
林龄对高寒沙地乌柳光合特性的影响1)
2012-08-09 11:08刘海涛贾志清朱雅娟李清雪
东北林业大学学报 2012年12期
刘海涛 贾志清 朱雅娟 于 洋 李清雪
(中国林业科学研究院荒漠化研究所,北京,100091)
乌柳(Salix cheilophila)是杨柳科柳属落叶灌木或小乔木,为中国特有种,主要分布在甘肃、西藏、陕西、宁夏、青海等地,多生长在海拔750~3 000 m的地区[1]。乌柳的生存能力很强,耐寒抗旱,较耐风蚀、沙埋和轻度盐渍化,在我国环境恶劣的西北沙区,发挥了显著的水土保持、防风固沙、改善环境的功能。目前有关乌柳的研究较少,主要集中在沙区造林技术[2]、水分生理[3-4]、氮素利用效率[5]、群落特征[6]、土壤种子库[7]等方面,对其光合生理特性的研究尚未见报道。光合作用是植物最重要的生理过程,决定了植物生长发育的基础和生产力的高低[8],作为绿色植物对各种内外因子最敏感的生理过程之一,能有效表征植物生长环境的变化,很大程度上反映了植物在环境中的生存竞争能力,可作为评价其生产力和适应性的重要指标[9-10]。近年来,随着生理生态测试仪器不断更新发展,同步测量气体交换和叶绿素荧光,已成为生理生态学研究的热点[11-13]。
乌柳引入共和盆地历史虽不长,但已成为常用的防护林带造林树种之一,对当地的流沙治理及生态环境恢复起到了重要作用。为了明确乌柳在光合荧光特性方面如何与当地特殊的高寒干旱环境相适应,不同林龄乌柳的光合生理有多大差异,以及处于哪个龄级的乌柳有着更强的生存竞争能力,笔者以共和盆地不同林龄乌柳为研究对象,通过观测其自然状态下光合荧光特性参数,对各林龄乌柳的光合能力进行对比分析,以期深入了解不同林龄乌柳光合生理差异,为高寒干旱、半干旱荒漠区乌柳人工林的植被建设、保护与经营提供参考。
1 研究区概况
研究区位于青海共和盆地沙珠玉乡治沙示范区内,地理坐标 N36°03'~36°40',E99°45'~100°30',海拔2 871~3 870 m。该区具有显著的高原大陆性气候特征,干旱、大风、寒冷是其主要的气候特点。1月平均气温-12.5℃,7月平均气温14.6℃,年均温2.4℃;年均降水量246.3mm,年均潜在蒸发量1 716.7 mm,无霜期91 d,太阳总辐射6 631.69 MJ·m-2·a-1;全年大风日数平均50.6 d,最多可达97 d,风向主要为西、西北风,平均风速2.7 m·s-1,最大风速可达40 m·s-1,下垫面主要由固定、半固定沙丘及流动沙丘组成[14-15]。
2 材料与方法
2.1 材料
本研究选取4个林龄的乌柳人工灌木林,造林时间分别为1974、1986、2000和2007年,造林方式均为1 m×2 m行带式插干造林。样地位于丘间地中,土壤质地为风沙土。
2.2 方法
2.2.1 光合作用参数日变化测定
测定仪器为Li-6400便携式光合仪。在每个林龄乌柳样地内选取3株长势一致、健康的植株作为标准株进行各项指标的测定。于2011年7月下旬晴天进行测定,选择各待测样株树冠南向中上部中等大小、健康的叶片作为标准叶样。日变化测定时间为08:00—18:00,每2 h进行一次活体测定,测定时每个叶片记录5次数据,取平均值。净光合速率(Pn)、胞间CO2摩尔分数(Ci)、蒸腾速率(Tr)、气孔导度(Gs)等生理指标及光合有效辐射(PAR)、大气CO2摩尔分数(Ca)、气温(Ta)、叶温(Tl)、大气相对湿度(RH)、叶片蒸汽压亏损(Vpdl)等微气象参数由Li-6400同步测出。计算瞬时水分利用效率(WUEt)=Pn/Tr和气孔限制值(Ls)=1-Ci/Ca[16]。测量重复3次。
2.2.2 光响应参数测定
光响应参数测定于7月下旬晴天进行。上午09:00左右,利用Li-6400自带的2 cm×3 cm红蓝光源叶室(Li-6400-02B)提供不同的光合有效辐射:2 000、1 800、1 500、1 200、1 000、800、600、400、200、150、100、80、50、20、0 μmol·m-2·s-1,测定不同光强下的Pn。根据沙漠腹地高温、高辐射的环境气候特点,测定时叶温设置为28℃,参比室CO2摩尔分数为380μmol·mol-1,重复3次。以Michaelis-Menten模型对光响应参数进行拟合[17]。模型表达式为:
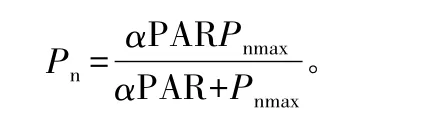
式中:Pn为净光合速率;α是光响应曲线的初始量子效率;PAR为光合有效辐射;Pnmax为最大净光合速率;Rd为暗呼吸速率。模型参数估计用SPSS统计分析软件中的非线性逐步回归方法。用下列公式计算光补偿点(LCP)和光饱和点(LSP):

2.2.3 CO2响应参数测定
CO2响应参数测定于7月下旬晴天进行。在下午15:00左右,利用Li-6400自带的CO2注入系统控制不同的 CO2摩尔分数:0、50、80、100、150、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·mol-1,测定不同 CO2摩尔分数下的 Pn。根据沙漠腹地高温、高辐射的环境气候特点,测定时叶温设置为28℃,光合有效辐射为1 600μmol·m-2·s-1,重复3次。以 Michaelis-Menten模型对CO2响应参数进行拟合[18]。模型表达式为:
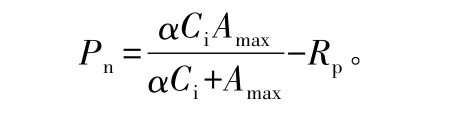
式中:Pn为净光合速率;Ci为胞间CO2摩尔分数,α是CO2响应曲线的初始羧化效率;Amax为光合能力;Rp为光呼吸速率。由于光下暗呼吸很弱,可近似将光下叶片向空气中释放CO2的速率看做光呼吸速率[19]。对低摩尔分数 CO2(Ci小于 200 μmol·mol-1)下的响应参数进行线性回归,回归方程为:Pn=-Rp+CE×Ci。式中,CE为表观羧化效率。当Pn=0时,Ci即为 CO2补偿点(CCP);当 Pn=Amax时,Ci即为CO2饱和点(CSP)。
2.2.4 叶绿素荧光参数日变化测定
利用便携式调制叶绿素荧光仪PAM-2100测定初始荧光(Fo)、最大荧光(Fm)、最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)等叶绿素荧光参数。各参数值均在选定模式下系统自动计算生成,每次测定前叶片暗适应30 min。测定时间与光合生理参数日变化测定同步。重复3次。同时测定凌晨06:00 的 Fo、Fm,Fv/Fm,此时阳光尚未直射到植物体上,叶片经过一整夜的暗适应,PSⅡ反应中心处于充分活化状态。
2.3 叶面积测定
由于乌柳叶片形状不规则,且单个叶片面积小于Li-6400标准叶室默认的面积,待观测结束后,将叶片剪下装入自封袋中迅速带回实验室,用扫描仪扫描后经图像分析软件Image-Pro Plus 6.0计算出实际叶片面积,回算得出真实的各项生理指标参数。
2.4 数据处理与统计分析
用SPSS 16.0统计分析软件进行数据处理与分析,One-way ANOVA进行不同林龄乌柳光合、荧光参数间的差异显著性检验,若差异显著,采用Duncan法进行多重比较。不同林龄乌柳Pn与主要环境因子及气体交换指标进行双变量相关分析。Excel 2007软件绘图。
3 结果与分析
3.1 不同林龄乌柳净光合速率等生理指标日变化比较
不同林龄乌柳Pn等指标的日均值方差分析结果见表1。由表1可知,4个林龄乌柳 Pn、Tr和WUEt日均值间差异显著(P<0.05)。4和37年生乌柳Pn和Tr日均值显著小于11和25年生乌柳(P<0.05);4年生乌柳WUEt日均值显著高于其它3 个林龄乌柳(P<0.05)。

表1 4个林龄乌柳光合生理参数日均值比较
一天中不同林龄乌柳Pn等光合特征参数随外界环境因子的变化而发生变化,但表现出不同的变化规律(表2)。25和37年生乌柳Pn日变化为双峰曲线,4和11年生乌柳Pn日变化为单峰曲线,各林龄乌柳Pn峰值大小和出现时间各异。4和37年生乌柳Ci总体呈下降趋势,18:00达到最低值;11和25年生乌柳Ci虽然总体也呈下降趋势,但最低值出现时间为14:00—16:00。各林龄乌柳Tr日变化均呈单峰曲线,但峰值大小和出现时间各异。4个林龄乌柳Gs日变化曲线基本一致,均为早晨最高,随后逐渐降低,18:00达到最低值。各林龄乌柳Ls变化曲线均为早晨较低,随后逐渐上升,但4和37年乌柳Ls在18:00达到最高值,而11和25年生乌柳则在14:00—16:00达到最高值。各林龄乌柳WUEt日变化曲线均为早晨较高,随后逐渐降低,12:00—14:00 达到最低值。

表2 4 个林龄乌柳 P n、C i、T r、G s、L s、WUEt等光合生理参数日变化
各林龄乌柳Pn与外界环境因子的相关性结果见表3。25和37年生乌柳 Pn与 PAR、Ta呈负相关,表明其对外界环境条件较敏感:在高温、高光强的环境条件下,气孔关闭以减少对水分的消耗,造成Ci降低,光合原料CO2不足使Pn降低;或是在恶劣的环境条件下,D1蛋白损伤使PSⅡ失活或电子传递链某一载体被破坏,造成光合电子传递效率下降,使Pn降低。25和37年生乌柳Pn与RH呈正相关,表明其对外界水分条件较敏感,易受水分亏缺影响。4、11和25年生乌柳Pn与Ca呈负相关,表明其光合能力较强,在光强逐渐增加的过程中,叶肉细胞光合活性逐渐增大,CO2消耗增多、加快,从而出现这种负相关关系。

表3 不同林龄乌柳P n与主要环境因子间的相关系数
各林龄乌柳Pn与气体交换指标间的相关性结果见表4。37年生乌柳Pn与Gs、Ci正相关,与Ls呈负相关,说明其光合能力有限,细胞内同化力、RuBP羧化酶的数量和活性有限;当外界环境条件适宜,光合速率增加时,细胞中CO2量足够,不会由于CO2供应不足使Pn下降。4、11和25年生乌柳Pn与Gs、Ci负相关,与Ls呈正相关,说明其光合能力较强;当外界环境条件适宜,叶肉细胞光合活性逐渐增大,对CO2消耗增多、加快,这与上述各林龄乌柳Pn与Ca相关性分析结果是一致的。25和37年生乌柳Pn与Tr、VpdL呈负相关,说明其易受到外界环境条件的胁迫。通常,叶片温度升高会使气孔下腔蒸汽压差增大,即VpdL增大,使Tr加快,起到降低叶片温度的作用,但Tr、VpdL增加Pn并没有相应增加,表明大量的水分并没有用于光合作用,而用于叶片降温了,此时高温、高光强可能已经对光合器官造成损伤,导致光化学效率下调。4和11年生乌柳Pn与Tr、VpdL呈正相关,说明植物体在增加蒸腾耗水降低叶片温度的同时,能有效地将机体中的水分加以利用,以增加Pn。

表4 不同林龄乌柳P n与气体交换指标间的相关性分析
3.2 不同林龄乌柳的光响应曲线及特征参数比较
由图1可以看出,采用Michaelis-Menten模型对各林龄乌柳光响应参数进行拟合,结果良好。各林龄乌柳Pn随PAR变化趋势基本一致,在200 μmol·m-2·s-1内,随着 PAR 增强,Pn迅速增大;PAR大于200μmol·m-2·s-1时,Pn缓慢增加,达到一定值后基本稳定。
由表5可以看出,4个林龄乌柳光响应参数中,除Pnmax以外,其它参数值间差异显著(P<0.05)。11年生乌柳α值显著高于其它3个林龄乌柳(P<0.05),这与其LCP在4个林龄中数值最低是相耦合的;11年生乌柳LSP值显著低于25和37年生乌柳(P<0.05),表明其利用强光的能力较弱;4和37年生乌柳Rd值显著高于11和25年生乌柳(P<0.05),表明其生理活性较高。

图1 4个林龄乌柳光合-光响应曲线

表5 不同林龄乌柳净光合速率光响应曲线模拟参数比较
3.3 不同林龄乌柳的CO2响应曲线及特征参数比较
由图2可以看出,采用Michaelis-Menten模型对各林龄乌柳CO2响应参数进行拟合,结果良好。各林龄乌柳Pn随CO2变化趋势基本一致:在200 μmol·mol-1内,随着CO2摩尔分数增加,Pn迅速增大;CO2摩尔分数大于200μmol·mol-1时,Pn缓慢增加,达到一定值后基本稳定。

图2 4个林龄乌柳光合-CO2响应曲线
由表6可以看出,4个林龄乌柳CO2响应参数间均产生了显著差异(P<0.05)。11和25年生乌柳Amax值显著高于4和37年生乌柳(P<0.05);11年生乌柳a值显著低于其它3个林龄的乌柳(P<0.05),与实际观测的各林龄低摩尔分数CO2下Pn值是一致的;4和37年生乌柳CCP和CSP明显低于11和25年生乌柳,表明前者利用低摩尔分数CO2的能力要高于后者,但其利用高摩尔分数CO2的能力却要低于后者;25年生乌柳Rp值显著高于4和11年生乌柳(P<0.05),表明25年生乌柳对强光的耐受性较强。

表6 不同林龄乌柳净光合速率CO2响应曲线模拟参数比较
3.4 不同林龄乌柳叶绿素荧光参数日变化比较
不同林龄乌柳 Fv/Fm、ΦPSⅡ、qP、NPQ 等指标的日均值方差分析结果见表7。可知,各林龄乌柳叶绿素荧光参数日均值间差异显著(P<0.05)。11年生乌柳Fv/Fm日均值显著低于4和37年生乌柳(P<0.05);4年生乌柳ΦPSⅡ日均值显著低于其它3个林龄乌柳(P<0.05);4年生乌柳qP日均值显著低于11和25年生乌柳(P<0.05);4年生乌柳NPQ日均值显著高于其它3个林龄乌柳(P<0.05),37年生乌柳NPQ日均值显著高于11年生乌柳(P<0.05)。

表7 不同林龄乌柳叶绿素荧光参数日均值比较
由表8可以看出,4个林龄乌柳Fv/Fm均表现为早晨较高,午间不断降低,傍晚又有所回升的趋势。ΦPSⅡ和Fv/Fm的变化规律并不一致,表明植物潜在的最大光量子产量并不能代表实际的光量子产量,ΦPSⅡ更能反映植物在特定环境条件下的生理活性状态及PSⅡ电子传递链的实际情况。各林龄qP的变化曲线不尽相同,11和25年生乌柳qP的变化趋势大体一致,均为早晨较高,随后逐渐降低,10:00—12:00降至最低值;4和37年生乌柳则为早晨较低,随后逐渐上升,12:00达到一个峰值,随后快速下降,14:00达到最低值。4个林龄乌柳NPQ变化曲线不尽相同,11、25和37年生乌柳NPQ的变化趋势大体一致,均为早晨较低,随后逐渐上升,10:00—14:00达到最高值;4年生NPQ乌柳则为早晨较高,随后有所降低,10:00达到一个峰值,随后快速上升,14:00达到最高值。

表8 4个林龄乌柳叶绿素荧光参数日变化
4 结论与讨论
植物叶片对光能的吸收、传递与利用能力不仅与外界环境因子有关,也与自身生理状态密切相关[20-21]。4个林龄乌柳Pn日变化曲线不尽相同,25和37年生乌柳Pn日变化为双峰曲线,出现了光合“午休”现象;而4和11年生乌柳Pn日变化为单峰曲线,表明不同林龄乌柳对外界环境因子的变化采取了不同的响应策略。光合“午休”是植物长期进化过程中形成的一种适应干旱环境的方法,这种适应是通过气孔的部分关闭实现的,可以提高植物的水分利用效率[22]。这种适应性的调节方式,对植物体节水与在不利环境中生存是有利的,但对光合作用和有机物的积累却是不利的。各林龄乌柳Pn日均值间产生了显著差异,表明林龄对乌柳的光合能力产生了显著影响。同时,各林龄乌柳表现出了不同的水分利用策略:4年生乌柳Pn虽然较低,但却能充分利用有限的水分,表现出较低的Tr和较高的WUEt,而11和25年生乌柳较高的Pn是以较多的水分消耗为代价的,37年生乌柳的Pn和WUEt均处于较低水平,植株可能已经处于生长发育衰退期了。这种处于不同生长阶段的乌柳所采取各异的水分利用方式是对当地特殊环境的适应,反映了不同林龄乌柳为了维持自身的碳水平衡,获取最大碳收益而采取的不同的生态适应对策。
判断植物体Pn降低的原因是气孔还是非气孔因素,主要看Ci和Ls的变化方向:Ci降低、Ls升高表明气孔因素是主要原因;Ci增高、Ls降低表明非气孔因素起主导作用[23]。在午间高温、高光强的环境条件下,4个林龄乌柳Pn值都出现了不同程度的下降,但下降的原因却是不同的:37年生乌柳14:00时Pn降低主要是由非气孔因素造成的,即叶肉细胞羧化能力降低或电子传递链受阻造成同化力不足而导致Pn降低,此时植物的光合器官可能已经受到损伤;25年生乌柳12:00时Pn降低主要是由非气孔因素造成的,14:00时Pn降低主要是由气孔因素造成的;11年生乌柳12:00时和4年生乌柳14:00时Pn降低都是由气孔因素造成的,即气孔关闭降低Tr以减少对水分的消耗,从而引起光合作用原料CO2供应不足造成的,这是植物体的一种保护调节机制[24]。
植物的光合光响应曲线和CO2响应曲线是研究植物净光合速率与光合有效辐射及CO2浓度之间关系的重要手段,对了解植物光化学过程中光化学效率非常重要。4个林龄乌柳α间的排序为11年生>25年生>37年生>4年生,11年生乌柳的α值显著高于其它3个林龄乌柳;Amax间的排序为25年生>11年生>37年生>4年生,11和25年生乌柳Amax值显著高于4和37年生乌柳。α是表征光能利用效率的重要指标,该值高,说明叶片光能转化效率高及利用弱光进行光合生产的能力强[25];Amax反映了植物叶片光合电子传递和磷酸化活性,该值的大小也能直接表征植物实际的 Pn[18],这两个指标的大小排序与实际观测到的Pn日均值大小排序基本上是一致的,进一步佐证了11和25年生乌柳光合能力较强的推断。同时,4个林龄Rd间的排序为4年生>37年生>25年生>11年生,4和37年生乌柳Rd值显著高于11和25年生乌柳(P<0.05)。由于暗呼吸产生的能量是植物生命活动的基础,在植物的生长、发育过程中具有重要作用[26]。因此,在相似的环境条件下,4和37年生乌柳Rd高,说明其要耗费更多的能量抵御外界不利的环境条件以维持自身的生命活动,这是不利于有机物积累的。
通常,植物吸收的光能只有3条去路:光合作用、叶绿素荧光和热,这3条途径存在着此消彼长的关系,光合作用和热耗散的变化会引起叶绿素荧光的相应变化,所以可以通过叶绿素荧光的观测来探究植物体光合作用和热耗散的情况。Fv/Fm是PSⅡ最大光化学量子产量,其大小反映了植物潜在最大光合能力,正常生理状态下该参数一般介于0.80~0.83,且不受物种和生长条件的影响,当其下降时,代表植物受到了胁迫。本研究中,各林龄乌柳的Fv/Fm值在一天中大多数时间要低于0.80,且日均值均在0.77左右,明显低于很多C3植物PSⅡ最大光化学效率[27]。同时,4和37年生乌柳Fv/Fm的最大值出现在08:00,分别为0.808 和0.798;11和25年生乌柳Fv/Fm的最大值出现在06:00,分别为0.797和0.808。理论上凌晨当太阳光尚未直射到植物体上时Fv/Fm应该是最大的,此时植物体经过一整夜的暗适应,所有电子门均处于开放状态,但4和37年生乌柳06:00的Fv/Fm要低于正常水平,且并不是一天中的最大值,11和25年生乌柳06:00的Fv/Fm也仅在正常水平的边缘。这可能是由于当地特殊的环境条件所造成的:当地属高寒沙地,即使在7月,早晨06:00平均气温也只有4.3℃左右,植物处于低温胁迫的状态中,参与光反应电子传递的各种蛋白质及光合色素并没有被完全活化,PSⅡ潜在活性中心受到抑制,使PSⅡ传递电子的能力减弱,所以会出现凌晨Fv/Fm值较正常值偏低的现象。ΦPSⅡ表征了自然光照条件下实际的光化学效率,可以反映PSⅡ中心的开放程度及植物体实际的光合效率[28];qP表征了植物体吸收的光能用于光合作用的比例,反映了PSⅡ捕获的光量子转化成化学能的效率。4个林龄乌柳的ΦPSⅡ和qP的大小排序均为11年生>25年生>37年生>4年生,4年生乌柳ΦPSⅡ和qP日均值显著低于11和25年生乌柳(P<0.05),表明11和25年生乌柳光合能力较强,不易发生光抑制,这与实际观测的Pn日均值的大小是一致的。NPQ是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散掉的那部分能量,反映了植物耗散过剩光能为热的能力,也就是光保护能力[28-29]。4个林龄乌柳的NPQ排序为4年生>37年生>25年生>11年生,4年生乌柳NPQ日均值显著高于其它3个林龄乌柳(P<0.05),37年生乌柳NPQ日均值显著高于11年生乌柳(P<0.05),4和37年生的NPQ较高,表明其吸收的光能有很大一部分不能用于光合作用,而是以热的形式散失掉了,这与各林龄乌柳qP的数值也是相吻合的。
在自然条件下,林木的光合作用受到多种因素的影响,同一树种的光合能力会随生长发育的阶段而变化。本研究是在同一气候条件下进行的,因此各林龄乌柳光合生理特性的差异主要应是由林龄的变化而引起对环境因子适应能力不同造成的。11和25年生乌柳的光合能力相对较强,但其水分利用效率及热耗散能力却相对较弱,处于不同生长阶段的乌柳表现出了不同的生存适应对策。综合上述研究结果推断,4年生乌柳可能处于生长发育初期,11和25年生乌柳可能处于生长发育成熟期;37生乌柳可能已经处于生长发育衰退期。对处于生长发育初期的乌柳,由于易受外界环境条件的胁迫,要多加强抚育措施,如生长季可对其进行一至两次浇水处理以缓解其干旱胁迫,提高其存活率和适应性;而对处于生长发育衰退期的乌柳要及时进行平茬复壮或采取补种措施以弥补其由于自然死亡而出现的种群数量不断降低,使整个乌柳群落处于一种动态平衡当中,不至于出现逆行演替,使已经固定的沙地重新活化。
[1]中国科学院中国植物志编辑委员会.中国植物志:20卷[M].北京:科学出版社,1984:353.
[2]王生录,杨红文.共和盆地高寒沙区乌柳造林技术研究[J].青海农林科技,2002,22(2):7.
[3]宋云民,包哈申.毛乌素沙地沙柳、乌柳及旱柳蒸腾耗水变化规律的研究[J].林业实用技术,2006(5):8-10.
[4]哈申格日乐,宋云民,李吉跃,等.水分胁迫对毛乌素地区4树种幼苗生理特性的影响[J].林业科学研究,2006,19(3):358-363.
[5]Yuan Z Y,Li L H.Soil water status influences plant ni Trogen use:a case study[J].Plant and Soil,2007,301(1/2):303-313.
[6]朱雪林,黄清麟,张超,等.西藏乌柳群落特征[J].山地学报,2011,29(1):116-122.
[7]石晓东,高润梅,郭晋平,等.庞泉沟自然保护区河岸林群落的土壤种子库特征[J].林业科学研究,2010,23(2):157-164.
[8]许大全.光合速率、光合效率与作物产量[J].生物学通讯,1999,34(8):8-9.
[9]于贵瑞,王秋凤.植物光合蒸腾与水分利用的生理生态学[M].北京:科学出版社,2010:171-172.
[10]Xu D Q.Progress in photosynthesis research:from molecular mechanisms to green revolution[J].Acta Phytophysiologica Sinica,2001,27(2):97-108.
[11]郑淑霞,上官周平.8种阔叶树种叶片气体交换特征和叶绿素荧光特征比较[J].生态学报,2006,26(4):1080-1087.
[12]简在友,王文全,孟丽,等.芍药组内不同类群间光合特性及叶绿素荧光特性比较[J].植物生态学报,2010,34(12):1463-1471.
[13]Bertamini M,Muthuchelian K,Nedunchezhian N.Shade effect alters leaf pigments and photosynthetic responses in Norway spruce(Picea abies L.)grown under field conditions[J].Photosynthetica,2006,44(2):227-234.
[14]李锋,孙司衡.景观生态学在荒漠化监测与评价中应用的初步研究:以青海沙珠玉地区为例[J].生态学报,2001,21(3):481-485.
[15]张登山,冯立孝,雷梅,等.青海沙珠玉高寒区风沙土的数值分类研究[J].中国沙漠,1998,18(1):64-69.
[16]Larcher W.植物生理生态学[M].北京:中国农业大学出版社,1997:93-94.
[17]刘海涛,张川红,郑勇奇,等.抗虫转基因欧洲黑杨苗期光合特性研究[J].北京林业大学学报,2011,33(1):36-43.
[18]Harley P C,Thomas R B,Reynolds JF,et al.Modeling photosynthesis of cotton grown in elevated CO2[J].Plant Cell and Enviroment,1992,15(3):271-282.
[19]蔡时青,许大全.大豆叶片CO2补偿点和光呼吸的关系[J].植物生理学报,2000,26(6):545-550.
[20]Gimenez C,Mitchell V J,Lawlor D W.Regulation of photosynthetic rate of two sunflower hybrids under water stress[J].Plant Physiology,1992,98(2):516-524.
[21]许大全.光合作用测定及研究中一些值得注意的问题[J].植物生理学通讯,2006,42(6):1163-1167.
[22]许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990,26(6):5-10.
[23]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[24]Xu H,Li Y,Xu G Q,Zou T.Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation[J].Plant,Cell and Enviroment,2007,30(4):399-409.
[25]许大全.光合作用效率[M].上海:上海科学技术出版社,2002:33-51.
[26]Lusk CH,Reich P B.Relationships of leaf dark respiration with light environment and tissue nitrogen content in juveniles of 11 cold temperate tree species[J].Oecologia,2000,123(3):318-329.
[27]许大全,张玉忠,张荣铣.植物光合作用光抑制[J].植物生理学通讯,1992,28(4):237-243.
[28]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[29]Ort D R.When there is too much light[J].Plant Physiology,2001,125(1):29-32.
猜你喜欢
防护林科技(2022年5期)2022-08-22
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
智富时代(2019年4期)2019-06-01
智富时代(2019年4期)2019-06-01
Coco薇(2017年5期)2017-06-05
现代电生理学杂志(2015年4期)2015-07-18
浙江理工大学学报(自然科学版)(2015年5期)2015-03-01
