
渤海湾西南部典型站位营养盐限制特性的加富培养实验研究
2012-08-14 05:48:52穆迪李清雪陶建华赵海萍聂红涛
海洋通报 2012年6期
穆迪,李清雪,陶建华,赵海萍,聂红涛
(1.河北工业大学 海水资源高效利用化工技术教育部工程研究中心,天津 300130;2.河北工程大学 分析测试中心,河北 邯郸 056038;3.天津大学 环境科学与工程学院,天津 300072)
近年来,随着一些海域富营养化和赤潮问题的加剧,以及利用海洋生物泵吸收CO2缓解温室效应问题的升温,营养盐对于浮游植物的限制作用再次成为海洋学研究的热点(刘慧等,2002)。加富培养实验直观、可操作性强,接近于自然状况,是研究营养盐限制特性的常用方法之一(蒲新明等,2000)。在我国,这项研究始于20世纪末,研究海域主要集中在黄渤海、东海近海海域(邹立等,2001;王勇等,1999;刘媛等,2004)、台湾海峡(王玉珏等,2008;胡俊等,2008)及胶州湾(王勇等,2002)、大亚湾(王晓伟等,2007)等海域,结果表明,氮和磷是这些海域主要的限制性营养盐,但具体的限制类型又体现了较大的时空差异性。目前这类实验的目的主要还是营养盐限制类型的判定,对培养条件生态影响的研究则较少。氮磷比是影响浮游植物生长的重要因素之一(Hodgkiss et al,1997),各种浮游植物对氮磷营养盐的需求不同,一个海域的最适宜氮磷比通常具有特异性,Redfield比只是一个平均水平(Hecky et al,1988),但以往的多因素培养实验多是基于Redfield比进行设计,没有对所研究海域的最适宜氮磷比进行定量分析并加以考虑。唐松(2010)在印度洋进行氮、磷、铁加富的现场培养实验时,分析了实验过程中氮磷比与浮游植物生长之间的关系,认为二者相关性不大,氮磷比不能单独决定浮游植物的生长,但该实验是在相对独立、加富条件差异较大的培养组间进行的,此时营养盐的绝对浓度影响更大,对于氮磷比的作用还需在相对一致的条件下进行系统研究。此外,以往的加富培养实验采用的多是营养盐的一次性添加,这种添加方式更接近冲击性高负荷排污的情况,并不适合常见的排污情况,对于不同添加方式的影响应予以考虑。虽然王玉珏等(2008)在进行台湾海峡的研究时使用了连续添加,但是没有对同等药量的一次性添加进行对比,从而分析添加方式的影响。
渤海湾位于渤海西部,属典型的缓坡淤泥质半封闭浅海海湾。渤海湾沿岸有海河、永定新河等几十条河流注入,每年有大量污水入海。该海域水交换能力弱,污染物滞留时间长,致使该海域污染严重、赤潮频发(Tao,2002)。目前,对于渤海湾内营养盐限制及其相关特性的调查研究多是基于水体中溶解无机营养盐的现存量进行的对比分析(Zou et al,1985;沈志良,1999;阚文静等,2010)。这些研究表明,近年来渤海湾近岸海域水体营养盐的比例和含量发生了较大变化,进一步研究营养盐比例和含量的变化对浮游植物生长的影响是十分必要的。
本次研究的主要工作是通过改进实验设计,对渤海湾西南部海域典型站位2010年10月的营养盐限制特性进行加富培养的实验研究。研究的主要内容包括营养盐限制类型的判定,浮游植物对于不同氮磷比和不同硝酸盐添加方式的生态响应。本次研究不仅是一次实验调查,也是一次实验改进的尝试。
1 材料与方法
1.1 采样方法
采样站位位于渤海湾西南部(117°49′56″E,38°38′4″N),为渤海湾生态监控区常规监测站位之一,采样时间为2010年10月。采样时采集表层水,采样后用200 μm的筛网轻柔过滤,以滤除大型浮游动物,避免摄食干扰。
1.2 实验方法
水样采集后,在常温状态下,用汽车迅速运回实验室,进行混合、分装和培养,水样的采集和运输共耗时6 h左右。每桶初始水量15 L,进行营养盐添加后将桶置于光照培养架上进行培养。用空调对培养室室温进行了调节,日间控制室温在18℃~20℃左右,夜间利用常温培养,室温最低在11℃~13℃左右。光照强度控制在初始水位处光强6 000 lux,明暗周期为12-12 h。光照是影响浮游植物光合作用的重要因素,在海上进行现场培养实验时通常采用自然光照(邹立等,2001;王勇等,1999;王玉珏等,2008)或维持培养环境与现场接近(刘媛等,2004;胡俊等,2008;王勇等,2002),在室内进行模拟现场培养实验时为模拟现场环境或控制适宜培养条件通常需要控制光照(韦桂秋,2006;邓华健等,2004),本文光照强度的设置主要考虑将其控制在浮游植物生长的适宜范围。一般而言,在一定范围内,浮游植物的光合作用随着光照的增强而增强,但达到一定强度(饱和光强)后,光强的增加不再具有促进作用,反而会对光合作用产生抑制,浮游植物最适宜的光强范围一般在 2 000~10 000 lux(邓华健 等,2004;孙霞,2005)。本文参照此规律及相关研究结果并考虑了水深的影响,选用了6 000 lux。12-12 h的明暗周期是比较常见的模拟实际光照的方法。
每天固定时间混合水样3次,防止浮游植物聚集。培养周期为14 d。本文浮游植物培养周期的设置主要参考了相关实验研究。文献表明,加富培养实验的培养周期通常在3~12 d不等(王晓伟等,2007;胡俊等,2008;邓华健等,2004),或动态的指定到某一生长阶段(如指数生长期/最大增值期第4 d)(韦桂秋,2006),本文考虑到实验初期的生长延迟现象,最终确定培养周期为14 d。浮游植物培养周期的选取主要与浮游植物的生长周期有关,应尽量使二者相吻合,能在培养期间清楚地观察到整周期的生长变化。
营养盐添加时,硝酸盐、磷酸盐和硅酸盐分别以KNO3、KH2PO4、Na2SiO3形式进行添加。采样和实验所用器具使用前均使用HCl渍洗和海水润洗。
1.2.1 实验一:营养盐限制类型的加富培养实验
当浮游植物受到某种营养盐限制时,人为添加这种营养盐,会促进浮游植物的生长。把一种和多种营养盐加入到一定水体中,通过观察比较浮游植物的生长情况,判断其中的营养盐限制情况,是用加富培养实验判定营养盐限制类型的原理和设计依据。这种判定方法也称AGP(Algal Growth Potential,藻类增长潜力)法(韦桂秋,2006)。
无机氮(硝酸盐、亚硝酸盐、氨氮)、磷酸盐和硅酸盐是对海洋浮游植物生长最重要的几种营养盐。陆源排放是渤海湾营养盐的主要来源,近年来渤海湾营养盐陆源污染严重、比例失调,近岸海域水体营养盐的比例和含量发生了较大变化(阚文静等,2010),实验一即为验证水体营养盐的比例和含量发生变化的情况下,浮游植物的营养盐限制类型是否也发生变化。
本实验中,为判定原水样的营养盐限制类型,对水样进行了硝酸盐、磷酸盐、硅酸盐的单一和全部添加(表1)。在浮游植物适宜范围内,在系统本底浓度的基础上按倍数增大营养盐的浓度是加富培养实验的一种常见的营养盐添加方式,文献中的增大倍数在2倍(邹立等,2001)、10倍(刘媛等,2004;王晓伟等,2007)和100倍(王晓伟等,2007)不等。本文营养盐的添加量也是依据营养盐的本底浓度确定的。其中氮盐增大为本地浓度的30倍左右,磷酸盐增大为本底浓度的20倍左右,硅酸盐增大为本底浓度的15倍左右,进行取整,并考虑了同时添加时各营养盐浓度的比值接近Redfield比。
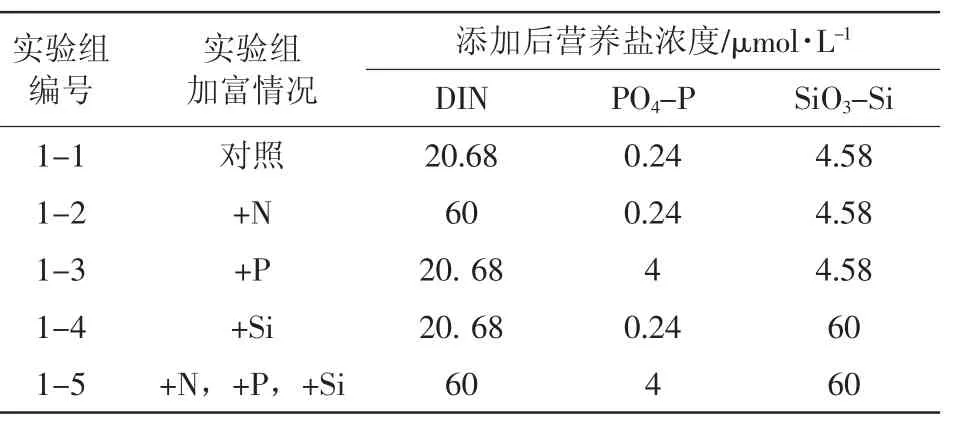
表1 实验一加富方案
1.2.2 实验二:氮磷比生态响应的加富培养实验为考察不同氮磷比对于浮游植物生长的影响,在保证营养盐绝对值充足的情况下(Fisher et al,1999;Nelson et al,1995),添加和控制硝酸盐浓度为一定水平,变化磷酸盐浓度,设计了8个实验组(表2),代表8种不同的氮磷比水平。各实验组浓度梯度的选取主要考虑了普遍条件下适宜浮游植物生长的氮磷比范畴和渤海湾常见的氮磷比范畴(蒲新明 等,2000;Hodgkiss et al,1997;Zou et al,1985;沈志良,1999;阚文静等,2010),并考虑了添加和测定的精度要求。
1.2.3 实验三:硝酸盐添加方式生态响应的加富培养实验
设计了4组对照实验,考察硝酸盐一次性添加和硝酸盐连续性添加情况下,浮游植物生态响应的差异(表3)。其中,硝酸盐一次性添加是模拟冲击性高负荷排污情况,连续性添加是模拟持续性低负荷排污情况。这里,营养盐的连续性添加是指,将与一次性添加等量的营养盐均匀地分7天连续添加,每日药量的计算中考虑了每日取样量对体积的影响。这里控制磷酸盐浓度一定,考察硝酸盐添加的影响主要是基于近年来渤海湾氮污染严重的特点(阚文静等,2010)。
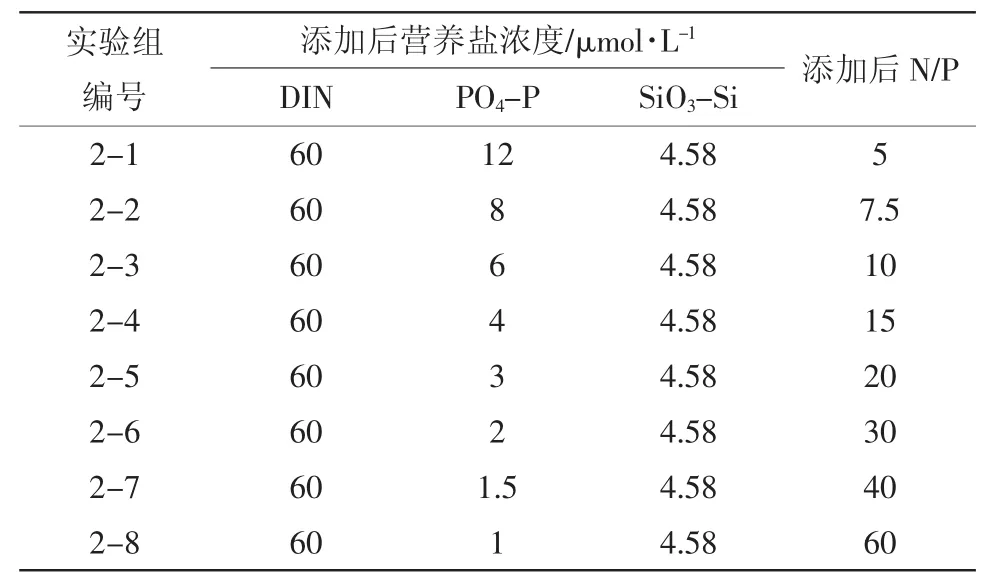
表2 实验二加富方案
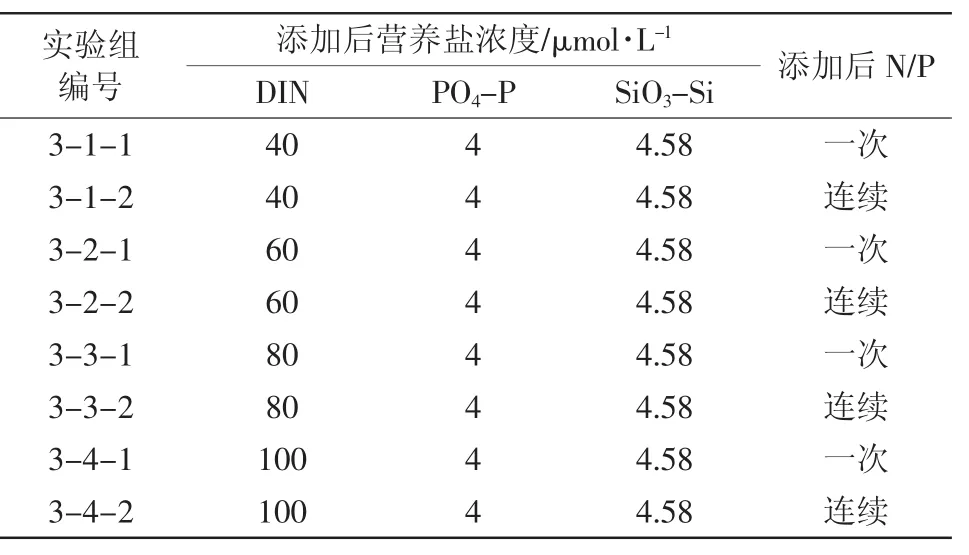
表3 实验三加富方案
1.3 分析方法
2 结果与讨论
2.1 原水样水质
原水样中溶解无机氮浓度为20.68 μmol/L,其中:硝酸盐浓度为16.58 1 μmol/L,亚硝酸盐浓度为 2.356 μmol/L,氨氮浓度为 1.746 μmol/L。水样中磷酸盐浓度0.24 μmol/L,硅酸盐浓度4.58 μmol/L(表1)。原水样中叶绿素a浓度为1.05 μg/L,浮游植物细胞密度为1 080 cells/L,主要有甲藻和硅藻,数量占优势的藻种为叉状角藻和园筛藻。在水样运输过程中,可能出现少量浮游植物死亡,释放营养盐的情况。
2.2 营养盐限制类型
培养期间,单一添加营养盐的各组中,添加磷酸盐的1-3组叶绿素a浓度和浮游植物细胞密度的增长状况最显著,在培养的前8天时间里,1-3组叶绿素a浓度和浮游植物细胞密度的变化趋势最接近于营养盐全部添加的1-5组。其中,1-3组叶绿素a浓度峰值为空白对照组1-1组的2.48倍,达到营养盐全加组1-5组同期浓度的48%,其细胞密度峰值为1-1组的1.66倍,达到1-5组同期密度的72%,这里只给出了细胞密度的变化趋势(图1(a))。同时,各组磷酸盐都出现了明显的消耗,在没有磷酸盐添加的情况下都是最早被耗尽(图1(b))。硅酸盐的消耗也较显著,在没有硅酸盐添加的各组中,最终都趋于耗尽,耗尽时间仅晚于磷酸盐(图1(c))。培养期间,各组溶解无机氮浓度波动较大,总体变化不明显。从浮游植物生长和营养盐消耗的特征来看,该水样为磷限制。
同时,对原水样营养盐浓度进行对比分析表明,若以Fisher(1999)或Nelson(1995)的阈值标准评定氮、磷、硅营养盐都不构成限制,若以Justic(1995)和Dortch(1992)的比例关系评定,则该水样接近于磷限制(Si∶P=19.08,DIN∶P=86.17),这与上述培养实验结论基本一致。
综合以往我国不同海域的实验成果表明,氮和磷是这些海域主要的限制性营养盐,营养盐限制特性具有明显的时空差异性(表4)。同时,由于近岸地化过程和人类活动的影响,近岸海域营养盐限制特性通常呈现一定的复杂性(张平等,2001)。磷限制通常发生在沿岸和较封闭海域(刘慧等,2002),我国曾发现磷限制的海域包括渤海莱州湾附近(邹立等,2001)、大亚湾的湾中和湾顶(王晓伟等,2007)和东海长江口附近站位(刘媛等,2004)。近年来,由于排污,一些近岸海域出现了营养盐浓度增高比例失调,营养盐限制特性发生转变的情况。如对胶州湾进行的培养实验(王勇等,2002)和营养盐浓度分析(张均顺等,1997)表明,胶州湾水体正在趋向于硅限制。渤海湾,属典型的缓坡淤泥质半封闭浅海海湾,水交换能力弱,自净能力差。近年来随着沿岸经济的快速发展,渤海湾营养盐的含量和比例发生了显著的变化,表层海水氮磷比和硅磷比增加,硅氮比较稳定,且小于1,近年来渤海湾浮游植物可能正在从氮限制逐渐转变为磷限制或硅限制(阚文静等,2010)。本文的实验结果与此前报道基本一致。
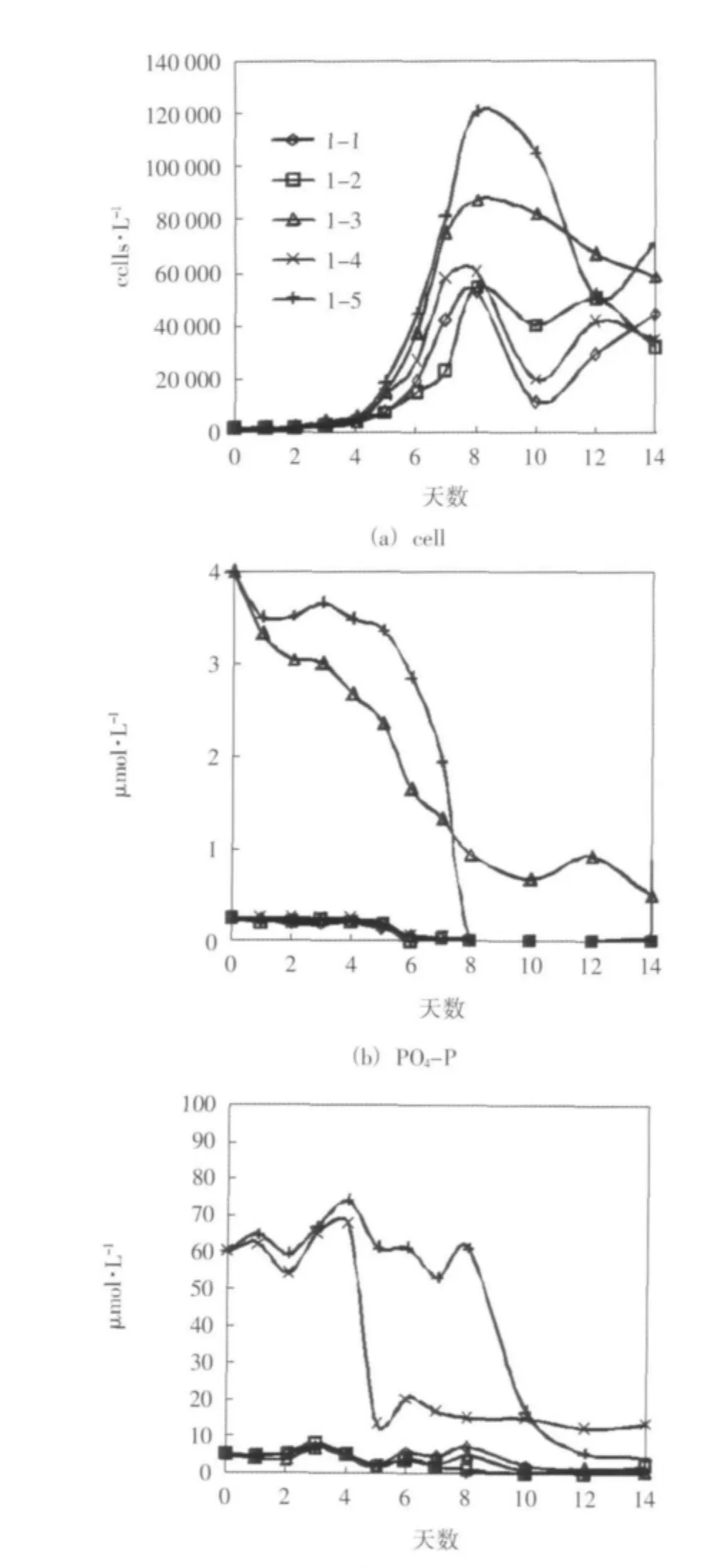
图1 实验一 培养期间细胞密度和营养盐浓度的变化趋势
2.3 不同氮磷比的生态响应
对各组在培养期间叶绿素a浓度、浮游植物细胞密度的变化趋势进行了对比,并对前八天时间里不同氮磷比对应的叶绿素a浓度最大值和细胞密度最大值进行了统计。差异较显著的前五组的细胞密度随时间的变化的情况如图2(a)所示(后3组的细胞密度总体水平较低,差异不显著,这里没有给出)。各组细胞密度最大值与对应氮磷比的关系如图2(b)所示。
结果表明,叶绿素a浓度和浮游植物细胞密度的增长总体上随磷酸盐浓度的升高和氮磷比的降低而增大,并在5-15附近增长最显著,这个范围可以判定为该实验条件下浮游植物生长的最适宜氮磷比,这个氮磷比的范围与香港海域主要赤潮藻种的最适宜氮磷比4-16的范围接近(Ho et al,1993)。相比于Redfield比,这个最佳氮磷比范围偏低,这可能与样品中甲藻较多有关,甲藻对磷的需求量更大,适宜低氮磷比的环境(萧朴云等,2009)。另外,本次实验是在不添加硅酸盐、氮硅比较高(DIN∶DISi=13.1)的情况下进行的,可能存在硅的相对不足而限制硅藻的生长。根据本实验可以推测,如果渤海湾磷酸盐浓度增大,氮磷比降低,渤海湾的富营养化可能造成更为严重的后果,这从另一方面体现了磷的限制作用。
相对于以往的研究(唐松等,2010),本次实验体现了较好的规律性,这可能与本实验中控制无机氮、硅酸盐初始浓度相同,系统条件相对一致有关,同时实验中营养盐浓度偏离正常值较多,这可能促进了少数耐污种类的生长,降低了浮游植物的多样性,从而使响应更单一,这虽然与现实情况有所偏差,但是可以更清晰地反映浮游植物的响应规律。营养盐变化对浮游植物种类的影响还有待进一步研究。
2.4 不同硝酸盐添加方式的生态响应
对各对照组在培养期间叶绿素a浓度、浮游植物细胞密度和营养盐的浓度变化进行了两两对比。生长较显著的3-2组(3-2-1、3-2-2)的情况如图3所示。其他3个对照组的变化规律基本相似,但没有3-2组浮游植物的生长显著、差异明显,这里没有给出。对照组间的这种差异可能与营养盐条件不同有关,3-2组的磷酸盐含量和氮磷比均适宜浮游植物的生长;3-1组虽然氮磷比适宜,但磷酸盐含量偏低;3-3与3-4组无机氮源充足但氮磷比偏高,故3-2组浮游植物的生长更显著。

表4 国内近岸海域加富培养实验研究及其成果
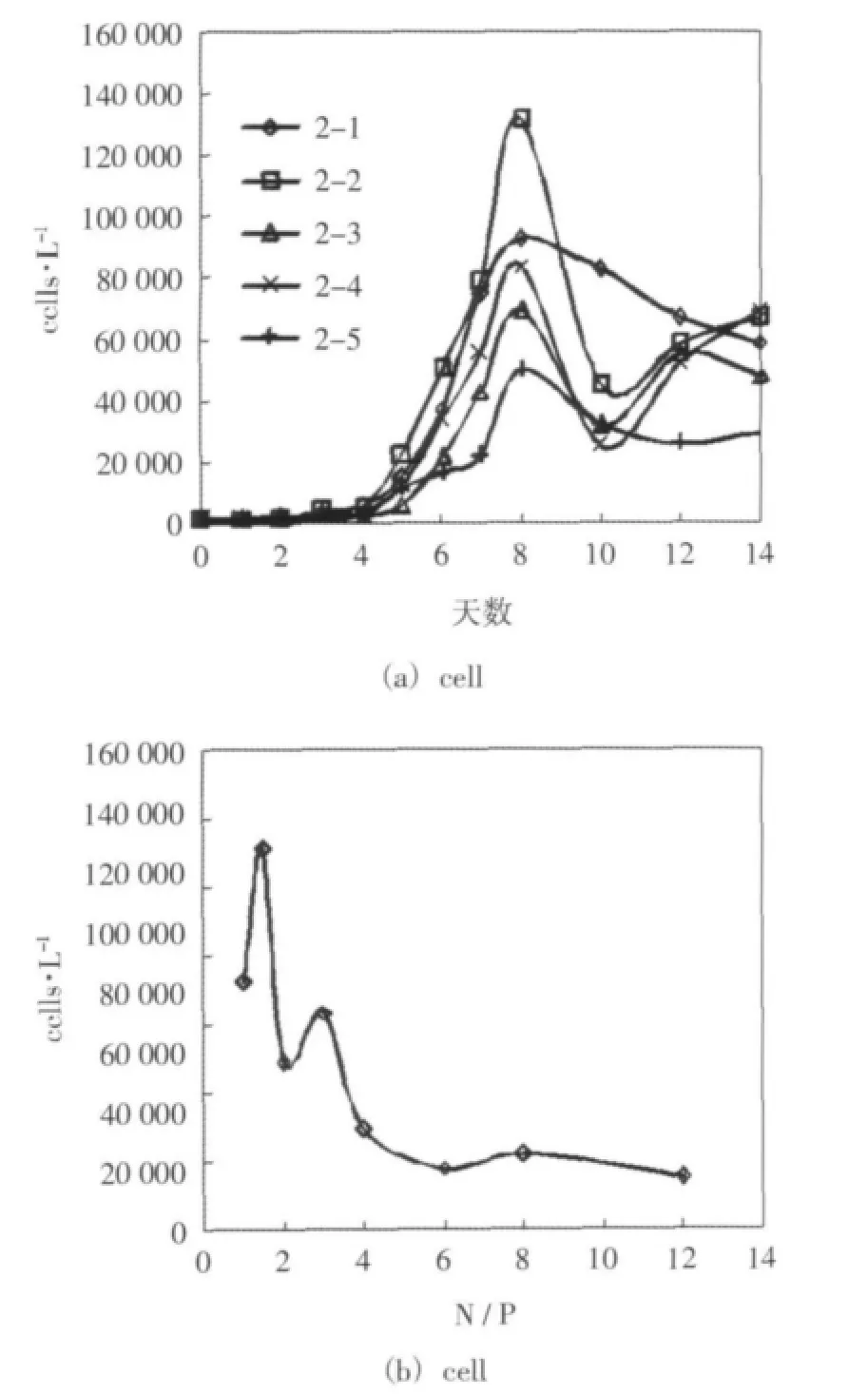
图2 实验二 培养期间细胞密度的变化趋势和细胞密度最大值与氮磷比的关系
结果表明,在其他条件相同的情况下,硝酸盐分七天连续添加时相对于一次性添加时能达到更高的叶绿素a浓度和浮游植物细胞密度。在浮游植物生长旺盛时期,连续性添加组的磷酸盐较一次性添加组的磷酸盐吸收快、浓度低。硝酸盐浓度变化的波动较大,但前期连续性添加组浓度逐渐攀升的趋势明显。在培养前期,由于营养盐添加方式的不同,连续性添加组的氮磷比低于同期一次性添加组的氮磷比,按上文给出的规律,这可能促进了连续性添加组磷的吸收和低氮磷比浮游植物的生长,但这种优势随着硝酸盐的连续添加会逐渐缩小。同时,在培养的后期连续性添加组维持了较高的硝酸盐浓度水平,这对于后期浮游植物的生长可能也是有利的。
该实验结果表明低浓度长期持续性氮污染可能会比高浓度冲击性氮污染更有效地刺激浮游植物的增长,从而造成更严重的生态影响。同时该结果也表明在本实验条件下,用以往的一次性添加培养实验可能会低估浮游植物的增长潜力。
2.5 与渤海湾海域实际状况的比较
为分析自然状况下该海域营养盐变化时浮游植物的变化情况,对20世纪50年代末至21世纪初渤海湾水体营养盐和浮游植物的几次大规模调查研究进行了搜集整理(表5和表6)。综合结果表明,半个世纪以来渤海湾氮磷营养盐含量整体上呈现增长的趋势,尤其是近年来溶解无机氮增长很快,而水体磷酸盐浓度得到一定控制,结果造成水体氮磷比逐年增大,明显偏离了Rieldfied比,水体基本处于磷限制。同期,渤海湾浮游植物的生态特性也发生了较大变化,浮游植物细胞丰度增大,多样性降低,组成更单一,且时空分布极不均匀。水体营养盐浓度和比例结构的变化可能是造成浮游植物群落结构变化的一个直接原因,而根本的原因还是受到陆源污染的影响。
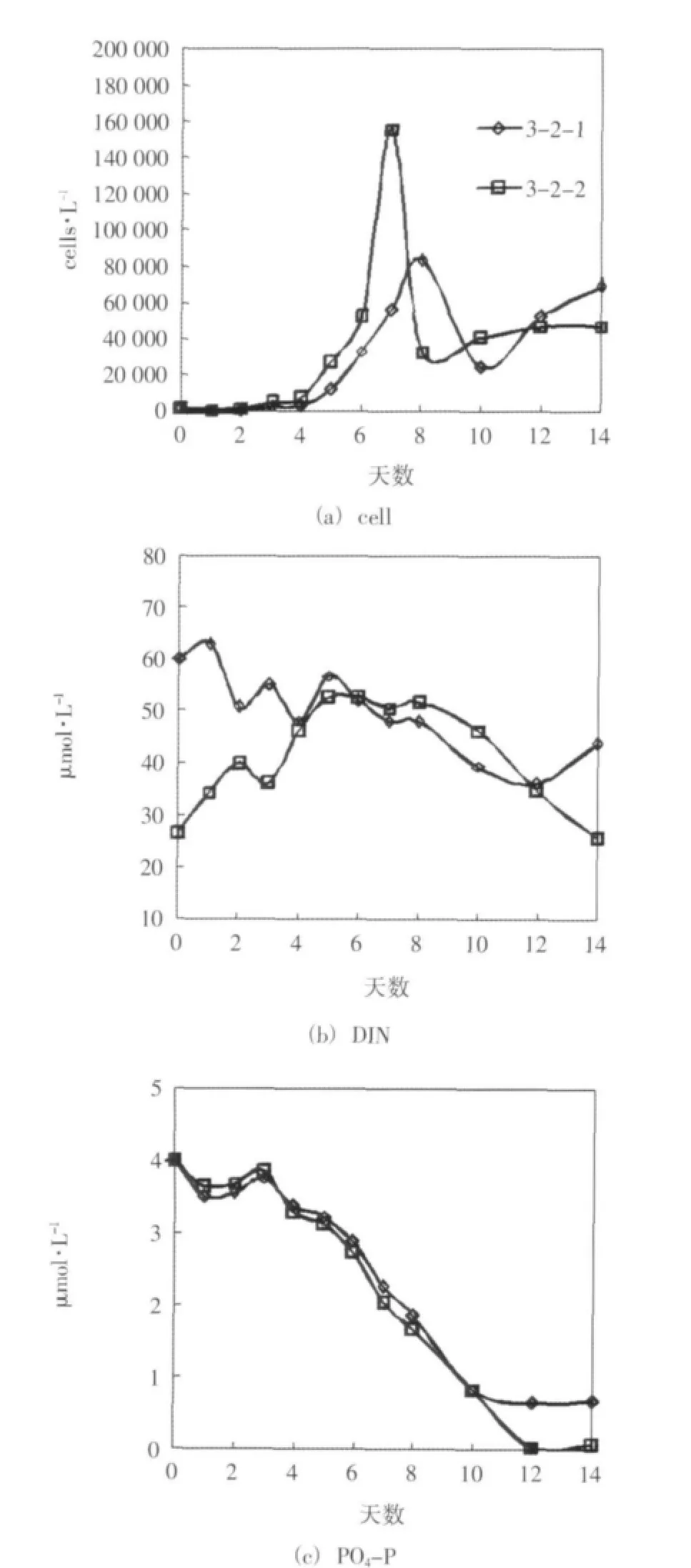
图3 实验三 培养期间细胞密度和营养盐浓度的变化趋势
本文在控制条件下,对渤海湾西南部海域典型站位表层水体进行了模拟现场的加富培养实验,研究了水体的营养盐限制类型以及不同氮磷比和不同硝酸盐添加方式的生态响应。结果表明水体处于磷限制,这与海域的实际情况一致。同时,实验结果表明,在保持氮充足的情况下变化磷酸盐浓度时,较低的氮磷比(5-15)更能促进浮游植物的生长,根据该结论可以推测,如果渤海湾磷酸盐浓度增大,氮磷比降低,渤海湾的富营养化可能造成更为严重的生态后果。近年来,伴随着水体氮磷比增大,渤海湾浮游植物也出现了总量增大的趋势,这主要还是受多种营养盐,尤其是氮污染加剧的影响,与本文实验条件的情况是不同的。对于不同排污方式实际影响的研究目前还未见报道,因此没有进行对比分析。
图3 实验三 培养期间细胞密度和营养盐浓度的变化趋势
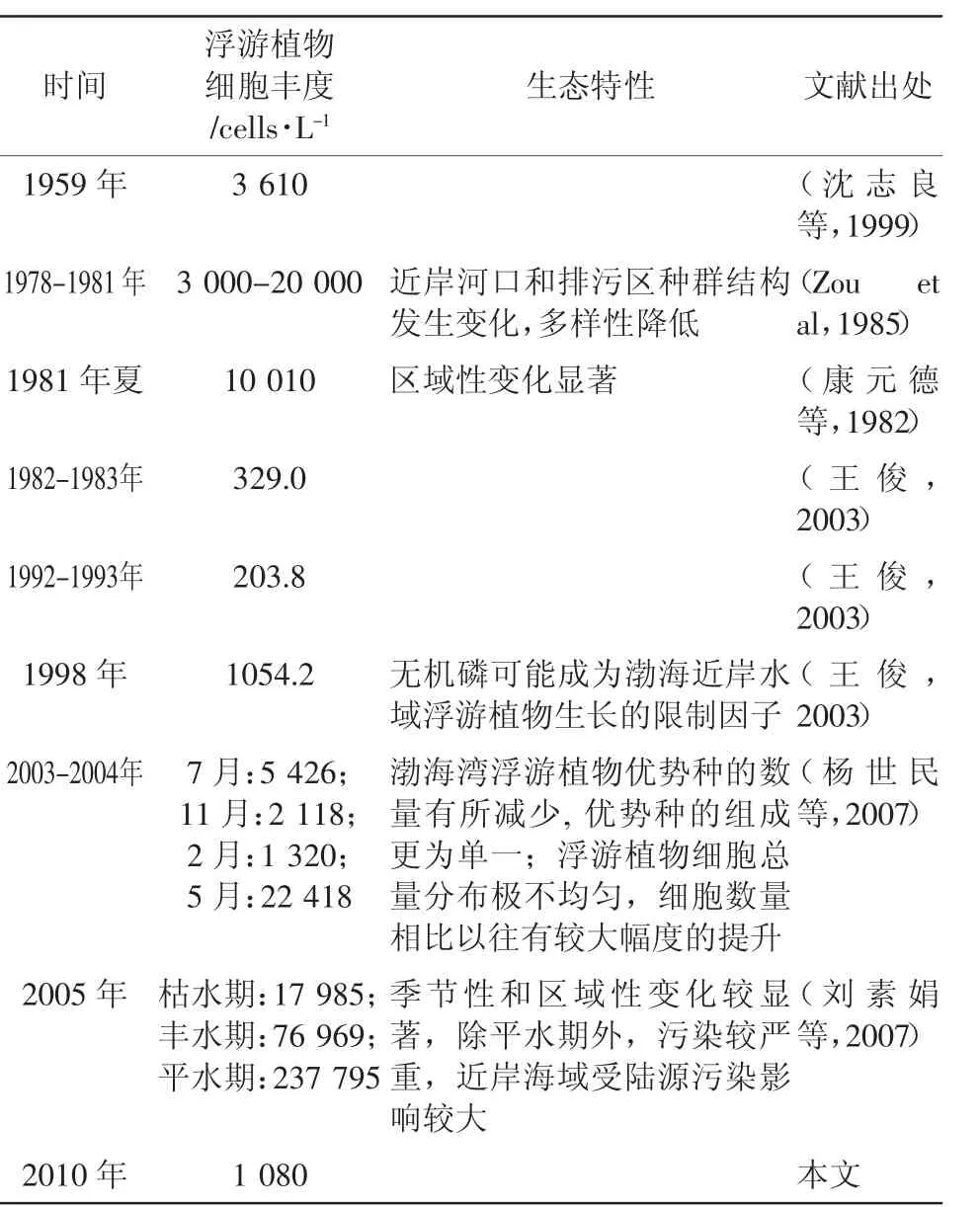
表6 渤海湾浮游植物生态特性的历史变化
本文只是针对一个站位一次采样的研究,可能存在一定的偶然性,实验结果并不能完全代表海域的实际情况,但是在一定条件下揭示了浮游植物生长的重要规律,对于预测和控制海域富营养化具有重要意义。在进一步的研究中应加大采样范围和密度,同时采用更贴近现场环境的培养条件,并对浮游植物的种类变化加以细致的研究。
3 结论
2010年10月,对渤海湾西南部海域典型站位表层水体进行了营养盐添加的模拟现场培养实验,实验判定了水样的营养盐限制类型,并在控制条件下,研究了不同氮磷比和不同硝酸盐添加方式的生态响应,得到结果如下:
(1)单一添加磷酸盐的实验组浮游植物生长最明显,且空白对照组磷酸盐最早被耗尽,水样体现了磷限制的特性。
(2)浮游植物的增长在总体上随着氮磷比的降低而增大,最适宜氮磷比在5~15左右,略低于Redfield比。
(3)硝酸盐的连续性添加比一次性添加更有利于浮游植物的生长,相对于高浓度冲击性氮污染,低浓度持续性氮污染可能会产生更严重的生态影响,此处用一次性添加培养实验可能会低估浮游植物的增长潜力。
了解海域的营养盐限制特性对于海域水生态环境管理和修复具有重要的理论和实际意义,对于渤海湾的研究还需要大量和长期的实验积累,本次实验只是一个开始。但作为方法的创新,本文特色鲜明,在判定水样营养盐限制类型的基础上,对培养实验进行了改进,研究了水样中浮游植物群落对于不同氮磷比和不同硝酸盐添加方式的生态响应,并得到了较合理的结果。在进一步的研究中应加大采样范围和密度,同时采用更贴近现场环境的培养条件,并对浮游植物的种类变化加以细致的研究。
致谢感谢国家海洋局天津海洋环境监测站的石海明、徐玉山等同志在采样期间的帮助。感谢天津大学和河北工程大学的徐晓甫、孙娇、贾丽英、刘娟、刘巨波等同学在实验期间的帮助与支持。
Dorth Q,Whitledge T E,1992.Does nitrogen or silicon limit phytoplankton production in the Mississippi river plume and nearby regions?Continental Shelf Research,12(11):1293-1309.
Fisher T R,Gustafson A B,Sellner K,et al,1999.Spatial and temporal variation of resource limitation in Chesapeake Bay.Marine Biology,133:763-778.
Hecky R E,Kilham P,1988.Nutrient limitation of phytoplankton in freshwater and marine environments:a review of recent evidence on the effects of enrichment.Limnology and Oceanography,33(4):796-822
Ho K C,Hodgkiss I J,1993.Assessing the limiting factors of red tide by bottle bioassay.Asian Marine Biology,10:77-94.
Hodgkiss I J,Ho K C,1997.Are changes in N:P ratios in coastal waters the key to increased red tide blooms?Hydrobiologia,352(1-3):141-147.
Justic D,Rabalais N N,Turner R E,et al,1995.Changes in nutrient structure of river-dominated coastal waters:stoichiometric nutrient balance and its consequences.Estuarine,Coastal and Shelf Science,40:339-356.
Nelson D M,Tréguer P,Brzezinski M A,et al,1995.Production and dissolution of biogenic silica in the ocean:Revised global estimates,comparison with regional data and relationship to biogenic sedimentation.Clobal Biogeochemical Cycles,9(3):359-372.
Tao J,2002.The eco-hydrodynamic environmental character of Bohai Bay.Proceedings of the Second international workshop on coastal eutrophication,China,1:15-24.
Zou J,Dong L,Qin B,1985.Preliminary studies on eutrophication and red tide problems in Bohai Bay.Hydrobiologia,127:27-30.
陈峰,姜悦,1999.微藻生物技术.北京:中国轻工业出版社.
邓华健,陶建华,2004.浮游植物增长光响应动力学实验研究.河北建筑科技学院学报,21(1):28-30,34.
胡俊,柳欣,张钒,等,2008.台湾海峡浮游植物生长的营养盐限制研究.台湾海峡,27(4):452-458.
阚文静,张秋丰,石海明,等,2010.近年来渤海湾营养盐变化趋势研究.海洋环境科学,29(2):238-241.
康元德,吕培顶,张坤诚,1982.渤海湾浮游植物光合作用强度的测定.海洋湖沼通报,4:47-51.
刘慧,董双林,方建光,2002.全球海域营养盐限制研究进展.海洋科学,26(8):47-53.
刘素娟,李清雪,陶建华,2007.渤海湾浮游植物的生态研究.环境科学与技术,30(11):4-10.
刘媛,曹振锐,黄邦钦,等,2004.东、黄海典型海区浮游植物对营养盐添加的响应.厦门大学学报(自然科学版),43(增刊):147-152.
蒲新明,吴玉霖,2000.浮游植物的营养盐限制研究进展.海洋科学,24(2):27-30.
沈志良,1999.渤海湾及其东部水域的水化学要素.海洋科学集刊,41:51-59.
孙霞,2005.光照对东海赤潮高发区赤潮藻类生长的影响.博士学位论文.青岛:中国海洋大学.
唐松,2010.印度洋海水营养盐添加模拟实验中浮游植物生长的营养盐限制作用.海洋科学,34(7):34-40.
王俊,2003.渤海近岸浮游植物种类组成及其数量变动的研究.海洋水产研究,24(4):44-50.
王晓伟,李纯厚,李占东,等,2007.大亚湾海域春季浮游植物生长的限制性营养元素.中国水产科学,14(5):836-842.
王勇,赵澎,单宝田,2002.胶州湾营养盐限制浮游植物生长的初步模拟现场实验研究.海洋科学,26(10):55-59.
王勇,焦念志,1999.北黄海浮游植物营养盐限制的初步研究.海洋与湖沼,30(5):512-518.
王玉珏,洪华生,王大志,等,2008.台湾海峡上升流区浮游植物对营养盐添加的响应.生态学报,28(3):1321-1327.
韦桂秋,2006.AGP试验在珠江口监测的应用.硕士论文.青岛:中国海洋大学.
萧云朴,李扬,李欢,等,2009.温州南麂列岛海域硅藻、甲藻群落变化与环境因子的关系.海洋环境科学,28(2):167-169,201.
杨世民,董树刚,李锋,等,2007.渤海湾海域生态环境的研究Ⅰ.浮游植物种类组成和数量变化.海洋环境科学,26(5):442-445.
张均顺,沈志良,1997.胶州湾营养盐结构变化的研究.海洋与湖沼,28(5):529-535.
张平,沈志良,2001.营养盐限制的水域性特征.海洋科学,25(6):16-19.
邹立,张经,2001.渤海春季营养盐限制的现场实验.海洋与湖沼,32(6):672-678.
猜你喜欢
工会博览(2022年17期)2022-07-15 07:25:22
海洋通报(2022年2期)2022-06-30 06:06:44
矿产勘查(2020年5期)2020-12-19 18:25:11
钻井液与完井液(2019年4期)2019-10-10 01:57:10
疯狂英语·新读写(2018年3期)2018-11-29 22:37:11
水利技术监督(2017年6期)2017-12-19 13:28:17
电镀与环保(2016年2期)2017-01-20 08:15:25
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
应用海洋学学报(2015年3期)2015-11-22 07:39:10
无机化学学报(2014年12期)2014-02-28 17:33:54
