
中国伞形科前胡属果实表面微形态特征及分类学意义
2012-09-06 01:51李美芝宋春凤刘启新
植物资源与环境学报 2012年2期
李美芝,宋春凤,刘启新
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏 南京 210014〕
中国伞形科前胡属果实表面微形态特征及分类学意义
李美芝,宋春凤,刘启新①
〔江苏省·中国科学院植物研究所(南京中山植物园),江苏 南京 210014〕
采用扫描电镜方法对中国伞形科(Apiaceae)前胡属(PeucedanumL.)22种2变种的果实表面微形态特征进行了观察和描述。结果显示:供试种类在果实棱槽细胞平滑度和细胞轮廓可见度、蜡质纹饰类型、细胞表面突起的有无及类型、表皮毛的有无及类型及其纹饰类型、表皮分泌物的有无及类型等方面有明显差异,具有丰富的种间多样性。根据这些特征可将供试种类分为4种类型。类型Ⅰ:棱槽细胞粗糙多毛—细胞轮廓不可见—簇状蜡质纹饰发达,包含滨海前胡(P.japonicumThunb.)、天竺山前胡(P.ampliatumK.T.Fu)、华北前胡(P.harrysmithiiFedde ex Wolff)及其2变种、长前胡(P.turgeniifoliumWolff)、华山前胡(P.ledebourielloidesK.T.Fu)和泰山前胡〔P.wawrae(Wolff)Su ex Sheh〕;类型Ⅱ:棱槽细胞平滑无毛—细胞轮廓不可见或凹陷—条形蜡质纹饰发达或明显,包含芷叶前胡(P.angelicoidesWolff ex Kretschm.)、竹节前胡(P.dielsianumFedde ex Wolff)、南川前胡〔P.dissolutum(Diels) Wolff〕、红前胡(P.rubricauleShan et Sheh)、细裂前胡(P.macilentumFranch.)、前胡(P.praeruptorumDunn)、华中前胡(P.medicumDunn)、台湾前胡(P.formosanumHayata)、南岭前胡(P.longshengenseShan et Sheh)、会泽前胡(P.acauleShan et Sheh)和马山前胡(P.mashanenseShan et Sheh);类型Ⅲ:棱槽细胞有稀疏毛—细胞轮廓凹陷可见或不明显—蜡质纹饰几无,包含北京前胡(P.caespitosumWolff)、草原前胡(P.stepposumHuang)、毛前胡(P.pubescensHand.-Mazz.)和刺尖前胡(P.elegansKomarov);类型Ⅳ:棱槽细胞不平无毛—细胞轮廓呈近圆形凸起—蜡质纹饰为微波状长条形,仅包含石防风〔P.terebinthaceum(Fisch.ex Trevir.)Fisch.ex Turcz.〕1种。结合外部形态特征以及地理分布对各类型种类的分类关系进行了讨论,并对会泽前胡、马山前胡和石防风的特殊分类地位进行了分析,明确了果实表面微形态特征在前胡属中的分类学意义。
前胡属;伞形科;果实;扫描电镜;微形态特征;分类学
前胡属(PeucedanumL.)是林奈于1753年以P.officinaleL.为模式种建立的,隶属于伞形科(Apiaceae)芹亚科(Apioideae)前胡族(Peucedaneae)阿魏亚族(Ferulinea)[1]。全世界约有120多种,主要分布于欧洲、亚洲和非洲;中国约有40种,其中33种为特有种,全国各地均有分布[2]。前胡属在伞形科中是较大的属,种类来源复杂、形态变异较大、属的界线模糊[3-8]。对于该属的认识目前有广义和狭义2种观点:前者如Tutin等[5]将ImperatoriaL.和TommasiniaBertol.的一些种类组合到前胡属中;后者如Reduron等[6]将意大利分布的前胡属种类分为9个属(包括TommasiniaBertol.、HolandreaReduron,Charpin et Pimenov、CervariaWolff、ImperatoriaL.、OreoselinumMill.、PeucedanumL.s.str.、XanthoselinumSchur、ThyseliumRaf.和PteroselinumRchb.),Pimenov等[4]认为该属只有欧亚地区的8~10个种,Coulter等[7]将北美原属于前胡属的40个种类移入到LomatiumRaf.中,Pieter等[8]基于分子系统学研究结果将前胡属的非洲种类与欧亚地区种类进行分离,等等。而且迄今为止前胡属属下也没有一个理想的分类系统,因而,前胡属是一个分类较困难的类群,也是伞形科研究的难点。
单人骅等[9]根据萼齿、苞片和果实侧棱等形态特征将中国分布的前胡属40余种分为5组,即:刺尖前胡组(Sect.ElegantiaGorov.)、多小苞片组(Sect.BracteolataSheh et Shan)、山亮蛇床组〔Sect.Oreoselinum(Adans.)Rchb.〕、近亮蛇床组(Sect.SelinoidesDC.)和前胡组(Sect.PeucedanumSynops.)。2005年佘孟兰等[2]虽然对该属又做了修订,共计40种,但未建立属下分类系统。
国外学者对前胡属开展了染色体核型、植物化学成分、免疫系统学和分子系统学等方面的研究[10-18],但涉及中国产的种类较少,且中国特有种尚未涉及。国内学者对前胡属的研究主要集中在孢粉学、血清分类学和同工酶变异等方面[19-23]。由于结构与性状的独特性和稳定性,果实已成为伞形科系统分类的重要依据[24],但有关伞形科果实表面微形态特征的研究报道并不多[25-28],更无前胡属果实表面微形态特征的研究报道。为此,作者采用扫描电子显微镜对前胡属22种2变种的果实表面微形态特征进行观察,以期讨论前胡属果实表面微形态特征的分类学意义,探讨属下分类关系,为前胡属的系统分类研究提供果实微形态学依据。
1 材料和方法
1.1 材料
前胡属分布于全国各地[2],多数种类分布区域狭窄,加之环境变化和人为干扰,有些种类生境已遭破坏,野外取材较为困难。本文采用除前胡组(中国仅分布1种)外的其余4组22种2变种作为研究对象。供试材料为取自野生植株或腊叶标本的成熟果实。每种类各取果实3~5粒。供试种类及其来源和凭证标本见表1。
1.2 方法
选取成熟果实(分生果),用无水乙醇超声波振荡清洗1~2 min,自然风干。将果实黏置于导电胶上,置于离子溅射仪中喷金,之后将其移入JSM-5610LV型扫描电镜下观察拍照。
在对果实的背棱、棱槽、侧棱和合生面等部位进行观察比较后,发现各部位微形态特征都具有一定的种间差异性,但棱槽部位的差异较为明显和稳定,因此,作者重点观察分生果果棱之间的棱槽部位。观察内容为棱槽表面细胞平滑度及可见度、蜡质纹饰类型、表面突起和表皮毛类型及密度、表皮分泌物类型及多寡。参照有关文献[25-26,29]对果实表面微形态特征进行描述。
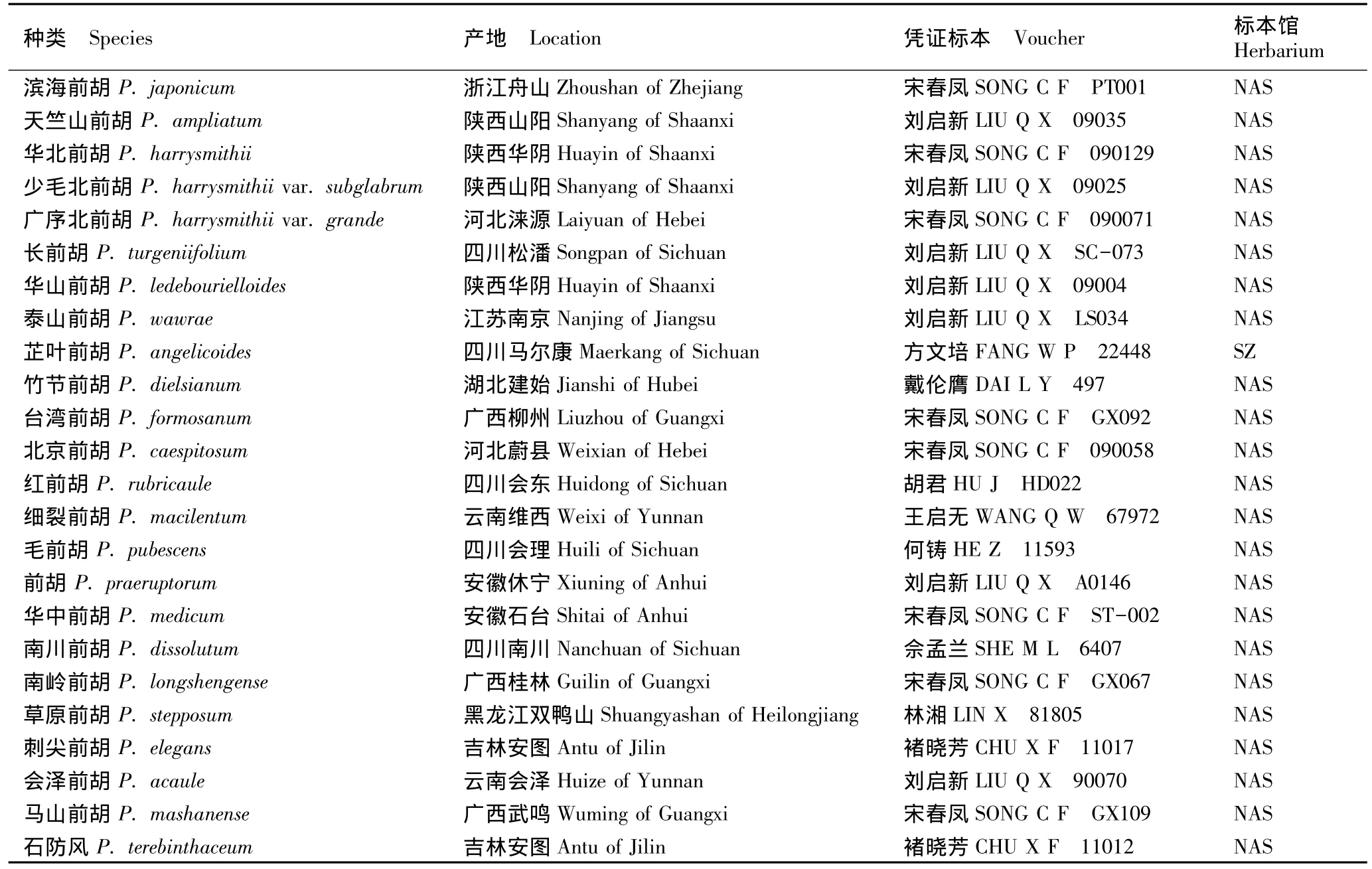
表1 供试前胡属种类的产地及凭证标本Table 1 Location and voucher of Peucedanum L.species tested
2 结果和分析
前胡属果实(本文指分生果)为椭圆形、长圆形或近圆形,背部压扁,中棱和背棱丝线状,侧棱为较厚的窄翅。经作者测量,前胡属果实的平均长度为3.1~8.0 mm,平均宽度为1.2~3.8 mm。在扫描电镜下,其棱槽细胞、蜡质纹饰、表面突起、表皮毛和表皮分泌物等特征呈现出丰富的多样性(见表2,图版Ⅰ和Ⅱ)。
2.1 外表皮细胞
供试的22种2变种果实棱槽部位外表皮细胞平滑度可分为粗糙(具突起或表皮毛)、微光滑、凹陷和凸起4类。细胞表面粗糙的种类有滨海前胡(P.japonicumThunb.)、天竺山前胡(P.ampliatumK.T.Fu)、华北前胡(P.harrysmithiiFedde ex Wolff)、少毛北前胡〔P.harrysmithiivar.subglabrum(Shan et Sheh)Shan et Sheh〕、广序北前胡〔P.harrysmithiivar.grande(K.T.Fu)Shan et Sheh〕、长前胡(P.turgeniifoliumWolff)、华山前胡(P.ledebourielloidesK.T.Fu)和泰山前胡〔P.wawrae(Wolff)Su ex Sheh〕(图版Ⅰ-1,4~11);细胞表面光滑的种类有芷叶前胡(P.angelicoidesWolff ex Kretschm.)、竹节前胡(P.dielsianumFedde ex Wolff)、台湾前胡(P.formosanumHayata)、会泽前胡(P.acauleShan et Sheh)和马山前胡(P.mashanenseShan et Sheh)(图版Ⅰ-2,12~14和图版Ⅱ-10~13);细胞表面凹陷的种类有北京前胡(P.caespitosumWolff)、红前胡(P.rubricauleShan et Sheh)、细裂前胡(P.macilentumFranch.)、毛前胡(P.pubescensHand.-Mazz.)、前胡(P.praeruptorumDunn)、华中前胡(P.medicumDunn)、南川前胡〔P.dissolutum(Diels)Wolff〕、南岭前胡(P.longshengenseShan et Sheh)、草原前胡(P.stepposumHuang)和刺尖前胡(P.elegansKomarov)(图版Ⅰ-3,15和图版Ⅱ-1~9);细胞表面呈凸起状的种类仅石防风〔P.terebinthaceum(Fisch.ex Trevir.)Fisch.ex Turcz.〕1种(图版Ⅱ-14,15)。
从表皮细胞形状及可见度来看,供试多数种类果实棱槽部位外表皮细胞形状不可见或隐约可见;仅北京前胡、红前胡、细裂前胡和毛前胡等少数种类外表皮细胞为四至六边形(图版Ⅰ-15和图版Ⅱ-1~3)。
2.2 蜡质纹饰类型
供试的22种2变种果实表面的蜡质纹饰类型可分为条纹状、簇状(围绕突起和表皮毛呈簇状,走向几乎不可见)、网状和束状,多数种类具条纹状和簇状纹饰,且纹饰的长短、疏密和平直度等有一定差别。其中,具有条纹状纹饰的种类有芷叶前胡、竹节前胡、台湾前胡、北京前胡、红前胡、细裂前胡、毛前胡、前胡、华中前胡和石防风(图版Ⅰ-12~15和图版Ⅱ-1~5,14,15);具有簇状纹饰的种类有滨海前胡、天竺山前胡、华北前胡、少毛北前胡、广序北前胡、长前胡、华山前胡和泰山前胡(图版Ⅰ-4~11);会泽前胡具有束状纹饰(图版Ⅱ-10,11);马山前胡具有网状纹饰(图版Ⅱ-12,13);而南川前胡、南岭前胡、草原前胡和刺尖前胡则无蜡质纹饰(图版Ⅱ-6~9)。
2.3 细胞表面突起
供试的部分种类棱槽细胞表面没有突起,多数种类棱槽细胞表面有瘤状、刺状或疣状等不同类型突起。竹节前胡、台湾前胡、细裂前胡、前胡、华中前胡、南川前胡、马山前胡和石防风无突起(图版Ⅰ-13,14和图版Ⅱ-2,4~6,12~15);南岭前胡、草原前胡和刺尖前胡具刺状突起(图版Ⅱ-7~9);毛前胡和会泽前胡具疣状突起(图版Ⅱ-3,10,11);其他种类均具瘤状突起。
2.4 表皮毛类型
供试的部分种类具囊状、长囊状、尖囊状或带状的表皮毛,包含的种类有天竺山前胡、华北前胡、少毛北前胡、华山前胡、毛前胡、滨海前胡、广序北前胡、长前胡、泰山前胡和北京前胡(图版Ⅰ-4~11,15和图版Ⅱ-3),其中前5个种的表皮毛较密集,后5个种的表皮毛较稀疏,其余种类均无表皮毛。
表皮毛上具有蜡质纹饰,除毛前胡果实表皮毛上的纹饰几乎不可见外,其他种类的表皮毛均具有拟脑纹状的蜡质纹饰。
2.5 表皮分泌物
供试的22种2变种的果实表面分泌物的有无、多寡及类型也有一定的差别。具颗粒状分泌物的种类有滨海前胡、天竺山前胡、长前胡、芷叶前胡、台湾前胡、南川前胡、南岭前胡、草原前胡、会泽前胡和马山前胡(同时具丝状分泌物)(图版Ⅰ-4,5,9,12,14和图版Ⅱ-6~8,10~13);华中前胡具丝状分泌物(图版Ⅱ-5);刺尖前胡具盘状分泌物(图版Ⅱ-9);其余种类无分泌物。
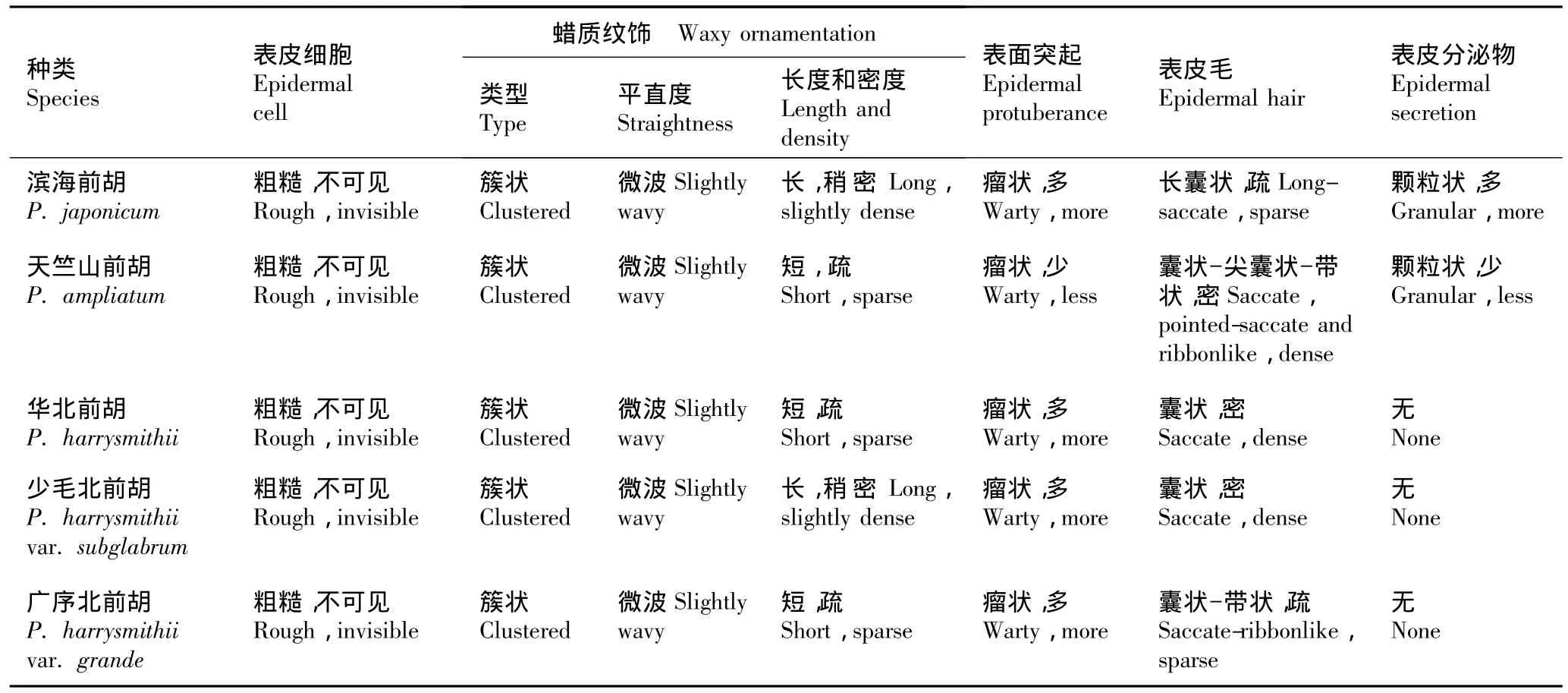
表2 前胡属种类果实棱槽部位表面微形态特征Table 2 Surface micromorphological features at vallecular part of mericarp of Peucedanum L.species

续表2Table 2 (Continued)
3 讨论
3.1 前胡属果实表面微形态的类型和归类
供试种类的果实表面微形态特征差异较大,且以棱槽部位的变异最为丰富。根据果实棱槽部位的细胞平滑度、细胞轮廓和蜡质纹饰等特征,可将供试的22种2变种划分为如下几种类型:
类型Ⅰ:棱槽细胞粗糙多毛—细胞轮廓不可见—簇状蜡质纹饰发达。该类果实的棱槽表面具有明显的瘤状突起和表皮毛,数量较多;整个果体表面外果皮细胞轮廓常不可见;蜡质纹饰在整个棱槽都比较发达且变化多样,除了在细胞表面呈近平行分布外,有2个突出的特殊分布式样:1)以瘤状突起和表皮毛为基点,蜡质纹饰在其基部及周围呈簇状分布,纹理呈放射状,并且受其影响,蜡质纹饰稍弯曲,有时走向不明显;2)毛被上的蜡质纹理较短,高度密集和弯曲,呈拟脑状,布满整个毛被,并向顶部汇集。该类型包含的种类有滨海前胡、天竺山前胡、华北前胡、少毛北前胡、广序北前胡、长前胡、华山前胡和泰山前胡。
类型Ⅱ:棱槽细胞平滑无毛—细胞轮廓不可见或凹陷—条形蜡质纹饰发达或明显。该类果实的棱槽表面比较平滑,没有瘤状突起或表皮毛;蜡质纹饰为条形,纹理清晰可辨,走向一般近平行,并且在整个棱槽中呈不同程度的均匀分布;因蜡质纹理较密细胞轮廓不可见,或因为蜡质纹理较疏而可见,并呈一定程度的凹陷。该类型包含的种类有芷叶前胡、竹节前胡、南川前胡、红前胡、细裂前胡、前胡、华中前胡、台湾前胡、南岭前胡、会泽前胡和马山前胡。各种类间的区别在于蜡质纹饰的长短、疏密和平直度等。
类型Ⅲ:棱槽细胞有稀疏毛—细胞轮廓凹陷可见或不明显—蜡质纹饰几无。该类型果实的棱槽细胞不仅表皮毛很少,而且表皮毛上的蜡质纹理也很少,棱槽细胞表面基本无蜡质纹饰。该类型包含的种类有北京前胡、草原前胡、毛前胡和刺尖前胡。
类型Ⅳ:棱槽细胞不平无毛—细胞轮廓呈近圆形凸起—蜡质纹饰为微波状长条形。该类型与上述各类型的最大不同点是细胞明显向外隆起,呈泡状。该类型仅石防风1种。
总体而言,除石防风果实的棱槽细胞外侧壁向外隆起外,大多数种类棱槽细胞外侧壁的轮廓或不可见或向下凹陷,因此,后者基本为供试种类的共性特征。但不同种类在棱槽细胞平滑度、细胞轮廓可见度、蜡质纹饰类型和突起、表皮毛和分泌物的有无及类型等方面表现出较大的差异,可以为前胡属的分类提供重要的微形态学依据。
3.2 不同果实微形态类型与前胡属种间关系的讨论
前胡属果实表面微形态特征比较复杂,但根据上述类型进行划分和归类,对研究宏观外部形态性状比较相近的种类有重要的参考价值。此外,虽然用果实表面微形态特征对种类进行的归类与经典分类中的分组有一定差异[9],但是据此探讨属内种间关系仍具有一定的意义。
3.2.1 类型Ⅰ中的种间关系分析在类型Ⅰ包含的前胡属种类中,仅滨海前胡分布到江苏、浙江、福建和台湾的沿海海滨,其他种类均分布于长江以北地区,它们主要位于华北—西北—华东地区,并以秦岭为中心向西到西北地区的岷山山系,向东到沿海,向东北到太行山脉。其中以太行山、秦岭为界,两大山系及其以北的黄土高原地区分布着华北前胡;秦岭西段的秦、巴、岷交汇地区分布着长前胡,华北前胡在此地区也有分布;太行山北段至秦岭东段北坡分布着广序北前胡;秦岭东段的南坡分布有特产陕西山阳的天竺山前胡,秦岭东段的南坡和河南西部山地分布着少毛北前胡,此外还有特产于秦岭山区的华山前胡。秦岭、太行山以南地区的种类有泰山前胡和滨海前胡,并且前者零星分布于山东至长江以北的低山丘陵地区,后者分布于我国东部沿海滩地和近海岛屿[23]。可见该类型的种类在地理分布上有各自的范围,同时又相互交叉。地理分布上的特点也验证了外部形态特征及果实表面微形态特征的相似性,这也说明相对于前胡属其他类型种类,这一类型的种间亲缘关系较近。
1)滨海前胡与天竺山前胡的关系:滨海前胡的果实表皮毛较稀疏,蜡质纹饰在突起和表皮毛处呈簇状分布,但总体仍可见微波状稍密集的条纹,并且瘤状突起和颗粒状分泌物较多;天竺山前胡的果实表皮毛较前者更为密集且呈囊状、尖囊状和带状等多种形态,纹饰呈簇状,但是由于突起和表皮毛密集,纹饰走向已不可见,而且瘤状突起及颗粒状分泌物较少。从地理分布来看,前者分布于沿海地区,后者分布于秦岭山区。但从生境来看,二者都生长于石缝中,区别在于前者生于基岩海岸石缝中,而后者生于内陆的山顶崖壁上。从宏观形态来看,二者有许多相近的特征,如根茎粗壮;叶片光滑无毛,一至二回三出式羽状分裂,末回裂片宽大,且叶缘具宽大圆齿;花序梗粗壮并被毛;分生果长圆形且背部压扁明显。可见,这2个种在果实微形态和宏观形态方面都具有较多的相似性,显示出较为密切的亲缘关系。
2)华北前胡及其变种与天竺山前胡的关系:华北前胡果实表面微形态特征与天竺山前胡相似,但其簇状纹饰稀疏,仅有囊状表皮毛,无分泌物;少毛北前胡和广序北前胡的果实微形态特征与华北前胡十分相似,区别在于2变种的簇状纹饰较华北前胡更为密集,且后者具长表皮毛。宏观形态上,天竺山前胡果实形状及毛被特征与华北前胡特别是广序北前胡有相似之处;少毛北前胡植株各部分的毛被较华北前胡少,广序北前胡的花序比华北前胡大。因此,结合果实微形态和宏观性状可知它们的关系较近。
3)长前胡与华北前胡的关系:长前胡果实表面微形态特征与华北前胡相似,区别仅在于其具有较密集的簇状纹饰、表皮毛较少、仅有少量颗粒状分泌物。二者在宏观形态上也相近,分生果均为卵状椭圆形、表面被毛、背棱线状突起。
4)泰山前胡与华山前胡的关系:华山前胡果实的最大特点是表皮毛囊状且呈簇状密布于果体表面;泰山前胡与华山前胡果实微形态特征相似,特殊之处在于其棱槽处的瘤状突起生于凹陷的细胞中。二者的宏观形态特征也较相近,共同特点是茎枝细挺且多分枝、复伞形花序分枝多且常二叉式展开、花序梗短、伞形花序小而多、伞幅少、萼齿显著。
3.2.2 类型Ⅱ中的种间关系分析类型Ⅱ包含的种类均分布于长江以南地区。从地理分布看,主要位于西南—华中—华东和华南地区,从青藏高原的横断山以东一直分布到南岭山系的武夷山脉。
1)芷叶前胡、竹节前胡和台湾前胡的关系:三者均为多小苞片组种类,果实微形态特征均为棱槽细胞表面光滑,细胞形状不可见,蜡质纹饰较多且为条纹状。芷叶前胡的条纹微波状,较短且稀疏,偶见瘤状突起,无分泌物;竹节前胡的条纹比前者长且密集,无突起和分泌物;台湾前胡的蜡质纹饰为近平直、较长的粗条纹。宏观形态上,三者均具有萼齿细小不显著、总苞片无或极少、小苞片多数、花柱细短弯曲、花柱基圆锥形等特征。由此可见,这3个种类在果实表面微形态特征和宏观形态上具有较大的共性。
竹节前胡分布于四川盆地东南边缘及渝鄂交界区;芷叶前胡分布于云贵高原及四川盆地西部地区;台湾前胡分布于中国东南部。从地理分布上看,竹节前胡与芷叶前胡的关系较台湾前胡近,而台湾前胡与南岭前胡(分布于中南、西南地区)在地理分布上存在交叉,且宏观形态相似,二者植株自上部分枝,基生叶具长柄,叶片轮廓三角状且宽大,花序梗具毛,花枝短且弯曲,花柱基圆锥形。这与依据果实表面微形态进行的归类不一致,其亲缘关系还需借助其他学科手段进一步研究探讨。
2)红前胡与细裂前胡的关系:这2个种类均隶属于山亮蛇床组,它们的棱槽细胞均凹陷,细胞轮廓可见且多为四至六边形,蜡质纹饰为条纹。但红前胡的条状纹饰较长而密集,表面突起呈疣状;细裂前胡细胞形状为清晰的四至六边形,纹饰为微波状及短密的条纹。从宏观形态看,二者的叶片均为多回羽状全裂,末回裂片线形,顶端具小尖头;萼齿显著,花瓣倒卵形,顶端凹陷;果实侧棱较狭窄而厚。二者均分布于西南地区,生长于海拔2 000~3 000 m的山坡上,只是红前胡分布于川滇交界处,细裂前胡分布于滇西。由此可见这2个种类关系较为密切。
3)前胡、华中前胡、南川前胡与南岭前胡的关系:它们的果实表面细胞形状隐约可见,条状纹饰少或无。前胡的细胞形状较其他种类清晰,条状纹饰较平直,长且稀疏;华中前胡和南川前胡的条状纹饰极稀疏,具丝状分泌物,但后者细胞形状隐约可见,为菱形;南岭前胡无纹饰,并具少量刺状突起。前胡分布于华东、华中及西部地区;华中前胡主要分布在华中、华南地区及四川等地;南川前胡主要分布于与云贵高原东接壤的川东南地区;南岭前胡分布于中南、西南地区,与前胡在叶片轮廓、毛被、萼齿等形态特征上具相似性。4个种都为中国特有种,前胡的分布区域与其他种类相互交接,可能在地理分布上存在着彼此替代的现象;华中前胡与南川前胡均具有萼齿发达,总苞早落等特征,在宏观形态上较前者更为相似,二者分布于云贵高原东北山地及华中地区湘鄂西部,这一地区被认为是中国特有属种演化发展的次中心。因此,结合地理分布特点和酯酶同工酶研究结果[23],认为华中前胡与南川前胡亲缘关系更近。
4)会泽前胡和马山前胡与该类型中其他种类的关系:会泽前胡和马山前胡与上述种类有明显的不同,其中最大的差异是二者具有极其发达的蜡质纹饰。会泽前胡的蜡质纹理较直且多成束分布;马山前胡的蜡质纹理多弯曲且呈交织状。在地理分布和经典分类的分组处理上二者也明显有异。
3.2.3 类型Ⅲ中的种间关系分析该类型的种类有北京前胡、草原前胡、毛前胡和刺尖前胡。
1)北京前胡与草原前胡的关系:二者的相同点是果实表面有少量蜡质纹饰,区别仅在于草原前胡具颗粒状分泌物,北京前胡有稀疏瘤状突起和尖囊状表皮毛。二者的外部形态高度相似,叶片均为三回羽状全裂,末回裂片线状、全缘、末端具小尖头;萼齿细小且不明显;复伞形花序略呈伞房状;果实椭圆形且光滑无毛。草原前胡分布于东经124°~125°一线的草原地区;北京前胡分布于河北西北部及北京山区,生长于海拔1 300~2 500 m的山坡石缝间。二者都属于分布于华北地区的种类,具有较近的亲缘关系。
2)刺尖前胡和毛前胡的关系:二者果实表面微形态的共同点是基本没有蜡质纹理,但二者差异较大。前者具皮刺突起,分泌物为盘状;分布于东北地区;叶片均为三回羽状全裂,末回裂片线状、全缘、末端具明显刺尖。后者具囊状短硬毛和少量瘤状突起,毛上的纹饰几乎不可见;分布于川南和滇北地区;叶片二至三回三出式分裂,末回裂片宽卵形,边缘有大而疏的锯齿和圆锯齿。可见,二者的差异较大。
3.3 会泽前胡、马山前胡和石防风分类地位的讨论
会泽前胡果实表面蜡质纹饰呈平直、细密的束状,细胞表面多具疣状突起,与其他种类果实表面微形态特征有明显差异。会泽前胡特产于云南东北部,生于海拔约3 500 m的草地,为西南高山特化种;株高仅5~10 cm,是国产前胡属中株高最矮的1种;植株无茎,丛生,叶片很小,自根茎处着生,三至四回羽状全裂,末回裂片线形、急尖;复伞花序多数,从根茎处抽出呈丛生状。惠红等[22]根据该种的外部形态、地理分布及血清分类学研究结果,建议将其从刺尖前胡组中独立出来。本研究结果也支持这一观点。
马山前胡果实棱槽细胞表面微光滑,蜡质纹饰具皱波状、短密的网状纹饰并呈交织状,分泌物呈丝状、颗粒状,特别是其果实表面具较密集的气孔,在背棱两侧呈宽带状排布,并低于蜡质纹理层,可见其果实微形态特征与上述种类差异较大。马山前胡是分布于广西南岭以南石灰岩地区的特有种,生于低山灌丛或阴处石缝中;植株木质化程度较高,茎自下部开始分枝,呈二歧式;叶片革质,末回裂片较宽。其独特的形态特征可能是对石灰岩地区水分缺乏的特殊生长环境的适应结果。相对于前胡属的其他种类,马山前胡果实微形态和外部形态特征较独特,结合血清分类学和同工酶研究结果[22-23],认为马山前胡是国产前胡属中具有独特演化地位的种类。
石防风细胞表面凸起,细胞近圆形,纹饰为微波、长、密的条纹,与其他种类的果实表面微形态特征差异较大。该种分布于我国河北、内蒙古及东北三省全境,在俄罗斯、日本和朝鲜也有分布,是东亚植物地区向环北方植物地区过渡的种类。通过血清分类学研究,刘启新等[21]认为该种与国产前胡属其他种类间有一定差异,应自成一类。而其特殊的果实微形态特征也进一步说明其独特的演化地位。但该种是否应独立为新组或从前胡属中分出,仍需进一步研究确定。
[1]DRUDE C G O.Umbelliferae[M]∥ENGLER A,PRANTL K.Die Naturlichen Pflanzenfamilien:3(8).Leipzig:Verlagvon Wilhelm Engelmann,1898:63-250.
[2]WU Z Y,RAVEN P H.Flora of China:Vol.14[M].Beijing: Science Press,2005:182-192.
[3]BURTT B L.Umbelliferae of Southern Africa:an introduction and annotated checklist[J].Edinburgh Journal of Botany,1991,48: 133-282.
[4]PIMENOV M G,LEONOV M V.The Genera of the Umbelliferae[M].Chicago:University of Chicago Press,1993.
[5]TUTIN T G,HEYWOOD V H,BURGES N A,et al.Flora Europaea:Vol.2[M].Cambridge:Cambridge University Press,1968.
[6]REDURON J P,CHARPIN A,PIMENOV M G.Contributionala nomenclature générique des Apiaceae(Ombelliféres)[J].Journal Botanique de la Société Botanique de France,1997,1:91-104.
[7]COULTER J M,ROSE J N.Monograph of the North American Umbelliferae[J].Contributions from the United States National Herbarium,1900,7:1-256.
[8]PIETER J D W,ANTHONY R M,PHEPHU N.A new generic classification for African Peucedanoid species(Apiaceae)[J].Taxon,2008,57(2):347-364.
[9]中国科学院中国植物志编辑委员会.中国植物志:第五十五卷第三分册[M].北京:科学出版社,1992:127-175.
[10]SOLOV N M.Karyotaxonomic study of some species ofPeucedanum(Umbelliferae)[J].Plant Systematics and Evolution,1985,151: 89-101.
[11]HEYWOOD V H.The Biology and Chemistry of the Umbelliferae[M].London:Academic Press,1971.
[12]SHNEYER V S,KUTYAVINA N G,PIMENOV M G.Systematic relationshipswithinandbetweenPeucedanumandAngelica(Umbelliferae-Peucedaneae)inferred from immunological studiesof seed protein[J].Plant Systematics and Evolution,2003,236: 175-194.
[13]DOWNIE S R,RAMANATH S,KATZ-DOWNIE D S,et al.MolecularsystematicsofApiaceaesubfamilyApioideae: phylogeneticanalysesofnuclearribosomalDNAinternal transcribed spacer and plastidrpoC1 intron sequences[J].American Journal of Botany,1998,85:563-591.
[14]DOWNIE S R,PLUNKETT G M,WATSON,M F,et al.Tribes and clades within Apiaceae subfamily Apioideae:the contribution of molecular data[J].Edinburgh Journal of Botany,2001,58:301-330.
[15]DOWNIE S R,KATZ-DOWNIE D S.Phylogenetic analysis of chloroplastrps16 intron sequences reveals relationships within the woody southernAfricanApiaceae subfamily Apioideae[J].Canadian Journal of Botany,1999,77:1120-1135.
[16]DOWNIE S R,KATZ-DOWNIE D S.A molecular phylogeny of Apiaceae subfamily Apioideae:evidence from nuclear ribosomal DNA internal transcribed spacer sequences[J].American Journal of Botany,1996,83:234-251.
[17]VALIEJO-ROMAN K M,TERENTIEVA E I,SAMIGULLIN T H,et al.Molecular data(nrITS-sequencing)reveal relationships among Iranian endemic taxa of the Umbelliferae[J].Feddes Repertorium,2006,117:367-388.
[18]SPALIK K,REDURON J P,DOWNIE S R.The phylogenetic position ofPeucedanum sensu latoand allied genera and their placement in tribe Selineae(Apiaceae,subfamily Apioideae)[J].Plant Systematics and Evolution,2004,243:189-210.
[19]舒璞,佘孟兰.中国伞形科花粉图志[M].上海:上海科学技术出版社,2002.
[20]孟德玉,周颂东,何兴金,等.四川前胡属的花粉形态及其系统学意义[J].西北植物学报,2004,24(12):2341-2345.
[21]刘启新,惠红,佘孟兰.前胡属(PeucedanumL.)血清分类学研究[J].植物资源与环境,1998,7(1):20-26.
[22]惠红,刘启新,刘梦华.中国伞形科前胡族阿魏亚族血清分类及亲缘关系的研究[J].植物分类学报,2003,41(4):369-380.
[23]刘启新,惠红.中国前胡属酯酶同工酶与种类演化和地理分布格局[J].植物资源与环境,1999,8(4):17-24.
[24]LIU M,PLUNKETT G M,LOWRY P P,et al.The taxonomic value of fruit wing types in the order Apiales[J].American Journal of Botany,2006,93:1357-1368.
[25]蒲高忠,刘启新.中国伞形科滇芎属及其近缘属果实表面微形态特征的分类学意义[J].植物资源与环境学报,2006,15 (3):1-6.
[26]蒲高忠,刘启新.中国伞形科东俄芹属果实表面微形态特征及分类学意义[J].广西植物,2008,28(5):576-579.
[29]刘长江,林祁,贺建秀.中国植物种子形态研究方法和术语[J].西北植物学报,2004,24(1):178-188.
(责任编辑:佟金凤)
图版说明Explanation of Plates
图版Ⅰ1.华北前胡,示棱槽细胞表面粗糙;2.芷叶前胡,示棱槽细胞表面光滑;3.前胡,示棱槽细胞表面凹陷;4.滨海前胡;5.天竺山前胡; 6.华北前胡;7.少毛北前胡;8.广序北前胡;9.长前胡;10.华山前胡;11.泰山前胡;12.芷叶前胡;13.竹节前胡;14.台湾前胡;15.北京前胡.
PlateⅠ1.Peucedanum harrysmithiiFedde ex Wolff,showing rough surface of vallecular cell;2.P.angelicoidesWolff ex Kretschm.,showing slightly smooth surface of vallecular cell;3.P.praeruptorumDunn,showing concave surface of vallecular cell;4.P.japonicumThunb.;5.P.ampliatumK.T.Fu;6.P.harrysmithiiFedde ex Wolff;7.P.harrysmithiivar.subglabrum(Shan et Sheh)Shan et Sheh;8.P.harrysmithiivar.grande(K.T.Fu)Shan et Sheh;9.P.turgeniifoliumWolff;10.P.ledebourielloidesK.T.Fu;11.P.wawrae(Wolff)Su ex Sheh;12.P.angelicoidesWolff ex Kretschm.;13.P.dielsianumFedde ex Wolff;14.P.formosanumHayata;15.P.caespitosumWolff.
图版Ⅱ1.红前胡;2.细裂前胡;3.毛前胡;4.前胡;5.华中前胡;6.南川前胡;7.南岭前胡;8.草原前胡;9.刺尖前胡;10-11.会泽前胡;12-13.马山前胡;14-15.石防风.
PlateⅡ1.Peucedanum rubricauleShan et Sheh;2.P.macilentumFranch.;3.P.pubescensHand.-Mazz.;4.P.praeruptorumDunn;5.P.medicumDunn;6.P.dissolutum(Diels)Wolff;7.P.longshengenseShan et Sheh;8.P.stepposumHuang;9.P.elegansKomarov;10-11.P.acauleShan et Sheh;12-13.P.mashanenseShan et Sheh;14-15.P.terebinthaceum(Fisch.ex Trevir.)Fisch.ex Turcz.
Micromorphological features of mericarp surface of Peucedanum L.(Apiaceae)in China and its taxonomic significance
LI Mei-zhi,SONG Chun-feng,LIU Qi-xin①(Institute of Botany,Jiangsu Province and the Chinese Academy of Sciences,Nanjing 210014,China),J.Plant Resour.&Environ.2012,21(2):19-29
Micromorphological features of mericarp surface of 22 species and 2 varieties ofPeucedanumL.in China were observed and described with SEM.The results indicate that there are obvious differences in some micromorphological features of vallecular cells,such as,cell smoothness,cell outline visibility,waxy ornamentation type,with or without cell epidermal protuberance and its type of cells,with or without epidermal hairs and its type and ornamentation,with or without epidermal secretions and its type,which shows abundant diversity at interspecies level.According to these mericarp surface features,these species tested can be divided into four types.TypeⅠ:rough and more hairs on vallecular cell surface-invisible cell outline-dense clustered waxy ornamentation,which containsP.japonicumThunb.,P.ampliatumK.T.Fu,P.harrysmithiiFedde ex Wolff and its two varieties,P.turgeniifoliumWolff,P.ledebourielloidesK.T.Fu andP.wawrae(Wolff)Su ex Sheh.TypeⅡ: slightly smooth and no hairs on vallecular cell surface-invisible or concave of cell outline-dense or obvious striated waxy ornamentation,which containsP.angelicoidesWolff ex Kretschm.,P.dielsianumFedde ex Wolff,P.dissolutum(Diels)Wolff,P.rubricauleShan et Sheh,P.macilentumFranch.,P.praeruptorumDunn,P.medicumDunn,P.formosanumHayata,P.longshengenseShan et Sheh,P.acauleShan et Sheh andP.mashanenseShan et Sheh.TypeⅢ:sparse hairs on vallecular cell surfacecell outline concave or un-obvious-few waxy ornamentation,which containsP.caespitosumWolff,P.stepposumHuang,P.pubescensHand.-Mazz.andP.elegansKomarov.TypeⅣ:un-smooth and no hairs on cell surface-subcircular and convex of cell outline-slightly wavy and long striated waxy ornamentation,which only containsP.terebinthaceum(Fisch.ex Trevir.)Fisch.ex Turcz.Combining with the external morphological features and geographical distribution characters,the taxonomic relationships among species in different types are discussed,the specific taxonomic position ofP.acaule,P.mashanenseandP.terebinthaceumisanalyzed.Inaddition,taxonomicsignificanceof micromorphological features of mericarp surface is also defined inPeucedanumL.
PeucedanumL.;Apiaceae;fruit;SEM;micromorphological feature;taxonomy

李美芝,等:中国伞形科前胡属果实表面微形态特征及分类学意义图版ⅠLI Mei-zhi,et al:Micromorphological features of mericarp surface ofPeucedanumL.(Apiaceae)in China and its taxonomic significancePlateⅠ
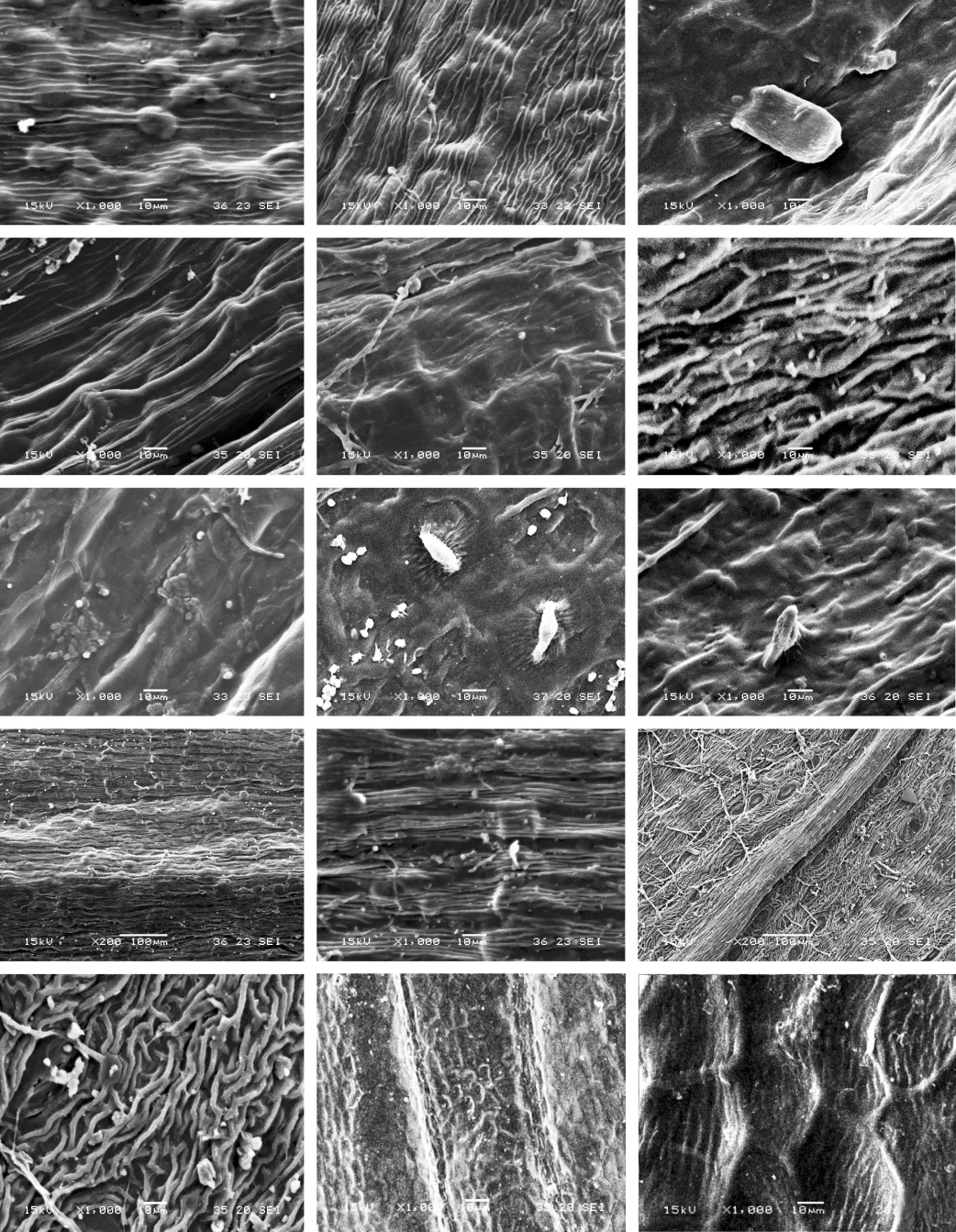
李美芝,等:中国伞形科前胡属果实表面微形态特征及分类学意义图版ⅡLI Mei-zhi,et al:Micromorphological features of mericarp surface ofPeucedanumL.(Apiaceae)in China and its taxonomic significancePlateⅡ
book=2012,ebook=86
Q944.1;Q949.763.3
A
1674-7895(2012)02-0019-11
2012-02-12
国家自然科学基金资助项目(38970077;30370102);中国科学院大科学装置开放研究项目(2009-LSF-GBOWS-01)
李美芝(1985—),女,山东青岛人,硕士研究生,主要从事植物系统分类研究。
①通信作者E-mail:naslqx@yahoo.com.cn
猜你喜欢
食品科学(2022年23期)2022-12-30
石材(2022年2期)2022-05-25
科学大众·教师版(2022年6期)2022-05-23
上海工艺美术(2022年1期)2022-04-21
辽宁农业科学(2021年6期)2021-12-02
北京航空航天大学学报(2021年9期)2021-11-02
紫禁城(2020年7期)2020-07-25
麦类作物学报(2018年8期)2018-08-28
中国兽医杂志(2018年11期)2018-04-16
少年漫画(艺术创想)(2018年12期)2018-04-04
