
长白山典型森林生态系统土壤呼吸的季节动态1)
2012-06-28 09:12刘颖
东北林业大学学报 2012年11期
刘 颖
(华南理工大学,广州,510640)
韩士杰
(中国科学院沈阳应用生态研究所)
全球气候变暖及其影响是当前人类所面临的最为严重的环境问题之一,陆地生态系统在全球碳循环中起着源/汇的双重作用,是人类活动与气候变化相互作用的重要环节。陆地生态系统碳循环中,土壤呼吸是大气CO2的重要排放源,其排放量是陆地生态系统向大气排放碳的最大通量之一[1]。从土壤呼吸产生的生理学机制看,自养呼吸为纯根呼吸,异养呼吸为土壤微生物呼吸(土壤动物呼吸忽略不计)[2]。在不同的森林生态系统中,由于所处地理位置、土壤类型、性质及群落组成等不同导致土壤呼吸中各组分对土壤呼吸的贡献不同。土壤呼吸及其组分的分离和量化已经成为当今生态系统生态学、碳循环研究、植物生理学、土壤科学和全球气候变化模拟中的一个重要议题[2]。尽管国内外有关土壤呼吸方面的研究很多[3-6],我国有关土壤呼吸组分的量化研究也较多,但是,量化方法存在很大的差异[7-8]。长白山北坡垂直植被带生态系统在我国林业和生态环境建设中占有重要的地位,对该地区森林生态系统土壤呼吸的季节变化进行测定,确定不同组分对土壤总呼吸的贡献,不仅对于评估该地区森林生态系统碳收支非常关键,而且对于评测中国温带森林生态系统在全球碳循环中的功能和地位也有着重要的意义。因此,本研究以长白山典型森林生态系统(阔叶红松林、红松云冷杉林、岳桦云冷杉林和岳桦林)为样地,通过对土壤呼吸、根系呼吸、土壤温度和湿度的测定,研究长白山典型森林生态系统土壤CO2释放规律及其根系对土壤总呼吸的贡献,为进一步揭示土壤CO2释放的过程机理提供基础试验数据。
1 研究区概况
长白山自然保护区位于吉林省东南部安图、抚松和长白3县境内,地理坐标为41°42'45″~42°45'18″N,127°33'30″~128°16'48″E,年平均气温在 6.1℃,年降水量600~900 mm。研究地点位于保护区北坡,由于海拔高度的上升导致小气候变化,在不同海拔梯度上形成了不同的森林类型,从下至上依次为阔叶红松林、红松云冷杉林、岳桦云冷杉林、岳桦林。研究样地分别设在位于中国科学院长白山森林生态系统定位站阔叶红松林、红松云冷杉林、岳桦云冷杉林、岳桦林4个植被类型的标准样地附近,各样地详细情况见刘颖等[9]。
2 研究方法
于2003年5月,在每个样地内用钢桶打5个深30 cm(根据细根分布定)的土柱,将根切断,插入内径10.5 cm的PVC管,并在每个管子旁边分别放置一个内径10.5 cm长4.5 cm的土壤环,于2004年5月—2004年10月,选择晴朗天气,用LI-COR公司生产的LI-6400光合作用分析仪(配带土壤呼吸室),对长白山4个典型森林生态系统(阔叶红松林、红松云冷杉林、岳桦云冷杉林和岳桦林)的土壤呼吸进行测定,测定频率约为1次/周。根据PVC管及土壤环内土壤呼吸的差值计算根系呼吸及其对总土壤呼吸的贡献。土壤5 cm温度由光合系统配带的温度探针测得,湿度用TDR测得,二者测量均与土壤呼吸的测量同步进行。
3 结果与分析
3.1 土壤总呼吸的季节变化
不同森林类型土壤总呼吸的季节变化都比较明显(图1),且阔叶红松林、红松云冷杉林和岳桦云冷杉林变化趋势基本相似,都呈双峰型,从5月份随着温度升高而逐渐上升,并于6月中下旬达到第1个峰值,分别为 5.37、5.72、5.31 μmol·m-2·s-1。此后土壤呼吸速率降低,直至8月中旬达到第2个峰值,分别为5.97、4.74、4.51 μmol·m-2·s-1。此后随温度的降低而下降;岳桦林土壤总呼吸速率随季节变化呈单峰型,7—8月份土壤呼吸速率维持在相对较高水平。4种森林类型土壤总呼吸随季节的变化趋势与该地区的降水及各类型植被所在的地形有关,由于该地区6月中下旬开始出现较大强度的降雨,阔叶红松林、红松云冷杉林和岳桦云冷杉林地形较为平坦,不利于土壤排水,导致土壤湿度过大,不利于土壤微生物和根系的活动,从而使土壤呼吸速率在一段时间内维持在较低水平,直至8月中旬土壤湿度过高现象消失。而岳桦林处于坡度较大的地形,即使降雨强度过大,水分也容易排出,不会造成土壤湿度过大而影响土壤呼吸。阔叶红松林、红松云冷杉林和岳桦云冷杉林在6月中下旬和8月中旬的水热条件较好,适合土壤微生物和根系的活动,因此土壤总呼吸速率出现峰值。岳桦林土壤总呼吸的峰值与土壤温度的最大值同步,出现在7月下旬。实验进行过程中,阔叶红松林、红松云冷杉林、岳桦云冷杉林和岳桦林土壤总呼吸的季节变化范围分别是1.49 ~5.97、1.38 ~5.72、1.06 ~5.31、2.02 ~4.96 μmol·m-2·s-1。

图1 不同森林土壤总呼吸的季节变化
3.2 土壤异养呼吸的季节变化
不同森林类型土壤异养呼吸都存在比较明显的季节变化(图2),且阔叶红松林和红松云冷杉林变化趋势基本相似,都呈双峰型,从5月份随着温度升高而逐渐上升,并于6月中下旬达到第1个峰值,二者均为 2.82 μmol·m-2·s-1。此后土壤呼吸速率降低,直至8月中旬达到第2个峰值,分别为2.91、2.73 μmol·m-2·s-1。此后随温度的降低而下降;岳桦云冷杉林和岳桦林土壤异养呼吸速率随季节变化相似,均呈单峰型,而且峰不很明显,7—8月份土壤异养呼吸速率维持在相对较高水平,其它时间较低。造成4种森林类型土壤异养呼吸随季节的变化趋势的原因与土壤总呼吸相同,即主要是由各类型植被所在地的土壤温度和水分共同作用的结果。试验进行过程中,阔叶红松林、红松云冷杉林、岳桦云冷杉林和岳桦林土壤异养呼吸的季节变化范围分别是0.81 ~2.91、0.85 ~2.82、0.61 ~2.26、1.19 ~2.67 μmol·m-2·s-1。

图2 不同森林土壤异养呼吸的季节变化
3.3 根系呼吸及其对土壤总呼吸贡献的季节变化
根呼吸(森林土壤自养呼吸)与林木地上光合作用所能提供的同化物密切相关。林木根系呼吸有明显的季节变化,一般生长季较高,休眠季节较低。长白山不同森林类型根系呼吸的季节变化都比较明显(图3),且阔叶红松林、红松云冷杉林和岳桦云冷杉林变化趋势基本相似,都呈双峰型。从5月份随着温度升高而逐渐上升,并于6月中下旬达到第1个峰值,分别为 2.55、2.90、3.20 μmol·m-2·s-1。此后根系呼吸速率降低,直至8月中下旬达到第2个峰值,分别为 3.06、2.25、2.32 μmol·m-2·s-1。此后随温度的降低而下降;岳桦林根系呼吸速率随季节变化而变化的幅度较小,呈单峰型。4种森林类型根系呼吸随季节的变化趋势与该地区的降水及各类型植被所在的地形有关,由于该地区6月中下旬开始出现较大强度的降雨,阔叶红松林、红松云冷杉林和岳桦云冷杉林地形较为平坦,不利于土壤排水,导致土壤湿度过大,抑制了根系的生命活动,从而使根系呼吸速率在一段时间内维持在较低水平,直至8月中旬这种土壤湿度过高现象消失,而岳桦林处于坡度较大的地形,即使降雨强度过大,水分也容易排出,不会造成土壤湿度过大而影响根系呼吸。阔叶红松林、红松云冷杉林和岳桦云冷杉林在6月中下旬和8月中旬的水热配置条件较好,适合根系的活动,因此根系呼吸速率出现峰值。岳桦林根系呼吸的峰值与土壤温度的最大值同步,出现在7月下旬,为 2.29 μmol·m-2·s-1。试验进行过程中,阔叶红松林、红松云冷杉林、岳桦云冷杉林和岳桦林根系呼吸的季节变化范围分别是0.79 ~3.06、0.49 ~2.90、0.40 ~3.20、0.83 ~ 2.29 μmol·m-2·s-1;平均值分别为 1.60、1.58、1.49、1.66 μmol·m-2·s-1。
根系对土壤总呼吸贡献的季节变化与根系呼吸的季节变化相似(图3)。阔叶红松林根系呼吸对土壤总呼吸的贡献随季节变化呈双峰型,峰值分别出现在7月中旬和9月上旬,变化范围是28.5% ~60.1%,平均为43.6%;红松云冷杉林在6月上旬至9月上旬根系对土壤总呼吸的贡献变化幅度比较小,基本维持在一定的水平,试验进行过程中的变化范围是 29.6% ~51.2%,平均为 44.1%;岳桦云冷杉林仅在7月上旬出现一个峰值,变化范围是34.4% ~60.3%,平均为 45.5%;岳桦林的季节变化范围是36.2% ~48.7%,平均为44.4%。
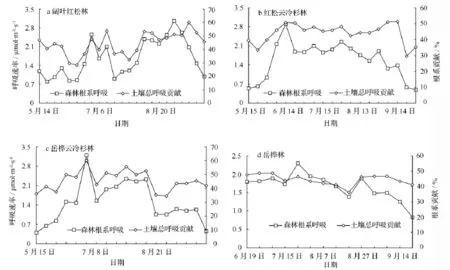
图3 不同森林根系呼吸及其对土壤总呼吸贡献的季节变化
4 结论与讨论
许多研究表明,土壤呼吸速率在生长季不同月份也表现明显的变化格局。如Fang等[10]发现佛罗里达州26年生的湿地松人工林土壤呼吸速率在10月最高,而在1月最低;Billings等[11]发现成熟的北方森林在7月前土壤呼吸速率较低,在8月最高;Xu等[12]发现土壤呼吸速率最高值在6月和11—12月,最低值在中夏;Davidson等[13]发现土壤呼吸速率在6—7月最高,在8月中旬到9月初下降。土壤呼吸速率季节性差异主要受土壤温度和土壤湿度变化的共同影响,但温度和湿度对土壤呼吸速率的影响程度不同。通常夏天的温度较高,土壤呼吸速率应该较高,但夏天的干旱和洪涝可能会限制土壤呼吸而抵消温度升高对土壤呼吸的促进作用。本研究结果表明,不同森林类型土壤总呼吸、断根土壤呼吸和根呼吸的季节变化都比较明显,阔叶红松林、红松云冷杉林和岳桦云冷杉林变化趋势基本相似,都呈双峰型,并于6月中下旬和8月中旬达到峰值。岳桦林呈单峰型,7—8月份土壤呼吸速率维持在相对较高水平,其它时间较低。造成4种森林类型断根土壤呼吸随季节的变化趋势的原因主要是由各类型植被所在地的土壤温度和水分共同作用的结果。阔叶红松林、红松云冷杉林在6月下旬至8月上旬,由于较大强度的降雨和地形不利于排水,导致土壤湿度过大,降低了土壤孔隙的通透性,O2是植物根系和土壤微生物进行有氧呼吸的必要条件,过高的土壤含水量限制土壤中的O2扩散,此时土壤处于嫌气状态,植物根系和好氧微生物的活动受到抑制,土壤有机质的分解速率降低,土壤中产生的CO2减少,从而使土壤呼吸速率在一段时间内维持在较低水平。
长白山4种森林类型根系对土壤总呼吸贡献的季节变化与根系呼吸的季节变化相似。4种森林根系呼吸对土壤总呼吸的贡献的变化范围是28%~60%。根系呼吸占土壤呼吸的10% ~90%,主要依赖于所研究的生态系统类型、测定季节和所用的测量方法[2]。根系呼吸对土壤总呼吸的贡献在北方生物群落中较高,其中北极冻原为50% ~93%[14]、北方森林为62% ~89%[15];在温带,根系呼吸占土壤呼吸的比例阔叶林为33% ~50%[16],针叶林为35% ~62%[17]。而世界范围内根系呼吸占土壤总呼吸的比例主要集中在40% ~60%[18-20]。与上述结果进行比较,本实验所测得的结果刚好处于北方温带森林范围内。
[1]Schlesinger W H,Andrews J A.Soil respiration and the global carbon cycle[J].Biogeochemistry,2000,48(1):7-20.
[2]Hanson P J,Edwards N T,Garten C T,et al.Separating root and soil microbial contributions to soil respiration:a review of methods and observations[J].Biogeochemistry,2000,48(1):115-146.
[3]Liu Ying,Han Shijie,Lin Lu.Seasonal changes of soil respiration in Betula platyphylla forest in Changbai Mountain,China[J].Journal of Forestry Research,2009,20(4):367-371.
[4]孟春,王俭.非主要生长季节白桦人工林林隙内伐根对土壤呼吸速率的影响[J].东北林业大学学报,2010,38(11):66-68.
[5]杨阔,王传宽,焦振.东北东部5种温带森林的春季土壤呼吸[J].生态学报,2010,30(12):3155-3162.
[6]张晓静,关德新,吴家兵,等.土壤温湿度对长白山阔叶红松林不同土壤层呼吸速率的影响[J].东北林业大学学报,2010,38(11):68-71.
[7]陆彬,王淑华,毛子军,等.小兴安岭4种原始红松林群落类型生长季土壤呼吸特征[J].生态学报,2010,30(15):4065-4074.
[8]刘颖,韩士杰.长白山四种森林土壤呼吸的影响因素[J].生态环境学报,2009,18(3):1061-1065.
[9]刘颖,韩士杰,林鹿.长白山四种森林类型凋落物动态特征[J].生态学杂志,2009,28(1):1-6.
[10]Fang C,Moncrieff J B,Gholz H L,et al.Soil CO2efflux and its spatial variation in a Florida slash pine plantation[J].Plant and Soil,1998,205:135-146.
[11]Billings S A,Richter D D,Yarie J.Soil carbon dioxide fluxes and profile concentrations in two boreal forest[J].Canadian Journal of Forest Research,1998,28:1773-1783.
[12]Xu Ming,Qi Ye.Soil-surface CO2efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California[J].Global Change Biology,2001,7:667-677.
[13]Davidson E A,Beld E,Boone R D.Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J].Global Change Biology,1998,4(2):217-227.
[14]Raich J W,Tufekcioglu A.Vegetation and soil respiration:correlations and controls[J].Biogeochemistry,2000,48:71-90.
[15]Rey A,Pegoraro E,Tedeschi V,et al.Annual variation in soil respiration and its components in a coppice oak forest in Central Italy[J].Global Change Biology,2002,8(9):851-866.
[16]Nakane K,Kohno T,Horikoshi T.Root respiration rate before and just after clear-felling in a mature,deciduous,broad-leaved forest[J].Ecological Research,1996,11:111-119.
[17]Striegl R G,Wickland K P.Soil respiration and photosynthetic uptake of carbon dioxide by ground-cover plants in four ages of jack pine forest[J].Canadian Journal of Forest Research,2001,31(9):1540-1550.
[18]Singh B,Nordgren A,Löfvenius M O,et al.Tree root and soil heterotrophic respiration as revealed by girdling of boreal Scots pine forest:extending observations beyond the first year[J].Plant Cell and Environment,2003,26:1287-1296.
[19]Subke J A,Inglima I,Cotrufo M F.Trends and methodological impacts in soil CO2efflux partitioning:a metaanalytical review[J].Global Change Biology,2006,12:921-943.
[20]Yi Zhigan,Fu Shenglei,Yi Weimin,et al.Partitioning soil respiration of subtropical forests with different successional stages in south China[J].Forest Ecology and Management,2007,243:178-186.
猜你喜欢
格言·校园版(2022年17期)2022-07-06
新农业(2022年9期)2022-05-16
小哥白尼(野生动物)(2021年9期)2022-01-17
中国林副特产(2020年5期)2020-10-09
快乐作文(1.2年级)(2019年3期)2019-09-10
英语文摘(2019年5期)2019-07-13
幼儿画刊(2018年10期)2018-10-27
现代营销(创富信息版)(2016年6期)2016-11-21
小雪花·成长指南(2016年2期)2016-03-16
现代营销(创富信息版)(2016年12期)2016-02-08
