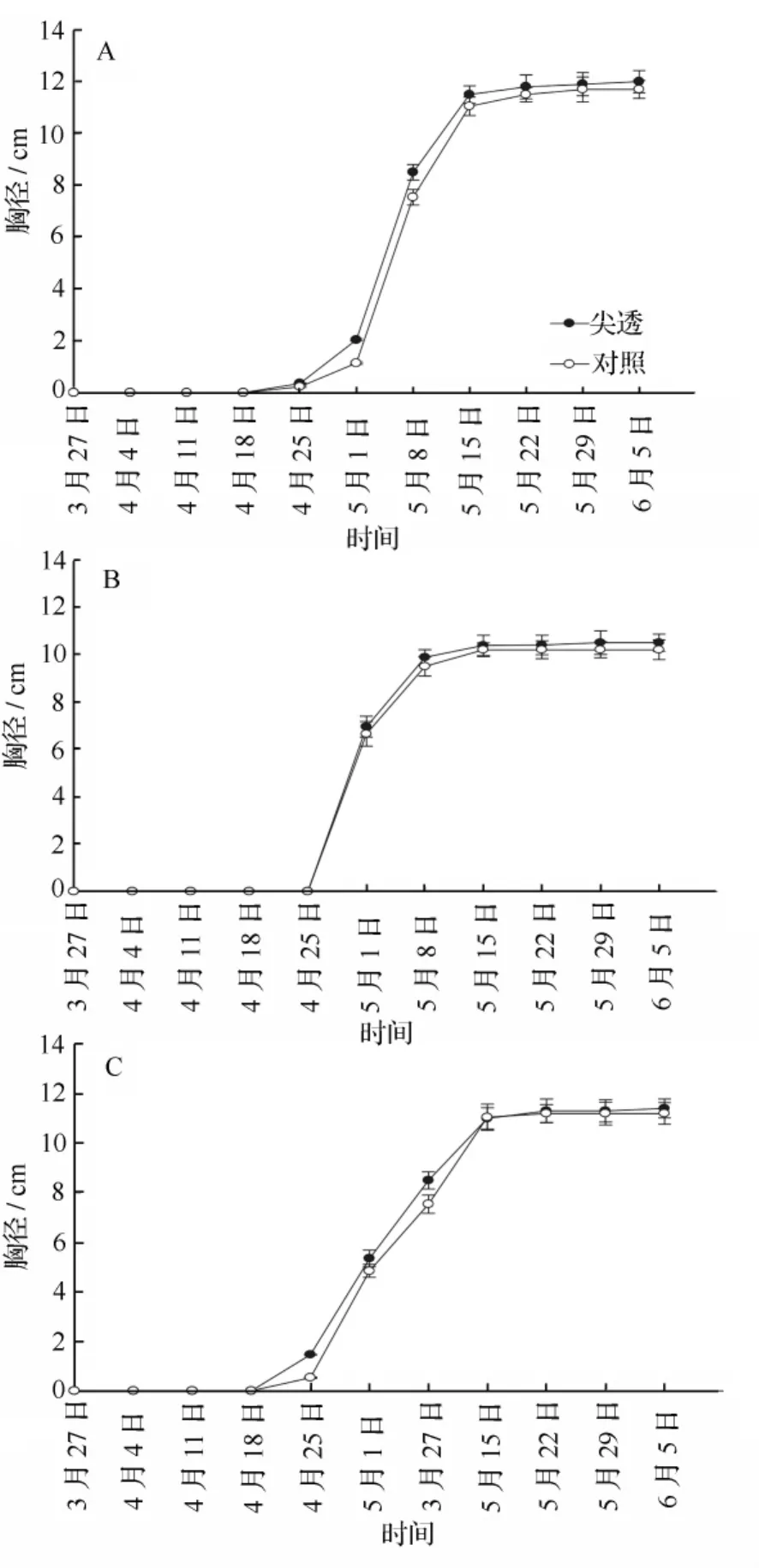
图2 不同遮光处理下不同样地毛竹的胸径动态变化(A、B、C为样地)
另外,光环境对植物生长的影响是独特和相当复杂的。在试验中遮光下改变的不只是毛竹的光照情况,其他因子如气温、土温、空气湿度、土壤湿度等可能会随之变化[12],同时采用遮光处理的材料、遮光程度和所选样地的不同,毛竹的生长是否受到影响,仍需作进一步的研究和探讨。

表2 不同遮光处理下不同样地毛竹的径高比动态变化
同时,不同立地条件对植物生长的影响也是复杂的。为了了解光环境的影响,文中选择了同一地区不同的3个毛竹林分,结果显示3个林分内毛竹幼竹生长虽有一定的差异,但遮光处理下,表现出非常的一致性,说明无论立地条件如何,遮光处理总会促进毛竹竹笋到幼竹生长过程。据刘烁等[18]研究表明,毛竹扩散形成的纯林内光照强度大于有毛竹扩散的针阔叶混交林和未有毛竹扩散的针阔叶混交林,这可能对扩散到针阔叶混交林内的毛竹竹鞭发笋成竹十分有利。这些为进一步了解毛竹成功扩张到周边林分的原因奠定了良好的基础。
[1]牛翠娟,娄安如.基础生态学[M].2版.北京:高等教育出版社,2007.
[2]Wiedemeyer W L.Biology of small juveniles of the holothurians Actinopyga echinites:Growth,mortality,and habitat preferences[J].Mar Biol,1994,120:81-93.
[3]张硕,陈勇,孙满昌.光强对刺参行为特性和人工礁模型集参效果的影响[J].中国水产科学,2006,13(1):20-27.
[4]商业部土产杂品局.毛竹经营知识[M].北京:中国财政经济出版社,1982:1.
[5]郑成洋,何建源,罗春茂,等.不同经营强度条件下毛竹林植物物种多样性的变化[J].生态学杂志,2003,22(6):1-6.
[6]Nakatsubo T,Suzaki T.Ecological studies on the vegetation and management of the Yahagi River I.The impact of bamboo cutting on the vegetation in Otsuridoba area[J].Rep Yahagi River Inst,1998(2):113-127.
[7]Suzaki T,Nakatsubo T.Impact of the bamboo Phyllostachys bambusoides on the light environment and plant communities on riverbanks[J].J For Res,2001(2):81-86.
[8]李睿,维尔格M J A,钟章成.施肥对毛竹(Phyllostachys pubescens)竹笋生长的影响[J].植物生态学报,1997,21(1):19-26.
[9]杨迪蝶,黄启民.毛竹适生苗生物量及光合速率的研究[J].林业科学研究,1990,3(5):461-465.
[10]邱栩文,王莉红,王卫军.生态工程在山区生态村建设规划中的应用:以安吉县高坞岭村为例[J].农机化研究,2006,5(5):200-203.
[11]潘春霞,李雪涛,吕玉龙.安吉县毛竹资源及其生物量研究[J].浙江林业科技,2010,30(1):82-84.
[12]陈菊艳,杨远庆.遮光对野扇花生长特性和生理指标的影响[J].西北植物学报,2010,30(8):1646-1652.
[13]Bjorkman O,Holmgren P.Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitat[J].Physoil Plantarum,1963,16(4):889-914.
[14]Titlyanov E A,Titlyanova T V,Yamazato K.Photo-acclimation dynamics of the coral Stylophora pistillata to low and extremely low light[J].J Exp Mar Biol Ecol,2001,263:211-225.
[15]迟伟,王荣富.遮光条件下的草莓光合生理特性[J].江苏农业学报,2001,17(1):62-63.
[16]陈建华,毛丹,马宗艳,等.毛竹叶片的生理特性[J].中南林学院学报,2006,26(6):76-80.
[17]郑波,高志民.毛竹苗期NPQ和ETR特征初步研究[J].世界竹藤通讯,2011,9(1):12-14.
[18]刘烁,周国模,白尚斌.基于光照强度变化的毛竹扩张对杉木影响的探讨[J].浙江农林大学学报,2011,28(4):550-554.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
乡村科技(2018年8期)2018-06-27
小天使·一年级语数英综合(2018年4期)2018-06-22
小天使·一年级语数英综合(2017年11期)2017-12-05
小布老虎(2017年4期)2017-08-10
小学生作文(低年级适用)(2017年3期)2017-07-06
湖南林业科技(2017年1期)2017-02-06
