
不同建植期人工草地优势种植物根系活力、群落特征及其土壤环境的关系
2012-08-20 04:06张莉王长庭刘伟王启兰李里向泽宇
草业学报 2012年5期
张莉,王长庭,刘伟,王启兰,李里,向泽宇
(1.中国科学院西北高原生物研究所,青海 西宁810001;2.中国科学院研究生院,北京100049;3.西南民族大学生命科学与技术学院,四川 成都610041)
草地退化对全球生态安全、环境质量及社会发展的负面影响日益严重,退化草地生态系统功能的恢复与重建已成为当今国际环境科学界关注的热点问题[1,2]。由于全球气候变化、人类长期超载过牧、草地鼠虫危害加剧等因素导致了江河源区草地生态环境日趋恶化,高寒草甸严重退化,以“黑土型”退化草地为主的草地退化现象日趋严重,已成为该区植被恢复重建中亟待重点解决的问题[3]。三江源目前有90%的天然草地出现退化,其中中度退化以上草地面积为571.90×104hm2占三江源总面积的49.83%;可利用草地面积占三江源草地面积的55.40%。“黑土型”退化草地面积为183.63×104hm2,其中核心区有23.48×104hm2,缓冲区有49.02×104hm2,试验区有111.13×104hm2[4]。同20世纪50年代相比,单位面积产草量下降30%~50%,优质牧草比例下降20%~30%,有毒有害类杂草增加70%~80%,草地植被盖度减少15%~25%,优势牧草高度下降了30%~50%,其中黄河源头80~90年代平均草场退化速率比70年代增加了1倍以上[5]。目前,江河源区“黑土型”退化草地植被恢复的主要手段之一是建植人工和半人工草地[6]。人工草地的建设在一定程度上阻止了土壤环境的进一步恶化和草地的放牧压力[7]。在三江源区,常采用老芒麦(Elymus sibiricus)、垂穗披碱草(Elymus nutans)、冷地早熟禾(Poa cryrnophila)、中华羊茅(Festuca sinensis)等多年生牧草建植人工草地,已经取得了较好的经济效益和生态效益[8]。退化生态系统的恢复主要是植被恢复,植被恢复能充分利用土壤-植物复合系统的功能改善局部生态环境,促进生物物种多样性的形成[9]、草地本身的土壤结构及其理化性质恢复,草地植被演替达到稳定阶段[10]。
良好的土壤生态形成是土壤自身质量与整个生态环境质量共同提高的结果,两者相互依存互为促进[11]。由于草地退化核心问题是土壤退化,所以对于退化草地的恢复和治理首先应考虑土壤特征对恢复治理措施的响应过程[12]。根系从土壤中吸取养分,是土壤资源的直接利用者之一;因此,退化草地恢复过程中优势种植物根系活力的重要性和作用是不容忽视的。退化草地的恢复不仅要恢复植被、土壤,更应该考虑根系活力与土壤微环境的相关性,只有在根系活力高、根系发达的情况下,才有源源不断的资源输送到植物地上部分,植物群落生产力、稳定性才有提高和稳定的可能性,土壤环境才会得以改善。但高寒地区人工修复草地优势种植物根系活力及其与土壤环境关系等方面的探讨和研究鲜有报道。因此,本试验以三江源区严重退化高寒草甸(“黑土滩”)、建植期6和10年的人工草地为研究对象,探讨不同建植期人工草地优势种植物根系活力、群落特征及其土壤环境的变化规律和关系,以期为高寒地区退化高寒草甸的恢复和治理提供科学依据。
1 材料与方法
1.1 研究区自然概况
研究区位于青海省果洛州玛沁县 (34°30′~33°34′N,100°29′~99°54′E),平均海拔4 150m,范围为3 800m~4 800m。属典型的高原大陆性气候,无四季之分,仅有冷暖季之别。冷季较长干燥而寒冷,暖季短暂湿润而凉爽。温度年差较小而日差较大,太阳辐射强烈,日照充足,年日照平均值在2 500h以上,年总辐射量在629.9~623.8kJ/cm2。年均气温-1.7℃,1月平均气温-14.8℃,7月平均气温9.8℃,年均降水量600mm,多集中在5-9月,约占年降水量的80%,年蒸发量1 160.3mm,无绝对无霜期。土壤为高山草甸土,高山嵩草草甸为该地区主要草地类型,建群种为小嵩草(Kobresia pygmaea),主要伴生种有矮嵩草(Kobresia humilis)、异针茅(Stipa aliena)等禾草,杂类草有高山紫菀(Aster alpinus)、湿生扁蕾(Gentianopsis paludosa)、高山唐松草(Thalictrum alpinum)、异叶米口袋 (Gueldenstaedtis diversifolia)、黄帚槖吾 (Ligularia virgaurea)等主要植物[13]。
1.2 研究方法
在青海省果洛州玛沁县军牧场分别选择3个不同处理样地,即重度退化的“黑土型”样地(以下简称黑土滩)、6年人工草地和10年人工草地,以“黑土滩”作为对照试验,样地面积均为50m×50m。所选人工草地是以嵩草属(Kobresia)植物为建群种的植被严重退化后经翻耕施肥,条播了垂穗披碱草、冷地早熟禾、中华羊茅和星星草(Puccinellia tenuiflora)的混播人工草地。确定垂穗披碱草为群落的优势种植物。2010年8月中下旬在上述3个试验样地对角线法设置10个50cm×50cm的样方进行植物群落特征(所有植物种的分盖度、高度和频度,以及植物群落的盖度)调查,然后齐地面分种剪草,并按禾本科、莎草科、豆科、杂类草分类后烘干称量。剪去植物地上部分后在50cm×50cm的样方中用内径为5cm土钻采集0~10和10~20cm的土样,分层混合成1个土壤样品过2mm筛,一部分鲜土样保存于4℃冰箱中,用于土壤酶活性的测定,采用比色法测定土壤酶活性[14];另外一部分土样用于测定pH值(电极法)、有机质(丘林法)、全氮和速效氮(凯氏法和康维皿法)、全磷和速效磷(钼锑抗比色法和碳酸氢钠浸提-钼锑抗比色法)含量[15]。采挖各样地群落共有的优势种植物垂穗披碱草带土的根系带回实验室后保存于4℃冰箱中,立即测定根系活力,仔细清洗干净根系后取根尖0~1.0cm处,利用TTC法测定根系活力。土壤容重采用环刀法,温度采用点温计测定,土壤含水量用经典的铝盒烘干法测定。
1.3 数据处理与分析
生物多样性指数[16]包括:

群落多样性指数的计算采用Shannon-Wiener指数:
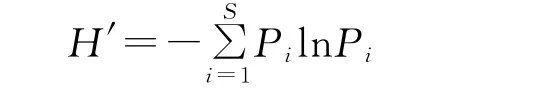
均匀度指数的计算采用Pielou指数:
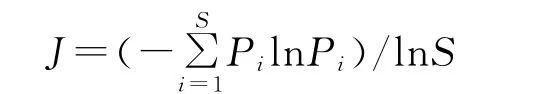
式中,Pi为种i的相对重要值=(相对高度+相对盖度+相对频度)/3,S为种i所在样方的物种总数。
数据分析采用SPSS 16.0进行。
2 结果与分析
2.1 人工草地优势种植物—垂穗披碱草根系活力变化
植物根系是土壤资源的直接利用者和产量的重要贡献者,根系吸收土壤中的水分和矿质养分,并输送到地上部分供植物利用[17]。植物群落结构和土壤环境的变化直接或间接的影响植物根系活力。采挖3处样地对具有代表性的共有种禾本科优势种植物-垂穗披碱草根系进行活力测定,结果显示,不同样地的垂穗披碱草根系活力无差异显著,6年人工草地垂穗披碱草根系活力高于10年人工草地(图1),其变化规律与垂穗披碱草在不同样地的盖度、高度、重要值的变化是相一致的。
2.2 人工草地植物群落结构功能变化
通过对植物群落特征调查分析表明,6年人工草地和10年人工草地植物群落数量特征、种类组成、功能群组成差异性显著(P<0.05)(表1,图2)。以“黑土滩”为对照,人工草地的群落生物量、盖度、高度、物种数、多样性指数提高。“黑土滩”植物群落只有杂类草和禾本科植物组成,杂类草和禾本科植物的生物量分别占群落生物量的90.76%和9.25%;6年人工草地,禾本科植物生物量最大、豆科植物开始入侵,杂类草、禾本科、豆科植物生物量分别占群落生物量的23.37%,76.51%,0.33%;10年人工草地的禾本科植物生物量低于6年人工草地,豆科植物大量入侵,莎草科植物出现,杂类草、禾本科、豆科、莎草科植物生物量分别占群落生物量的59.96%,24.69%,24.69%,12.18%;分析结果表明,随着人工草地建植年限的增加,群落生物量增大、物种数增多、生物多样性提高,植被开始逐渐恢复。
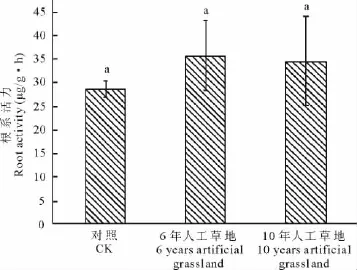
图1 人工草地垂穗披碱草根系活力变化Fig.1 Change of root activity of E.nutans in artificial grassland
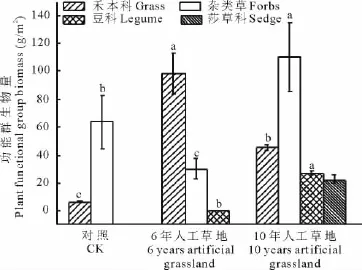
图2 人工草地植物群落功能群生物量变化Fig.2 Change of plant functional group biomass in artificial grassland

表1 人工草地植物群落数量特征Table 1 Quantitative characteristics of plant community in artificial grassland(Mean±SD,n=10)
2.3 人工草地土壤理化特征
土壤是植物赖以生长的基质,其理化特性决定着植物群落类型分布,植物群落又反作用于土壤,逐步改善其生境条件,植被演替随着生境的改变而发生变化。随着人工草地建植年限的延长而发生变化(表2):在0~10cm土层中,土壤含水量、有机质、全氮、速效氮、全磷、速效磷含量显著增大,容重、温度、pH值明显降低;在10~20 cm土层中,温度、pH值、含水量、有机质、全磷、速效磷差异性显著(P<0.05),全氮、速效氮在10年人工草地上明显增加,6年人工草地与黑土滩之间差异不显著。表明人工草地0~10cm土壤理化性质较10~20cm的变化大,土壤微环境得到改善,为土壤种子萌发创造了条件,植物的大量入侵提高了植被覆盖度,防止土壤水分蒸发、土壤含水量增加。土壤温度降低、pH值减小,土壤质量提高,植物赖以生长的土壤环境逐步恢复。
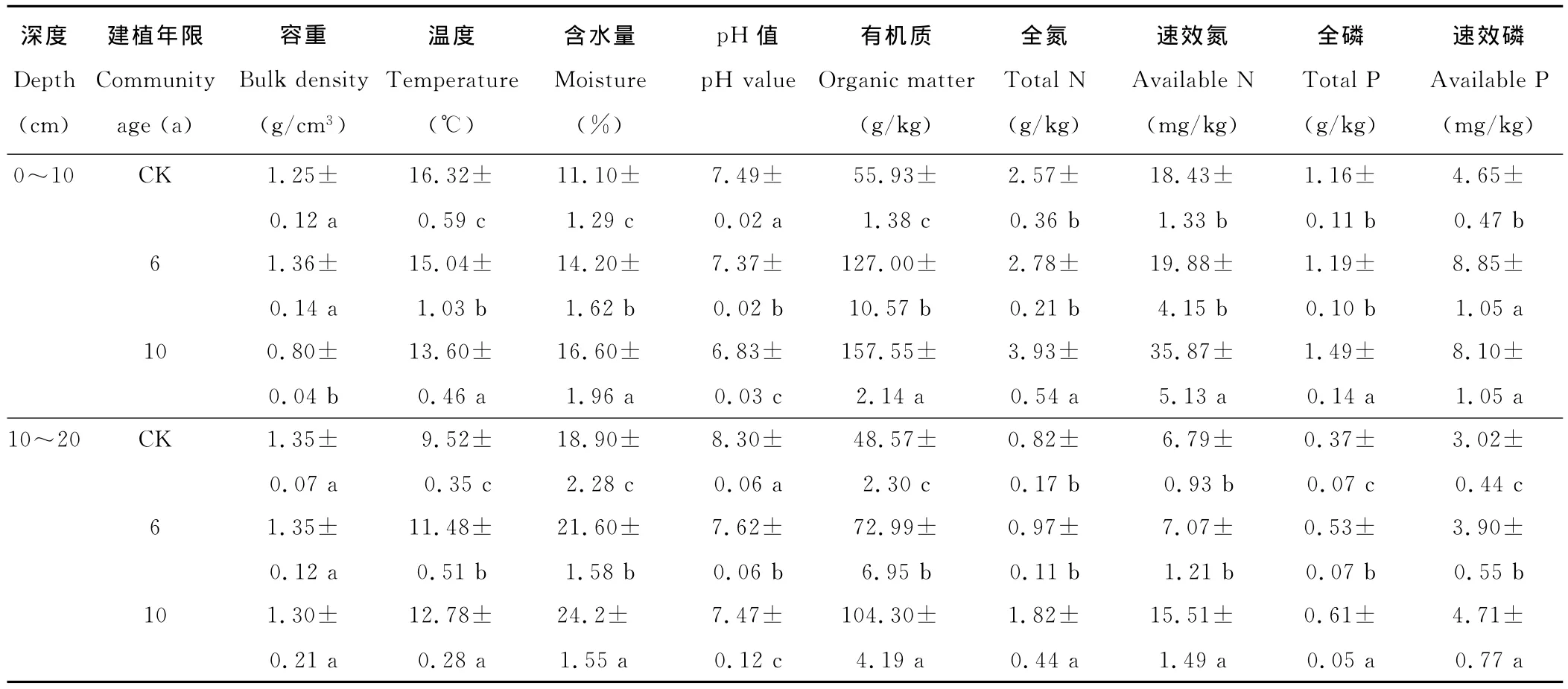
表2 人工草地土壤理化性质Table 2 Soil physicochemical properties of artificial grassland(Mean±SD,n=5)
2.4 人工草地土壤酶活性变化动态
土壤酶是土壤中的植物根系及其残体,土壤动物及其遗骸和微生物分泌的具有生物活性的物质,直接参与土壤物质和能量转化过程[18],土壤酶可作为表征土壤肥力的一个重要指标,研究土壤酶活性的变化,将有助于了解土壤肥力状况及其演变。分析表明,除了过氧化氢酶活性变化不明显外,其他土壤酶活性差异性显著(P<0.05)(图3);在6年人工草地和10年人工草地的蔗糖酶、碱性磷酸酶、蛋白酶、脲酶活性提高,多酚氧化酶活性较黑土滩的均有所下降(图3)。过氧化氢酶在0~10cm土层中无显著差异,10~20cm土层中黑土滩和10年人工草地的均高于6年人工草地,其变化规律不明显(图3)。
2.5 植物群落特征、垂穗披碱草根系活力与土壤微环境关系
通过逐步分析后发现,植物群落特征与垂穗披碱草根系活力、土壤理化性质、土壤酶活性有着不同程度的相关性。群落生物量与土壤有机质、速效氮、速效磷、蛋白酶、蔗糖酶活性正相关(表3~5);群落盖度与土壤含水量、全氮正相关,与pH值负相关;群落高度与土壤含水量、有机质、全磷、脲酶、蛋白酶正相关,与土壤pH值负相关;群落物种数与土壤有机质、全磷、速效磷、速效氮、脲酶、蔗糖酶、蛋白酶正相关。同时,群落生物量、高度、物种数与多酚氧化酶呈负相关。多酚氧化酶只有在土壤贫瘠时才会被激活,可将土壤中的酚类物质氧化为醌,而后形成类腐殖质的大分子化合物[19],所以可将多酚氧化酶活性作为植被恢复过程中土壤生物学的一个指标。土壤中全磷能够促进植物根系扩大,提高植物磷的营养效率[20],脲酶和蛋白酶活性的提高加快了氮素转化和含氮有机物的积累,酶作用底物的增加有力激发了水解酶类的活性[18],因此土壤肥力的逐渐提高,为更多植物入侵提供了养分资源上的保障。
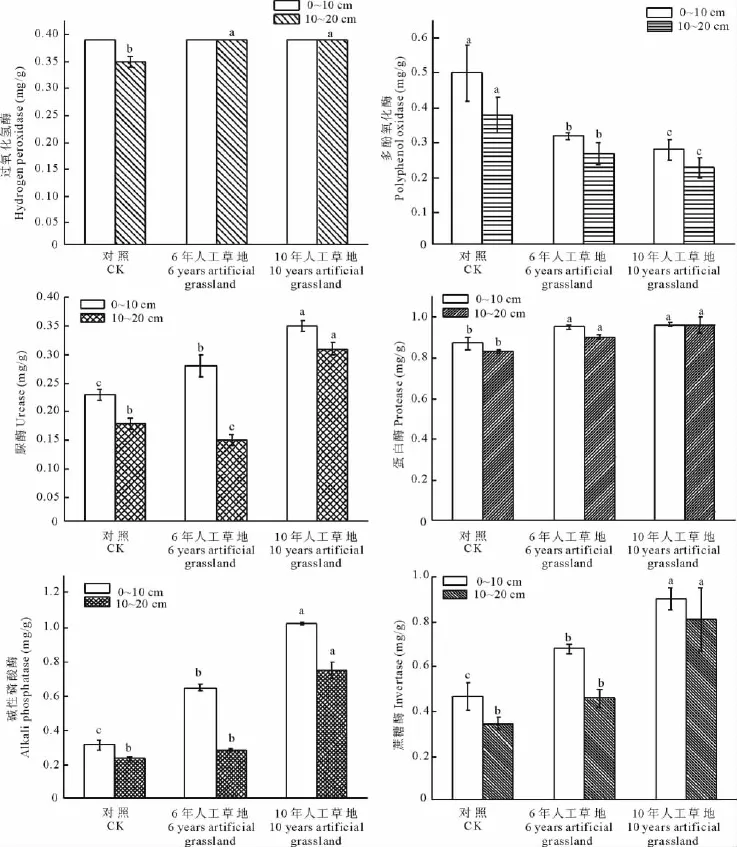
图3 人工草地土壤酶活性变化Fig.3 Change of soil enzymes in artificial grassland
植物根系是植物与土壤环境接触的重要界面,对环境变化较为敏感,更易对环境改变作出快速反应,通过分析优势种植物垂穗披碱草根系活力与群落特征、土壤理化性质、土壤酶活性之间的相关性(表6),发现垂穗披碱草根系活力与土壤含水量、pH值、蛋白酶、碱性磷酸酶和多酚氧化酶活性负相关。说明土壤酸碱度和含水量对垂穗披碱草根系活力有影响,建植人工草地后土壤含水量提高、pH值降低,垂穗披碱草根系活力也随之提高。因土壤中碱性磷酸酶和蛋白酶分别参与土壤中磷和氮的转化,碱性磷酸酶和蛋白酶活性降低影响到植物根系生长和分化所需的氮和磷,垂穗披碱草根系活力因逆境胁迫而降低,黑土滩土壤质量下降使得土壤酶活性降低。6年、10年人工草地由于植物入侵,植被覆盖度逐步提高,植物群落结构和功能发生了明显改变。土壤微环境改变使得植物间对于土壤资源利用竞争增大,优势种垂穗披碱草生物量分配因土壤环境的改变而发生变化,即6年人工草地垂穗披碱草根系活力较高,而10年人工草地群落组成、丰富度增加,垂穗披碱草生物量组成比例下降,其根系活力低于6年人工草地。
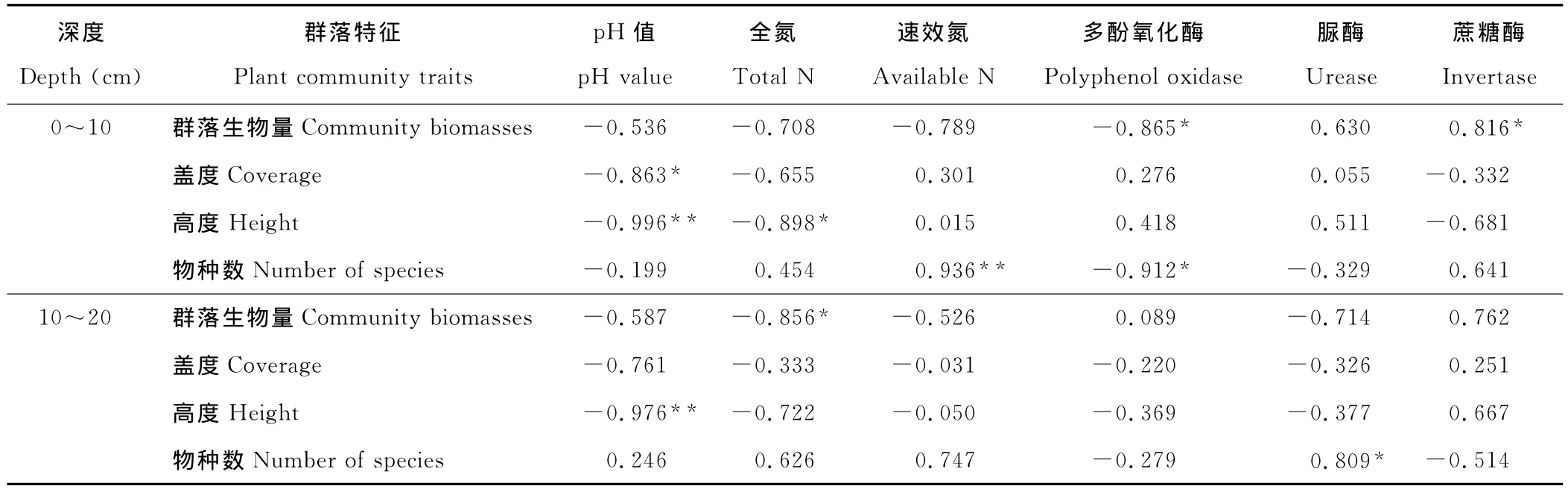
表3 黑土滩(CK)植物群落特征与土壤理化性质、土壤酶活性之间的相关关系Table 3 Correlativity between activities of plant community traits,soil physicochemical properties and soil enzymes in black soil patch(CK)
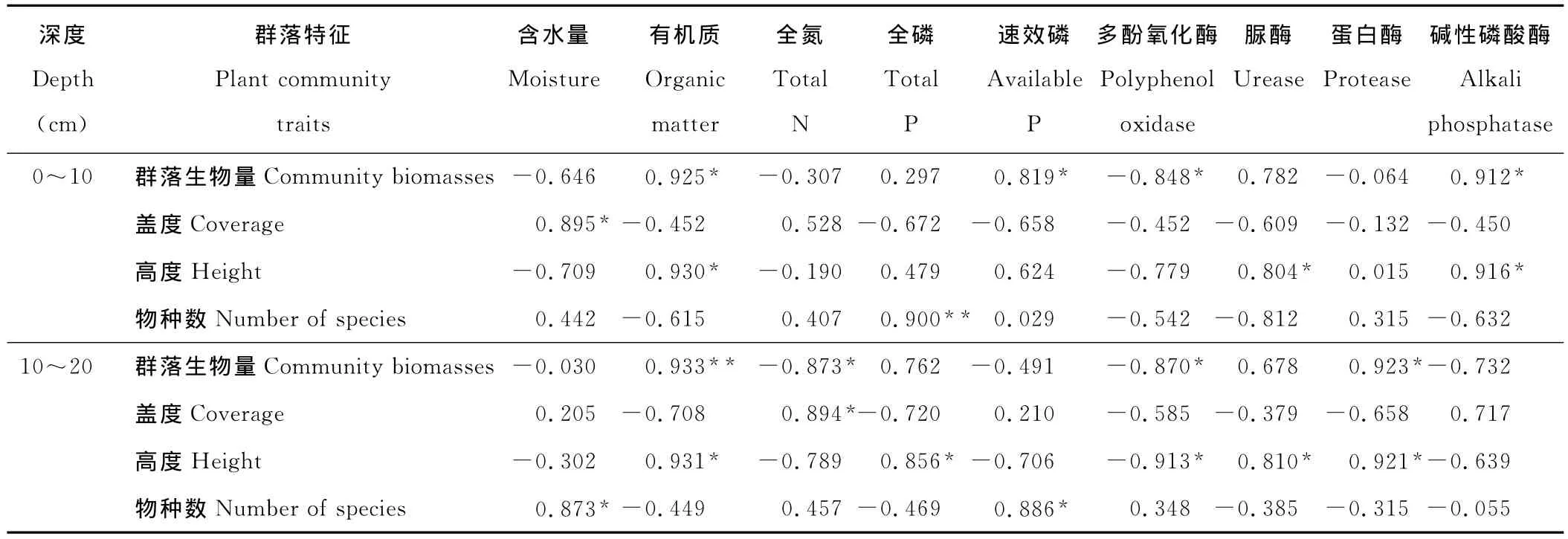
表4 6年人工草地植物群落特征与土壤理化性质、土壤酶活性之间的相关关系Table 4 Correlativity between activities of plant community traits,soil physicochemical properties and soil enzymes in 6years of artificial grassland
3 讨论
土壤是植物赖以生长的基质,土壤质量是土壤供养维持作物生长的能力,包括团聚作用、有机质含量、土壤深度、持水能力、渗透速率、pH值变化、养分能力等[21]。任何现状植被都处于演替系列的某个阶段,植物群落的演替可以改变土壤性质,而土壤性质的改变又可导致群落类型的改变[22]。植物群落的演替是群落对起初阶段异化的过程,不但体现在物种的竞争上,也体现在环境条件的改变上,使生境更适合于演替后来种[23]。而不同植物对土壤条件的适应性以及在不同肥力下的植物种群拓殖能力也不同。土壤状况,尤其是肥力状况影响着群落优势种的拓殖和更替,土壤肥力提高有利于演替后续种的生长和发展,促进群落演替过程。土壤作为影响植物群落生长的主要环境因子,其肥力是土壤基本属性和质的特征,它对群落演替的影响不容忽视[24]。
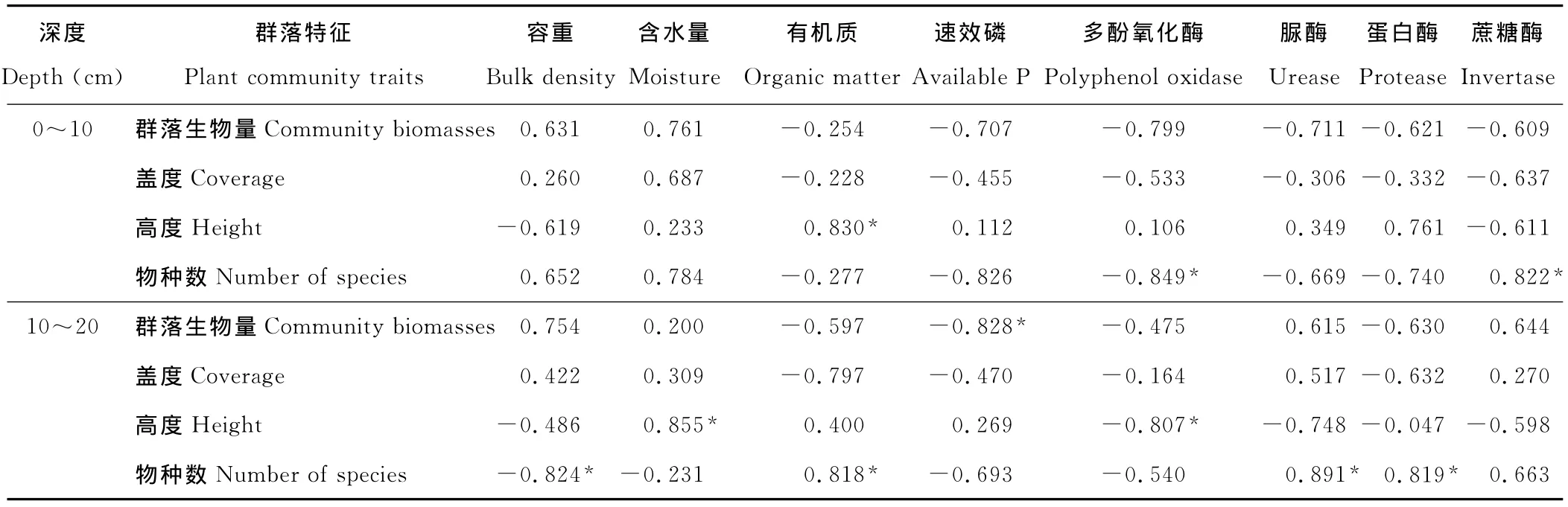
表5 10年人工草地植物群落特征与土壤理化性质、土壤酶活性之间的相关关系Table 5 Correlativity between activities of plant community traits,soil physicochemical properties and soil enzymes in 10years of artificial grassland
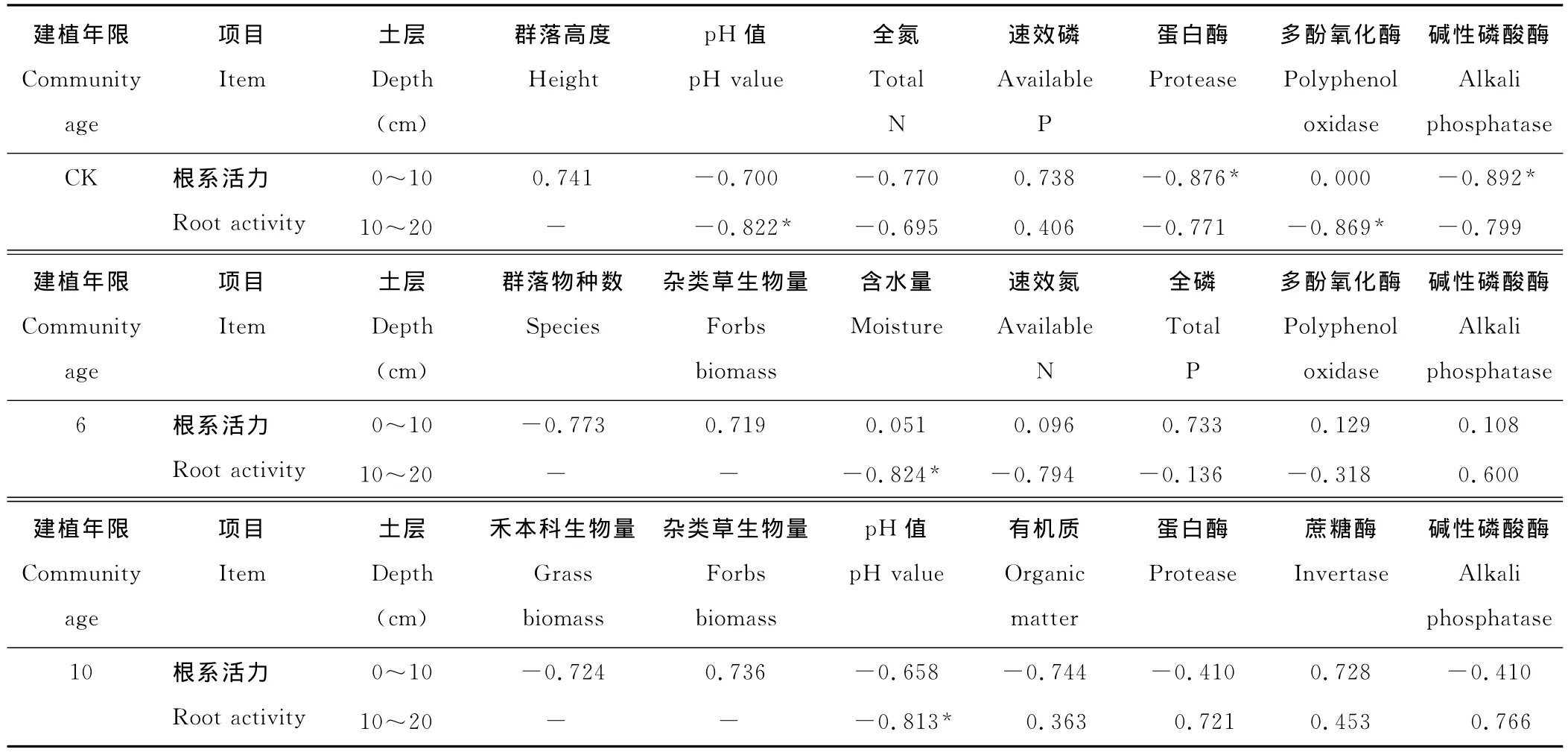
表6 人工草地垂穗披碱草根系活力与群落特征、土壤环境的相关关系Table 6 Correlativity between activities of root activity of Elymus nutans,plant community traits,soil micro-environment in artificial grassland
三江源区高寒草甸由于放牧管理不当,家畜践踏损伤牧草改变了草地植物竞争格局,以及鼠虫害对草地的影响,加速了植物群落内种群更新,引起群落结构与功能的变化[25]。超载过牧导致土壤质量下降[26],草场退化,生物多样性降低[27],生产力下降[28],草地生态环境日趋恶化,“黑土型”退化草地面积增加[29]。在黑土滩上,通过人工措施建立多年生人工草地,短期内可恢复植被,并具有一定生产力[30]。本研究发现随着人工草地建植年限的增加,植物群落结构发生显著变化(表1,图2),土壤质量明显提高(表2),此结论与魏学红等[12],王长庭等[31]的研究结论相同,通过相关性分析表明土壤理化性质对植物群落的影响是显而易见的(表3~5)。“黑土型”草地植物群落组成中主要有杂类草及少量的禾本科植物组成,其中毒杂草比率占到了60%~85%,草地基本上失去了利用价值;黑土滩0~10和10~20cm的土壤容重增大、温度升高、含水量下降、pH值增加、有机质严重减少、全氮和速效氮减少、全磷和速效磷流失严重,土壤质量明显下降;垂穗披碱草根系活力最低(图1),从而反映出黑土滩植物群落生境退化严重,鼠虫害和过度放牧使得植被覆盖度降低,土壤质量下降,植物根系从土壤中能够吸收的有效养分降低,从而影响到地上植物生长,优良牧草几乎消失殆尽。6年人工草地禾本科植物生物量最大(图2),杂类草生物量较黑土滩的降低,豆科植物开始入侵,植被覆盖度增大,土壤理化性质明显发生变化;垂穗披碱草盖度占群落盖度20%~80%,其根系活力最大(图1),表明土壤微环境改变向着适宜垂穗披碱草大量繁殖的方向发展,垂穗披碱草大量繁殖和杂类草生长为豆科植物入侵提供了必要条件。在10年人工草地上,杂类草生物量最大、毒杂草盖度降低,禾本科植物生物量比6年人工草地降低,豆科植物大量入侵,莎草科植物开始入侵,物种多样性提高;土壤微环境变化较明显,土壤质量显著提高,为莎草科的入侵提供了条件,杂类草生物量最大说明了群落演替并未达到高山嵩草草甸顶级阶段;垂穗披碱草根系活力较6年人工草地的有所降低,说明垂穗披碱草较其他禾本科植物更能适应贫瘠的土壤环境。10年人工草地土壤肥力提高为其他物种入侵提供条件,植物间竞争格局发生变化,与垂穗披碱草具有相似土地利用方式的物种入侵,致使垂穗披碱草地上和地下部分竞争力降低,植物群落物种数的增加、种间竞争及物种更替促进了植物群落优势种演替,植物群落多样性增加,群落稳定性提高。
植物群落的演替进程,是植物对土壤不断适应和改造的过程,土壤理化性质是植物演替的主要驱动力之一[32];植物群落与土壤彼此影响相互促进的作用,是植被恢复演替的动力。建植人工草地后,土壤理化性质随着群落演替而发生了变化。土壤演替过程中,土壤酶积极参与土壤发生与发育、土壤肥力的形成、土壤净化等土壤中重要的代谢过程,土壤酶活性反映土壤中生物代谢和物质转化状况,可以表征土壤中正在进行着的各种生物化学过程的方向和程度,可作为土壤活性和土壤生产力指标[33,34]。本研究分析表明土壤蔗糖酶、碱性磷酸酶、蛋白酶、脲酶、多酚氧化酶活性变化动态与土壤养分、群落特征、垂穗披碱草根系活力的变化是相符合的,蛋白酶活性随着人工草地建植年限的增加而提高,说明植物生长使土壤中含氮有机物得以积累,酶作用底物增加有力激发了水解酶类的活性[35]。土壤脲酶和蔗糖酶直接参与土壤含氮有机化合物和碳素循环的转化,碱性磷酸酶促进有机磷化合物的分解,增加了土壤中磷素和易溶性营养物。多酚氧化酶将土壤中多元酚氧化为醌,能促进有机碳累积和土壤腐殖化进程[36]。研究发现土壤多酚氧化酶活性与群落特征、垂穗披碱草根系活力呈负相关,随着人工草地建植年限的增加,多酚氧化酶活性在降低,此研究结论与王启兰等[37]和岳忠辉等[19]的研究结果相同。土壤酶活性变化反映了土壤理化性质的改变对植物群落特征和优势种植物垂穗披碱草根系活力的影响,6年人工草地群落中,土壤有效养分增加和土壤酶活性提高促进垂穗披碱草根系生长,其生物量和根系活力最高;10年人工草地的土壤肥力较高,土壤肥力提高有利于演替后来种的生长和发育,促进群落演替过程,其垂穗披碱草根系活力较6年人工草地的有所降低。随着人工草地建植年限增加,植物群落与土壤微环境彼此相互作用,人工草地群落逐渐稳定,演替进程向顶级方向发展。因此,植物群落的演替过程,是物种对土壤肥力不断适应和改造,更是不同物种在不同肥力梯度下相互竞争和替代的过程。
[1] 许中旗,李文华,鲍维楷,等.植被原生演替研究进展[J].生态学报,2005,25(12):3383-3389.
[2] 刘兴元,龙瑞军,尚占环.草地生态系统服务功能及其价值评估方法研究[J].草业学报,2011,20(1):167-174.
[3] 黎与.江河源区高寒草甸退化草地恢复与重建技术研究[J].草业与畜牧,2007,4:30-32.
[4] 三江源自然保护区生态保护与建设编辑委员会.三江源自然保护区生态保护与建设[M].西宁:青海人民出版社,2007:77-78.
[5] 刘纪远,徐新良,邵全琴.近30年来青海三江源地区草地退化的时空特征[J].地理学报,2008,63(4):364-376.
[6] 武高林,任国华,刘振恒,等.黄河首曲湿地功能区“黑土型”退化草甸恢复改良[J].兰州大学学报(自然科学版),2009,45(4):48-52.
[7] 马玉寿,尚占环,施建军,等.黄河源区”黑土型”退化草地人工群落组分配置技术研究[J].西北农业学报,2007,16(5):1-6.
[8] 马玉寿,郎百宁,李青云,等.江河源区高寒草甸退化草地恢复与重建技术研究[J].草业科学,2002,19(9):1-5.
[9] 王炜,刘钟龄,郝敦元,等.内蒙古草原退化群落恢复演替的研究Ⅰ.退化草原的基本特征与恢复演替动力[J].植物生态学报,1996,20(5):449-459.
[10] 欧阳克蕙,王堃.人工草地植被重建对退化红壤化学性质和酶活性的影响[J].江西农业大学学报,2007,29(3):474-478.
[11] Fu B J,Chen L D,Ma K M.The effect of land use change on the regional environmental in the Yangjuangou Catchment in the Loess Plateau of China[J].Acta Geographica Sinica,1999,54(3):241-246.
[12] 魏学红,孙磊,武高林.青藏高原东缘“黑土型”退化草甸人工草地改良的土壤养分响应[J].水土保持学报,2010,24(5):153-168.
[13] 王长庭,龙瑞军,王启兰,等.三江源区高寒草甸不同退化演替阶段土壤有机碳和微生物量碳的变化[J].应用与环境生物学报,2008,14(2):225-230.
[14] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
[15] 中国科学院南京土壤研究所.土壤理化分析[M].上海:上海科学技术出版社,1983:62-107.
[16] 马克平,刘玉明.生物群落多样性的测度方法.Iα多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[17] 周本智,张守攻,傅懋毅.植物根系研究新技术Minirhizotron的起源、发展和应用[J].生态学杂志,2007,26(2):253-260.
[18] Aon M A,Colaneri A C II.Temporal and spatial evolution of enzymatic activities and physical-chemical properties in an agricultural soil[J].Applied Soil Ecology,2001,18:255-270.
[19] 岳中辉,王博文,王洪峰,等.松嫩平原西部盐碱草地土壤多酚氧化酶活性及其与主要肥力因子的关系[J].草业学报,2009,18(4):251-255.
[20] 王庆仁,李继云,李振声.植物高效利用土壤难容态磷研究动态及展望[J].植物营养与肥料学报,1998,4(2):107-116.
[21] Larson W E,Pierce F J.Conservation and enhancement of soil quality[A].In Proc.of the Int.Workshop on Evaluation for Sustainable Land Management in the Developing World[C].Bangkok,Thailand:Int.Board for Soil Res.and Management,1991:12.
[22] Cooperrider A.Conservation of biodiversity on western rangelands[A].Van Wieren S E.Grazing and Conservation Management Kluwer,Dordrecht,the Netherlands[C].South Texas:Development and Impl-ementation,1991.
[23] 张庆费,宋水昌,由文辉.浙江天童植物群落次生演替与土壤肥力的关系[J].生态学报,1999,19(2):174-178.
[24] 邹厚远,刘国彬,王晗生.子午岭林区北部近50年植被的变化发展[J].西北植物学报,2002,22(1):l-8.
[25] 侯扶江,常生华,于应文,等.放牧家畜的践踏作用研究评述[J].生态学报,2004,24(4):784-789.
[26] 黄德青,于兰,张耀生,等.祁连山北坡天然草地地上生物量及其与土壤水分关系的比较研究[J].草业学报,2010,20(3):20-27.
[27] Crist P J,Thomas W K,John O.Assessing land use impacts on bio-diversity using an expert system tool[J].Landscape Ecology,2000,15:47-62.
[28] 侯琼,王英舜,师桂花,等.内蒙古典型草原作物系数的试验研究[J].草业学报,2010,20(4):34-41.
[29] 马玉寿,郎百宁,李青云,等.江河源区高寒草甸退化草地恢复与重建技术研究[J].草业科学,2002,19(9):1-4.
[30] 文金花,马玉寿,施建军,等.利用草地早熟禾改建江河源区“黑土型”退化草地的研究[J].草原与草坪,2006,(2):41-45.
[31] 王长庭,曹广民,王启兰,等.三江源地区不同建植期人工草地植被特征及其与土壤特征的关系[J].应用生态学报,2007,18(11):2426-2431.
[32] 张庆费,曲文辉,宋永昌.浙江天童植物群落演替对土壤化学性质的影响[J].应用生态学报,1999,10(1):19-22.
[33] Acosta-Martinez V,Tabatabai M A.Enzyme activities in a limed agricultural soil[J].Biology and Fertility of Soils,2000,31:85-91.
[34] Frankenberger W T,Dick W A.Relationships between enzyme activities and microbial growth and activity indices in soil[J].Soil Science Society of America Journal,1983,47:945-961.
[35] Acosta-martinez V,Cruz L,Sotomayor-ramirez D,et al.Enzyme activities as affected by soil properties and land use in a tropical watershed[J].Applied Soil Ecology,2007,35:35-45.
[36] An S S,Huang Y M,Zheng F L.Evaluation of soil microbial indices along a revegetation chronosequence in grassland soils on the Loess Plateau,Northwest China[J].Applied Soil Ecology,2009,41:286-292.
[37] 王启兰,王长庭,刘伟,等.江河源区人工草地植物群落和土壤酶活性变化[J].应用与环境生物学报,2010,16(5):662-666.
猜你喜欢
美文(2022年18期)2022-10-20
草地学报(2022年8期)2022-08-31
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
现代畜牧科技(2021年7期)2021-07-28
西藏农业科技(2021年4期)2021-04-18
河南科学(2020年3期)2020-06-02
种子(2019年8期)2019-09-11
生态学报(2019年4期)2019-04-08
草业学报(2018年7期)2018-07-30
