
基于地形因子的森林物种多样性与蓄积量分析1)
2012-08-09 11:08:36杨宇军刘兆刚
东北林业大学学报 2012年12期
杨宇军 李 超 张 锐 刘兆刚
(牡丹江医学院,牡丹江,157011) (东北林业大学)
植物群落的多样性是反映群落组成结构的重要指标[1],物种水平上的生物多样性测度,即物种丰富度指数(Species richness index),是最常用的物种多样性指数。影响物种丰富度分布特点的形成因素可能有多种,如时间(历史)、空间异质性、群落生产力、竞争及捕食作用等,它们在一定程度上都与所研究地理区域的生物丰富度有关[2]。生态、气候特点对物种丰富度及特有性空间分布有更密切的联系[3]。森林蓄积量是指一定森林面积上存在着的林木树干部分的总材积,它是反映一个国家或地区森林资源总规模和水平的基本指标之一,也是反映森林资源的丰富程度、衡量森林生态环境优劣的重要依据,往往与生物多样性相关。近年来,通过地形因子差异来研究植被的空间分布规律,了解和掌握各种因素对植被的影响已成为自然地理学、生态学等领域的重点研究方向之一。地形的细微变化可以通过数字高程模型(digital elevation model,简称DEM)来描述。DEM是对地球表面地形、地貌的一种离散的数字表达[4],可以提取任意尺度下的坡度、坡向、高程等地形因子,因而被广泛应用于地学分析、工程设计和土地利用规划等领域。在生态学和林学研究中,DEM多用于研究区域、景观等大尺度空间异质性对森林景观和植被分布的影响。本研究以凉水自然保护区为例,在群落尺度上,基于DEM提取主要地形因子坡度、坡向,分析不同地形条件下物种多样性与蓄积量的差异,旨在从空间微地形角度为森林可持续经营管理提供依据。
1 研究区域概况
黑龙江省凉水国家级自然保护区位于小兴安岭南坡伊春市带岭区的中心,海拔600 m左右。地理坐标为东经128°47'8″~128°57'19″,北纬 47°6'49″~47°16'10″。林场总面积 6 394 hm2,森林总蓄积量100万m3,森林覆被率98% ,被誉为“红松故乡”和“天然生物实验室”[5]。保护区内有原始成熟林、过熟林面积4 100 hm2,其中红松林面积占80%。1980年经原林业部批准建立,1997年晋升为国家级,主要保护对象为红松针阔叶混交林生态系统。保护区为典型的低山丘陵地貌,最高山峰为保护区北部的岭来东山,海拔为707 m,由北向南逐渐降低,至该区的东南端海拔仅有280.0 m。山地平均坡度为10°~15°,局部地段可出现 20°~40°的陡坡。区内既有处于演替顶极状态的原始阔叶红松林、云冷杉林和兴安落叶松林,又有处于不同演替阶段的次生林,几乎囊括了小兴安岭山脉的所有森林植被类型,它最大限度地保存了第三纪植物群落的结构特征,具有古老的区系发生与群落发生的历史,为人类提供小兴安岭原始红松林生态系统的“本底”资料,具有很高的科学价值。
2 数据资料与处理方法
2.1 数据资料
研究采用的是2009年9月由Riegl LMS-Q560获取的LiDAR离散回波数据,其频率为1 550 nm,激光脉冲的长度是3.5 ns,回波宽度的分辨率为0.15 m,激光离散角度是0.5 mrad,垂直精度达0.15 m,采样间隔为1 ns,采样的点云密度为4.2点/m2,飞行相对航高800 m,平均对地飞行速度是180 km·h-1。数据格式为 LAS(1.0版),坐标投影系统是UTM(Universe Transverse Mercator),参考椭球为WGS84(World Global System),每个激光点包含了激光点三维坐标值、回波强度值、类别值、回波类型等16个信息。通过TerraScan激光雷达数据处理软件的三角自适应算法对点云数据进行预处理和分类,通过规则点采样生成高精度DEM。并结合2009年在凉水国家级自然保护区圆形固定样地(面积0.06 hm2,间距500 m×500 m)(图1)的调查数据。调查内容主要包含样地号、样地面积、地理坐标、海拔、坡度、坡向、郁闭度、每木检尺因子(含胸径、树高、冠幅等)和生物多样性(在样地内四角和中心分别设置5个1 m×1 m的样方统计生物多样性)等,并通过黑龙江省二元材积表计算每个样地的林分蓄积。
2.2 处理方法
数学地形模拟是针对地形地貌的一种数学建模过程,这种建模的结果就是一个数字高程模型(DEM)。自从20世纪50年代后期被采用以来,DEM受到了极大的关注,并得到广泛应用。今天,数字高程模型作为地球表面地形的数字描述和模拟已经成为空间数据基础设施和“数字地球”的重要组成部分[6]。因此,说明坡度、坡向在群落尺度下的作用不能忽视[7]。DEM为群落尺度植被与地形因子关系的研究提供了有力工具。在森林经营中引入DEM,可以精确揭示森林群落组成要素在微地形中的空间分异特征,为从空间的角度探索森林可持续经营的途径提供可能。现在的地形分析方法是先实测离散采样点高程,通过对这些离散采样点进行插值,建立规则格网DEM,再用邻域高程提取局部地形因子[8]。应当指出,这种方法因采样点的数量较少,生成DEM较为粗糙,计算的坡度、坡向实际上是邻近区域的平均值[9],往往误差较大,尤其涉及微地形分析时,这种插值数据局限性明显[10]。

图1 圆形样地分布图
2.2.1 DEM 的生成
本研究直接应用LiDAR点云地面回波数据,基于规则采样点建立高精度DEM。运用TerraScan中的地面点分类算法,即Axelsson改进的不规则三角格网加密方法。首先,由最小邻近区域算法获取一个初始的稀疏不规则三角格网,每次将满足设定的阈值条件的点添加到三角网中;然后,重新构建新的不规则三角格网,并重新计算新的阈值条件,对剩余点进行同样的判断筛选,这样重复多次,直到不再有新点加入为止。并通过数据格式转换由LAS格式生成TXT格式数据。再运用ArcGIS9.3读取TXT格式的LIDAR点云数据,通过重投影实现了LiDAR数据与原GIS矢量辅助数据(林班、小班等)的空间坐标统一和空间位置匹配,为进一步的DEM生成等工作奠定基础。
因数据量巨大需对图像进行分块处理,合并生成区域图。本研究运用ArcGIS9.3中ArcToolbox-3D Analysis Tools-TIN Create TIN-Edit TIN生成TIN格式的数字高程模型,并运用ArcToolbox-3D Analysis Tools-Conversion-From TIN-TIN to Raster实现TIN格式的数字高程模型向DEM的转换和ArcToolbox-Data Management Tools-Raster-Raster Dataset-Mosaic to New Raster实现DEM的合并。最终,运用ArcToolbox-Data Management Tools-Raster-Raster Processing-Resample进行DEM的重采样,实现拼接缝隙的消除,从而生成精细DEM(图2)。

图2 DEM图
2.2.2 地形因子的计算
坡度算法:坡度是点位的函数,除非曲面是平面,否则曲面上不同位置的坡度是不相等的,给定点位的坡度是曲面上该点的法线方向N与垂直方向Z之间的夹角a(图3)。由数学分析知,对曲面Z=f(x,y),其给定点(x0,y0,z0)的切平面方程为:

该点的法线方程为:

其方向数为 fx(x0,y0)、fy(x0,y0)和-1,而垂直Z的方向数为0、0和1,则有:

由坡度的概念知,0≤a≤90°,所以坡度值可由(4)式来确定。
坡向算法:坡向与坡度是互相联系的2个参数,坡度反映斜坡的倾斜程度,坡向反映斜坡所面对的方向。坡向是过格网单元所拟合的曲面片上某点的切平面的法线的正方向在平面上的投影与正北方向的夹角,即法方向水平投影向量的方位角(图4)。坡向图中用b标识。
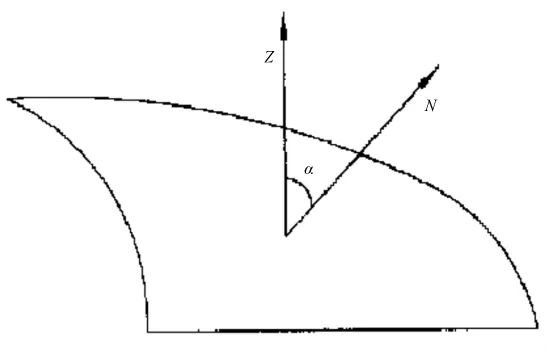
图3 坡度角示意图

图4 坡向角示意图
由数学分析知,设曲面 Z=f(x,y),在点(x0,y0,z0)的切平面方程为:
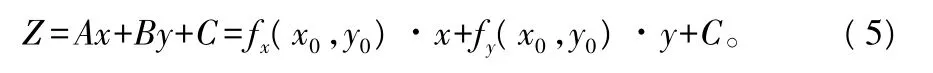
则该点的坡向为:

但根据此式计算的b值在(-π/2,π/2)中取值,而坡向应在(0,2π)中取值,判断b实际值是一项相当繁琐的工作,可以将b的取值根据A、B的取值情况列成表1,通过查表确定最后的坡向值。表中“≈”意味着当A(或B)的绝对值很小时,这与计算时的数值精度要求有关,一般地说,当A或B的绝对值足够小时,其b值趋向于±π/2,因此可以根据情况定一个ξ值,当时|A||B|<ξ,就可以认为|A||B|=0(表1)。

表1 坡向分析
2.2.3 地形因子的分级
利用地区的高程数据生成的该地区的DEM可以直观地看到样地表面形态。利用ArcGIS9.3的空间分析由DEM生成坡度、坡向图(图5、图6),进一步提取地面点对应的坡度、坡向值。地形因子是划分立地条件的重要因子[11],地形因子提取是DEM的重要功能。利用DEM可以提取地形因子坡度、坡向、地表粗糙度、曲面面积、估计特征等地形因子。本研究主要采用DEM的坡度、坡向地形因子提取与分析方法。在群落尺度上,基于DEM提取主要地形因子坡度、坡向[12],为进一步分析地形因子对物种多样性和蓄积量的影响做准备。
3 地形因子对森林生物多样性和蓄积量分布的影响
在群落尺度上,当气候条件基本一致,使群落生境分化的主要因素是光照和土壤条件等因素。地形因子通过对光照、水分、土壤条件的再分配来间接影响植物群落的物种多样性(本文主要研究草本、灌木、乔木的总物种数)和蓄积量。根据各样方物种的丰富度在坡度、坡向因子上的分布范围,结合表1的坡向分析,对坡度和坡向进行划分(根据凉水地区实际数据划分)。坡度分3级:1级0°~5°,2级7°~15°,3 级16°~25°。坡向分8个:E(东坡)方位角范围为68°~112°,SE(东南)方位角范围为113°~157°,S(南)方位角范围为 158°~202°,SW(西南)方位角范围为203°~247°,W(西)方位角范围为248°~292°,WN(西北)方位角范围为 293°~337°,N(北)方位角范围为338°~360°和0°~22°,无坡向坡度<5°。

图6 坡向图
3.1 单地形因子分析
3.1.1 坡度
坡度是在数字高程模型基础上形成的,是数字高程模型的派生模型[13],坡度不仅造成倾斜度上的不同,而且影响土壤的水、热及养分的再分配[14]。不同坡度等级之间,物种丰富度的最大差异为2种,1级坡度的样方平均植物种数约为23个,2级坡度的样方平均植物种数约为24个,而3级坡度约为22个。2级坡度的物种丰富度最高,比3级坡度高6.42%。坡度与蓄积量的关系为:1级坡度的样方平均蓄积量最小,为12.67 m3;2级坡度的样方平均蓄积量最大,为17.41 m3;3级坡度的平均蓄积量为13.47 m3。在相同面积下,2级坡度比1级坡度的平均蓄积量高37.37%。表明,针阔混交林群落中,坡度过大或过小均可引起物种多样性和蓄积量的下降。以2级坡度的物种多样性和蓄积量最高。
3.1.2 坡向
8个坡向中,物种多样性最低的为东坡,样方平均数约为22个,物种多样性最高的为西坡,样方平均数约为25个。物种多样性最高的坡向与最低的坡向之间相差约5个物种(表2)。不同坡向(由于北坡只有一块样地所以不具可比性)的蓄积量顺序为SE>S>NW>SW>E>W>无坡向,最高的是东南坡,样方平均蓄积量为20.38 m3。最低的是无坡向,样方平均蓄积量为11.77 m3。东南坡比无坡向平均蓄积量高73.15%(表2)。

表2 坡向与物种多样性、蓄积量关系
由此可见,凉水自然保护区物种多样性最高的是西坡,最低的是东坡;而蓄积量最高的是东南坡,最低的是无坡向。其他坡向差别不大。这种变化趋势与坡向重新分配太阳辐射所导致的土壤湿度随坡向的变化趋势相吻合[11]。
3.2 地形因子组合分析
对研究区域地形因子进行组合分析,可以揭示不同地形因子与物种多样性和蓄积量的关系。
在各级坡度中,蓄积量的顺序为SE>S>E>N>NW>SW>W>无坡向,在各级坡度中,无坡向均为最低。在各坡向、坡度组合中,无坡向、1级坡度的蓄积量最低,比最高的东南坡、2级坡度的蓄积量平均少118.5%(表3)。东坡、1级坡度的物种多样性最低,平均有20.5个物种,与最高的西坡、2级坡度之间相差约7.5个物种。其他坡向、坡度组合的物种差异不大(表4)。

表3 蓄积量的坡度、坡向分析结果 m3·样地-1

表4 物种多样性的坡度、坡向分析结果
4 结论与讨论
把基于LiDAR数据的高精度DEM提取的主要地形因子——坡度和坡向,引入凉水国家级自然保护区群落多样性与蓄积量差异分析。结果表明:东坡、1级坡度物种多样性最低,平均为20.5个/样方;西坡、2级坡度最高,平均为28个/样方。无坡、3级坡度的平均蓄积量最低,为11.77m3/样方,东南坡、2级坡度最高,为20.38 m3/样方。从单一地形因子角度看,在凉水自然保护区的群落中,坡度过大或过小均可引起物种多样性与蓄积量下降,以2级坡度的物种多样性和蓄积量最高。蓄积量随地形变化比物种多样性敏感,自北坡沿东方向过渡到西坡,蓄积量呈先升高后降低趋势。物种多样性、蓄积量最高的是南坡、东南坡,最低的是西坡、无坡向。说明坡度、坡向等地形因子在小尺度下对生态系统多样性及森林蓄积量的分布的影响不能忽视。高精度DEM为小尺度群落植被与地形因子关系的研究提供了有力工具。在森林经营中引入DEM,可以精确揭示森林群落组成要素在微地形中的空间分异特征,为从空间的角度探索森林可持续经营的途径提供可能。
当然,本研究限于数据量和区域大小,仅对坡度、坡向等地形因子对生物多样性及森林蓄积分布的影响进行了初步分析,为今后大区域的相关研究提供参考。对于森林生态系统来说,其他地形因子如高程、阴坡、阳坡等对其物种多样性和森林蓄积量分布的影响还有待进一步研究。
[1]Magurran A E.Ecological diversity and its measurement[M].Princeton:Princeton University Press,1988.
[2]杨清培,李鸣光,王伯荪,等.粤西南亚热带森林演替过程中的生物量与净第一性生产力动态[J].应用生态学报,2003,14(12):2136-2140.
[3]刘茂松,洪必恭.中国壳斗科的地理分布及其与气候条件的关系[J].植物生态学报,1998,22(1):41-50.
[4]李志林,朱庆.数字高程模型[M].武汉:武汉大学出版社,2003.
[5]任孝臣,彭彦柱.简述黑龙江省红松习性及经营现状[J].林业勘察设计,2005(2):52.
[6]张谧,熊高明,陈志刚,等.神农架米心水青冈-曼青冈群落的地形异质性及其生态影响[J].生态学报,2004,24(12):2686-2689.
[7]胡润田,靳洪生.数字化地形信息及其在森林资源管理中的应用[J].东北林业大学学报,1994,22(2):24-30.
[8]张谧,熊高明,陈志刚,等.数字高程模型在群落内物种共存研究中的应用:以神农架米心水青冈-曼青冈群落的地形模型建立为例[J].植物生态学报,2005,29(2):197-201.
[9]汤孟平,周国模,施拥军,等.不同地形条件下群落物种多样性与胸高断面积的差异分析[J].林业科学,2007,43(6):26-30.
[10]贺金生,马克平.物种多样性[M].杭州:浙江科学技术出版社,1997.
[11]史作民,程瑞梅,刘世荣,等.宝天曼植物群落物种多样性研究[J].林业科学,2002,38(6):17-23.
[12]香宝,刘纪远,张增祥.东亚土地覆盖环境背景数字地面模型研究[J].地理研究,2001,20(6):653-659.
[13]宋永昌.植被生态学[M].上海:华东师范大学出版社,2001.
[14]韩兴吉.油松个体林木生长的数学模拟[J].北京林业大学学报,1988,10(1):1-9.
猜你喜欢
测绘学报(2019年11期)2019-11-20 01:31:42
城市道桥与防洪(2019年5期)2019-06-26 00:56:04
山东林业科技(2018年6期)2019-01-08 09:48:04
山东林业科技(2017年1期)2017-06-29 07:54:06
电气化铁道(2016年4期)2016-04-16 05:59:46
林业与生态(2016年2期)2016-02-27 14:23:42
土壤与作物(2015年3期)2015-12-08 00:46:55
河北遥感(2015年2期)2015-07-18 11:11:14
医学研究杂志(2015年4期)2015-06-10 06:42:43
植物营养与肥料学报(2012年3期)2012-10-26 02:54:52
