
衢山岛海域秋季浮游植物群落组成及影响因素
2012-08-14 05:48:48高瑜江志兵朱旭宇周青松陈全震曾江宁
海洋通报 2012年6期
高瑜,江志兵,朱旭宇,周青松,陈全震,曾江宁
(国家海洋局第二海洋研究所 国家海洋局海洋生态与生物地球化学重点实验室,浙江 杭州 310012)
衢山岛位于舟山群岛的中北部,是我国第七大岛,它处于长江、钱塘江入海口外缘,属于舟山渔场的岱衢渔场海域。地理、水文、生物等优越自然条件,使岱衢渔场成为适宜多种鱼类繁殖、生长、索饵、越冬的生活栖息地,其中以大黄鱼尤为出名。但由于舟山群岛位于长江下游,长江、钱塘江等陆地径流带入了大量的营养盐,使得该海域富营养化十分严重(刘雪芹,2005),赤潮爆发频繁。
浮游植物是海洋食物链中最基本的环节,是一些经济鱼类的重要饵料。国内对舟山渔场邻近海域的浮游植物调查研究已有不少的报道。张宾等(2011)对舟山渔场氮磷营养盐对浮游植物的限制进行了研究。魏娜等(2010)对舟山渔场浮游植物的时空分布以及优势度进行了分析。刘子琳等(2001)分析了杭州湾—舟山渔场浮游植物的现存量并且计算了其初级生存力。朱根海等(2009)对衢山岛西北部崎岖列岛的浮游植物和水环境状况进行了研究。此外,还有不少学者对衢山岛海域的渔业资源进行了研究(陈华等,2009;李建生等,2006)。但尚无专门针对衢山岛海域浮游植物的研究,故笔者于2010年秋季大小潮期间对衢山岛海域的浮游植物进行调查,并对调查结果做出分析,为深入了解该海区生态系统变化提供基础数据。此外,舟山是浙江实行海洋经济战略的重点区域,舟山的海洋经济必将迎来新的开发热潮,其所面临的海洋环境问题也必将更加突出,浮游植物的变化情况,可以作为水质监测以及评价的重要指标。因此,对该海域浮游植物的调查研究,具有重要的理论和实践意义。
1 材料与方法
1.1 样品的采集与处理
浮游植物样品于2010年秋季大潮(10月22日-10月24日)和小潮(10月6日-10月14日)采集于衢山岛邻近海域23个大面站(图1)。采用网目为76 μm的浅Ⅲ浮游生物网从底至表作垂直拖网采集浮游植物样品,并用流量计计算滤水量。样品用中性甲醛固定,经浓缩后用Leica DM2500显微镜观察、鉴定和计数。水质分析指标分表底两层采集,但考虑到表层指标与浮游生物有较大关系,实际分析时仅采用表层指标(陈雷等,2009)。水质分析项目有水温、盐度、可溶性磷酸盐、可溶性无机氮、悬浮物等。水化学样品采集、处理和测定以及浮游植物样品的采集、贮存、运输、预处理及实验室分析测定均按《海洋监测规范》进行(国家质量技术监督局,1992,1999)。
1.2 数据处理

图1 衢山岛浮游植物采样站位图
分析浮游植物群落结构指数的计算方法(孙军等,2003):(1)香农-威纳多样性指数(H′,Shannon-Wiener index)(2)物种优势度,Y=ni·fi/N0;(3)Pielou均匀度指数,J=H′/log2S。式中:S为每站位所包含的总物种数;N为样品中的总个体数;ni为样品中第i种的个体数。
应用sufer 8.0绘制采样站位图以及细胞丰度图。群落结构分析使用PRIMER 6.0软件完成。
2 结果
2.1 浮游植物物种组成
大、小潮期间衢山岛海域采共有浮游植物7门181种。其中,硅藻133种(占73.9%);甲藻38种(占21.1%),蓝藻4种(2.2%),绿藻和金藻各2种(各占1.1%),黄藻和裸藻各1种(各占0.6%)。
大潮期间,共有浮游植物6门148种,其中硅藻109种(占73.6%),甲藻33种(占22.3%),蓝藻和金藻各2种(各占1.4%),绿藻和裸藻各1种(各占0.7%);小潮期间,共有浮游植物7门114种,种类数低于大潮期间,其中硅藻80种(占70.2%),甲藻26种(占22.8%),蓝藻3种(占2.6%),金藻2种(占1.8%),绿藻、黄藻和裸藻各1种(各占0.9%)。
2.2 浮游植物细胞丰度分布及优势种
大潮期间,衢山岛海域浮游植物细胞丰度为 2.21×104~19 637.25×104个/m3, 平 均 值 为2221.62×104个/m3。浮游植物细胞丰度高值区分布在大衢山北部黄泽山邻近海域(D14、D15、D16、D18和D20站),该区细胞丰度平均值为9 800.82×104个/m3;浮游植物细胞丰度低值区位于大衢山至大长途岛海域(S13、S14、S16、S17和S19站),平均值仅为17.69×104个/m3(图2a)。

图2 大小潮期间衢山岛浮游植物细胞丰度(×104个/m3)(a.大潮;b.小潮)
小潮期间,衢山岛海域浮游植物细胞丰度为5.90×104~58.10×104个/m3,平均值为 29.28×104个/m3。浮游植物细胞丰度分布高值区分布在大衢山至大长途岛海域(S13、S14、S16、S17和S19站),平均值高达42.28×104个/m3;浮游植物细胞丰度低值区位于调查海域西北部(D02、D05、D10、D14和D15站位附近),平均值仅为17.14×104个/m3(图2b)。
本次调查,衢山岛秋季大潮期间浮游植物的绝对优势种为中肋骨条藻(Y=0.987 2),这与魏娜等对舟山渔场海域的研究结果是相符合的。而本次调查区分了大小潮,小潮期间其优势种为中肋骨条藻、琼氏圆筛藻、整齐圆筛藻等。将大潮期间优势种前两位,小潮期间优势种前五位列于表1。
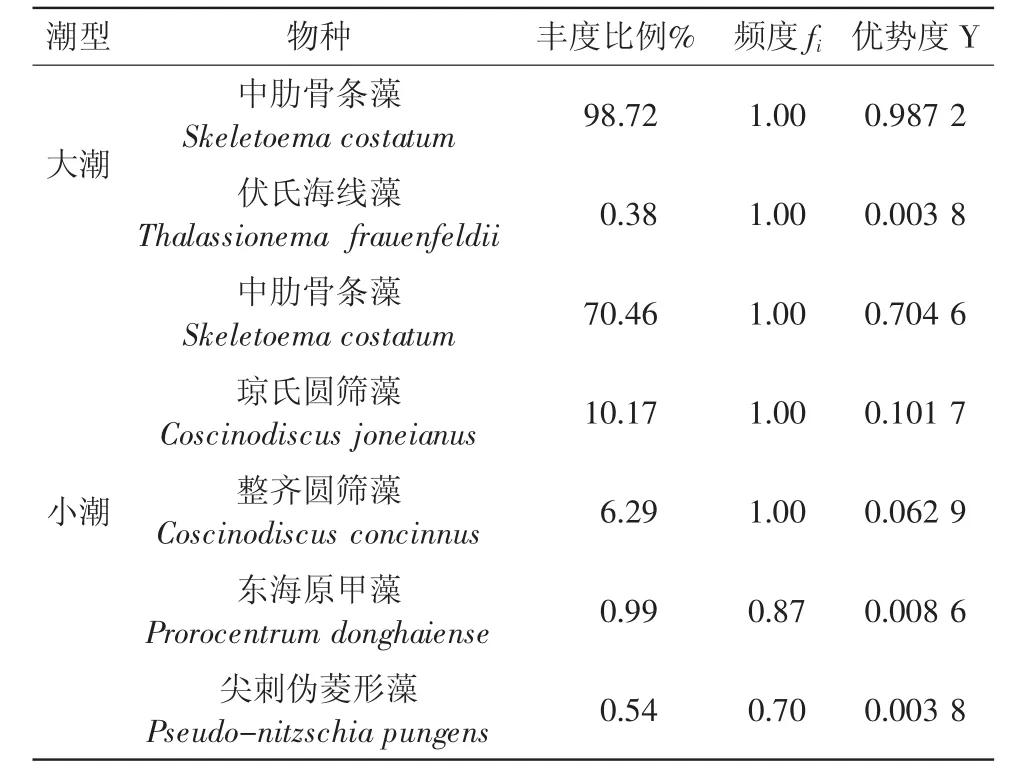
表1 调查站位浮游植物优势种
本次调查海域浮游植物的数量分布和变化具有显著的潮周期变化特点,表现在大潮期间浮游植物种类和细胞丰度明显高于小潮期。大潮期间,细胞丰度的高值区集中于衢山岛北部的D14、D15、D16、D18和D20站,而小潮期间,细胞丰度的高值区分布较为平均,且浮游植物的优势种较为多样。
2.3 群落多样性分析
大潮期间调查区浮游植物的多样性指数介于0.06~2.18,平均值为0.65,最低值出现在D20、D07、D16站;群落均匀度指数介于0.02~0.69,平均值为0.20。小潮期间此海域浮游植物的多样性指数介于0.49~1.74,平均值为1.10,最低值出现在S16站;群落均匀度指数介于0.17~0.57,平均值为0.35。调查海域浮游植物的香农-威纳多样性指数、均匀度指数列于表2。
2.4 群落聚类分析
分别对大小潮23个站位的浮游植物进行Cluster聚类(图3、图4)。Cluster结果以40%的群落结构相似性来划分,则23个站位大潮期间可分为5组,其中D14、D16、D20、D18、D15分为一组,S19和S14 各为一组,S17、S11、S16、D11、S13、S08 分为一组,其余站位分为一组;小潮期间可分为4组,其中S17和D10各为一组,D02、S03、S14、S06、S11、D14、D12分为一组,其余站位分为一组。

表2 大小潮期间衢山岛海域浮游植物群落生物多样性指数

图3 大潮期间衢山岛海域浮游植物群落的聚类图

图4 小潮期间衢山岛海域浮游植物群落的聚类图
大潮期间,群落相似性呈明显斑块分布,D14、D16、D20、D18、D15 5个站位是浮游植物细胞丰度的高值区,位于衢山岛的北部,S19和S14虽在聚类中分属不同的类群,但都是浮游植物细胞丰度的低值区,且位于衢山岛的东南部;小潮期间,群落相似性分布的规律性不明显。由此可见,潮汐对浮游植物的种类组成和细胞丰度都有着显著影响。
2.5 衢山岛海域水质情况
大潮期间,衢山岛海域无机氮分布情况为衢山岛北部高于南部(图5a),平均浓度为0.81 mg/L,最高值位于D16站位,达1.11 mg/L;无机磷含量分布较为均匀(图5b),平均浓度为0.04 mg/L。
小潮期间,无机氮分布同样为衢山岛北部海域稍高于南部(图5c),平均浓度为1.04 mg/L,最高值位于D10站位,达1.18 mg/L;无机磷含量与大潮期无较大差异(图5d),平均值为0.04 mg/L。
根据中华人民共和国海水水质标准(GB 3097-1997),衢山岛海域无机氮含量远远超过四类水标准(无机氮<0.50 mg/L),活性磷酸盐含量也仅仅是三类水范围。由此可见,衢山岛海域富营养化程度较为严重。
在本次调查中发现,大小潮对衢山海域的温度、盐度、pH影响极小,无机氮和活性磷酸盐的含量在大小潮期间的差异也并不显著,但是悬浮物浓度差异却极为明显。大潮期间,衢山岛海域悬浮物浓度平均值为464.02 mg/L,而小潮期悬浮物平均浓度仅为35.16 mg/L。故仅选取无机氮、活性磷酸盐及悬浮物3个参数与浮游植物丰度做相关性分析。结果表明(表3),衢山岛海域浮游植物丰度与无机氮、无机磷含量无显著相关关系,但与悬浮物浓度呈显著的负相关关系。

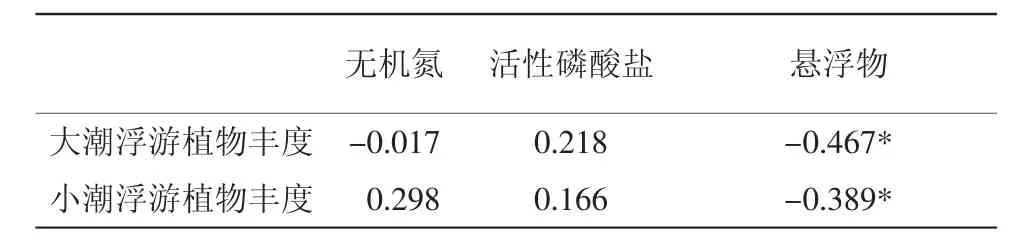
表3 大小潮期间浮游植物丰度与环境因子相关系数
3 讨论
3.1 浮游植物群落与水团、营养盐等环境因子的关系
衢山岛附近海域至少存在3个主要水团,分别为江浙沿岸水、台湾暖流表层水和黄海混合水(张启龙等,2007)。江浙沿岸水低温低盐,而台湾暖流则是高温高盐,寒暖流在此交汇,使得此区域温度适宜,非常适合浮游植物的生长,从而为渔场的形成创造了良好条件。
本海域营养盐的来源一方面是由于长江径流携带大量陆源营养物质,另一方面台湾暖流的入侵也为本海域提供了磷酸盐的补充(Chung et al,1999),因而营养盐并不是该海域浮游植物生长的限制因子。大潮期间,调查海域水体浑浊,透明度低,水环境条件极不稳定,多数种类很难繁殖生长,但是中肋骨条藻却占据了细胞丰度98%的比例,说明中肋骨条藻适宜偏淡水低盐环境,尤其是在径流量偏大水域(唐峰华等,2010)。
根据文献资料(徐兆礼 等,1999;杨世民,2006;王全喜等,2008)及水团因素,将调查区浮游植物分为:(1)近岸种,代表种为中肋骨条藻Skeletoema costatum和琼氏圆筛藻Coscinodiscus joneianus以及伏氏海线藻 Thalassionema frauenfeldii等,在此次调查出现的浮游植物中,这些种类是数量上具有举足轻重的优势种;(2)淡水种,代表性种为黄丝藻Triboneema sp.、颤藻Oscillatoria sp.、具孔盘星藻Pediastrum clathratum、单角盘星藻Pediastrum simplex等。衢山岛海域位于长江冲淡水的稀释区域,这些淡水种主要由长江径流输入,但秋季为长江平水期,故本类浮游植物出现数量均不大;(3)外海高盐种,代表种为虹彩圆筛藻Coscinodiscus oculus-iridis、笔尖形根管藻Rhizosolenia styliformis、密连角毛藻Chaetoceros densus等,本类群则由外海水携带而来。
3.2 浮游植物多样性分析
浮游植物群落的香农-威纳多样性指数、均匀度指数可以作为评价环境质量的依据(陈碧鹃等,2001)。物种组成的多少及每个物种个体数量是衡量生物群落多样性的基础,且各个物种间的均匀程度也是衡量多样性的重要因素(Magurran,1988)。《水生生物监测手册》(国家环保局水生生物监测手册编委会,1993)中指出,当0<H′<1时,表明水体受到人为的重度影响;当H′=1~3时,表明水体受到人为的中度影响。此外,实践应用中,当均匀度J>0.3时,表明海区内浮游植物的多样性较好(马建新等,2002)。现取H′<1且J<0.3作为多样性较差的标准对各站位进行综合评价:大潮期间,除 S13、S17、S19、S14、S08、D05 6 个站外,其他站位的多样性均较低;小潮期间,群落的多样性较大潮有所好转,只有S13、S19、S16、S14、S06、D20、D18、D11、D07站位多样性水平较低。当优势种较多,或无优势性特别突出的种类,此时浮游植物群落结构是稳定的(Pinckney et al,1998),所以,小潮期衢山岛海域浮游植物群落稳定性优于大潮期。
3.3 潮周期对浮游植物群落的影响
衢山岛海域浮游植物的种类和细胞丰度具有明显的潮周期变化特征,大潮期间浮游植物种类以及细胞丰度均大于小潮期,且大潮期间浮游植物总量斑块分布现象比小潮期间明显。衢山岛所处海域位于以强潮和流作用为主要特点的强动力环境中,涨、落潮历时受到长江和钱塘江入海径流的显著影响(陈沈良,2000),水动力实际上对本海区浮游植物的分布起着一种输移和聚集的作用(顾新根等,1995)。大潮期潮流急、流量大,在此潮流作用下位于外海浮游植物向近岸扩展,而小潮期间潮流流速相对较缓,此时浮游植物随水流向外海扩散,由此导致了浮游植物在大小潮期间数量的差异。
4 结论
(1)大、小潮期间衢山岛海域共鉴定出浮游植物7门180种。其中,硅藻133种,占总种数的73.89%,是衢山岛海域浮游植物群落的主要类群;甲藻38种,占总种数的28.50%;蓝藻4种,绿藻和金藻各2种,黄藻和裸藻各1种。大潮期浮游植物种类数大于小潮期。
(2)大潮期间,衢山岛海域浮游植物平均细胞丰度为2 221.62×104个/m3,中肋骨条藻为绝对优势种;小潮期间,浮游植物平均细胞丰度为29.28×104个/m3,此时出现了中肋骨条藻、琼氏圆筛藻、整齐圆筛藻、东海原甲藻等较为多样的优势种。
(3)大潮期间,衢山岛海域的多样性指数、均匀度指数、丰富度指数的均值分别为0.65、0.20、2.56;而小潮期间,其三大指数的均值分别为1.10、0.35、2.75。因此总体来说,衢山岛海域小潮期间的生物多样性和丰富度尚好,群落结构较为稳定。但大潮期间的生物多样性较差。
(4)通过聚类分析,大潮期间衢山岛海域浮游植物群落可以分为5个类群。小潮期间可以分为4个类群。不同类群之间差异性显著。
(5)影响衢山岛海域浮游植物种类和数量分布的决定性因素是潮汐,光照是影响浮游植物丰度的重要因素,而营养盐并没有成为该海域浮游植物生长的限制因子。
Chung S W,Jan S,Liu K K,2001.Nutrients fluxes through the Taiwan Strait in spring and summer 1999.J Oceanogr,57(1):47-53.
Magurran A E,1988.Ecological diversity and its measurement.Sydney:Croom Helm,54:7-46.
Pinckney J L,Paerl H W,Harrington M B,et al,1998.Annual cycles of phytoplankton community-structure and bloom dynamics in the Neuse River Estuary,North Carolina.Marine Biology,131(2):371-381.
陈碧鹃,陈聚法,崔毅,等,2001.莱州湾东部养殖区浮游植物的生态特征.海洋水产学研究,22(3):64-70.
陈华,徐兆礼,2009.舟山渔场衢山岛海域春夏季鱼类数量变化.海洋渔业,31(2):179-185.
陈雷,徐兆礼,姚炜民,等,2009.瓯江口春季营养盐、浮游植物和浮游动物的分布.生态学报,29(3):1571-1577.
陈沈良,2000.崎岖列岛海区的水文泥沙及其峡道效应.海洋学报,3(22):123-131.
顾新根,袁骐,沈焕庭,等,1995.长江口最大浑浊带浮游植物的生态研究.中国水产科学,2(1):16-27.
国家环保局水生生物监测手册编委会,1993.水生生物监测手册.南京:东南大学出版社.
国家质量技术监督局,1999.海洋监测规范第4部分:海水分析(GB17378-1998).北京:中国标准出版社.
国家质量技术监督局,1992.海洋监测规范第6部分:海洋生物调查(GB/T 12763-1991).北京:中国标准出版社.
李建生,李圣法,程家骅,2006.长江口渔场鱼类组成和多样性.海洋渔业,28(1):37-41.
刘雪芹,2005.舟山近岸海域富营养化评价.海洋湖沼通报,2:55-60.
刘子琳,宁修仁,蔡昱明,2001.杭州湾—舟山渔场秋季浮游植物现存量和初级生产力.海洋学报,23(2):93-99.
马建新,郑振虎,李云平,等,2002.莱州湾浮游植物分布特征.海洋湖沼通报,4:63-67.
孙军,刘东艳,宁修仁,等.2003.2001/2002年夏季南极普里兹湾及其邻近海域的浮游植物.海洋与湖沼,34(5):519-532.
孙军,刘东艳,2002.中国海区常见浮游植物种名更改初步意见.海洋与湖沼,33(3):271-286.
唐峰华,伍玉梅,樊伟,等,2010.长江口浮游植物分布情况及与径流关系的初步探讨.生态环境学报,19(12):2934-2940.
王全喜,曹建国,刘妍,等,2008.上海九段沙湿地自然保护区及其附近水域藻类图集.北京:科学出版社.
魏娜,胡颢炎,毛宏跃,等,2010.舟山渔场及其邻近海域浮游植物生态调查与研究.海洋环境科学,29(2):170-173.
徐兆礼,白雪梅,袁骐,等,1999.长江口浮游植物生态研究.中国水产科学,5(6):52-54.
杨世民,董树刚,2006.中国海域常见浮游硅藻图谱.青岛:中国海洋大学出版社.
张宾,章飞军,郭远明,等,2011.舟山渔场海域春夏季氮磷分布及其对浮游植物的限制.浙江海洋学院学报(自然科学版),30(3):185-189.
张启龙,王凡,赵卫红,等,2007.舟山渔场及其邻近海域水团的季节特征.海洋学报,29(5):1-9.
朱根海,施青松,张健,等,2009.崎岖列岛附近海域浮游植物与水环境状况研究.海洋学报,31(4):149-158.
猜你喜欢
少先队活动(2022年10期)2022-12-09 08:58:54
——全球首台3000吨超级浆态床锻焊加氢反应器成功登陆鱼山岛
大陆桥视野(2020年6期)2020-07-02 04:18:30
东坡赤壁诗词(2019年4期)2019-09-12 03:53:06
海峡姐妹(2019年1期)2019-03-23 02:42:54
作文周刊·高一版(2018年24期)2018-09-16 11:22:30
快乐语文(2018年13期)2018-06-11 01:18:12
作文周刊(高考版)(2018年4期)2018-02-06 21:26:33
时尚北京(2017年3期)2017-04-05 23:50:30
中国卫生(2015年1期)2015-11-16 01:06:02
小哥白尼·军事科学画报(2015年7期)2015-09-20 22:38:14
