
亚硝酸盐对聚磷菌厌氧代谢的影响
2012-06-08 11:46:20杨莹莹王向东李博晓彭永臻
土木与环境工程学报 2012年2期
曾 薇,杨莹莹,王向东,李 磊,李博晓,彭永臻
(北京工业大学 环境与能源工程学院,北京100124)
强化生物除磷(Enhanced biological phosphorus removal,EBPR)作为一种经济可持续的除磷工艺在污水处理厂中得到了广泛的应用。EBPR工艺的实现主要依靠聚磷菌(Phosphate accumulation organisms,PAOs)在交替的厌氧/好氧环境中放磷及过量吸磷,然后通过排放剩余污泥达到除磷的目的。厌氧条件下,PAOs体内的多聚磷酸盐(polyphosphate,poly-P)分解,产生的能量用于挥发性脂肪酸(volatile fatty acids,VFAs)的 吸 收,并 以 聚 羟 基 烷 酸 (poly-βhydroxyalkanoates,PHA)的形式储存于细胞内。PHA合成所需的还原力由糖原的分解供给。好氧条件下,PAOs分解体内储存的PHA,获得的能量用于细胞的生长、糖原的补充和磷的过量吸收及储存[1]。
亚硝酸盐作为硝化和反硝化过程的中间产物,广泛存在于生活污水脱氮除磷系统中,并在一定条件下形成积累,含量甚至能够达到19 mg/L以上[2-3]。有报道指出在污水生物处理系统中,高浓度的亚硝酸盐对多种微生物产生抑制作用,包括普通需氧型异养菌和硝化细菌等[4-6]。近年,亚硝酸盐对PAOs好氧吸磷和反硝化吸磷的抑制引起了学者的广泛关注。Zhou等[7]以4种不同的污泥为研究对象,考察了亚硝酸盐对PAOs反硝化吸磷过程的影响,发现亚硝的游离态形式——游离亚硝FNA(Free nitrous acid,FNA)才是真正的抑制剂,当FNA浓度达到0.02 mg HNO2-N/L时,吸磷作用则被完全抑制。当FNA从0.002 mg HNO2-N/L增加到0.02 mg HNO2-N/L时,聚磷菌的反硝化速率减小了约40%。Weon等[8]人以Acinetobacter sp为对象,研究了亚硝酸盐对聚磷菌好氧代谢的影响,指出Acinetobacter sp的好氧生长和吸磷过程都受到了亚硝酸盐的抑制。Pijuan等[9]进一步提出了FNA是真正的抑制剂,对好氧条件下Accumulibacter PAO的生长、糖原的合成以及吸磷都造成严重的抑制。可见,亚硝酸盐的存在对于EBPR系统的稳定运行有着重要的影响。
目前,亚硝酸盐对PAOs代谢影响的研究多集中于反硝化吸磷和好氧吸磷过程,而亚硝酸盐对PAOs厌氧代谢影响的研究较少。本研究以来自不同的EBPR系统的活性污泥为研究对象,在厌氧条件下投加一定量的亚硝酸盐,分析亚硝酸盐对PAOs厌氧代谢的影响机制,有利于进一步了解亚硝酸盐对生物除磷过程的影响。
1 材料与方法
1.1 实验污泥和反应器运行
试验所用污泥取自2个不同的EBPR系统。
活性污泥1:取自实验室SBR 1反应器,该反应器采用厌氧/好氧运行方式富集聚磷菌,进水为人工合成废水。有效容积10 L,每天运行2周期,每周期290 min(厌氧120 min,好氧160 min,沉淀排水10 min),排水比1:4。通过调节流量计维持溶解氧在2.0±0.5 mg/L之间。用1M HCL和1M NaOH调节p H,维持厌氧进水初始p H=7.5,好氧段p H在7.3~7.6之间。
活性污泥2:取自实验室SBR 2反应器,以厌氧/好氧方式运行,进水为实际生活污水。有效容积为7 L,每天运行3周期,每周期420 min(厌氧120 min,好氧240 min,沉淀排水60 min),排水比为1∶2。好氧阶段溶解氧维持在2~3 mg/L之间,好氧末端亚硝酸盐积累率达到95%以上。
1.2 试验水质
SBR1反应器采用人工配水,每升配水所含成分见表1,其中营养液成分(1 L):1.5 g FeCl3·6 H2O,0.15 g H3BO3,0.03 g CuSO4·5H2O,0.18 g KI,0.12 g MnCl2·4 H2O,0.06 g Na2MoO4·2 H2O,0.12 g ZnSO4·7 H2O,0.15 g CoCl2·6 H2O,10 g EDTA。配水的水质为:COD浓度为800 mg/L,磷酸盐浓度为20 mg/L。
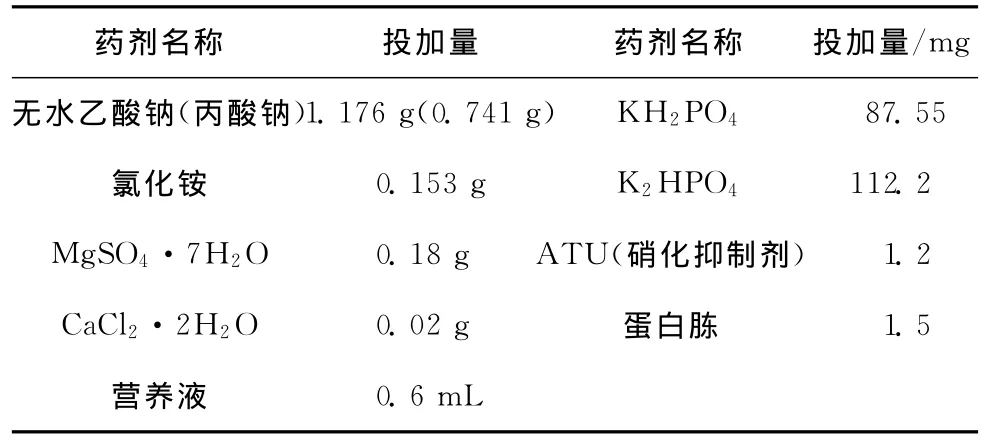
表1 人工配水成份及水质指标
SBR2反应器的实验用水取自某生活小区化粪池的实际生活污水,试验期间平均水质主要参数为:COD=175.7~223.8 mg/L,平均 194.5 mg/L;-N=60.42~79.43 mg/L,平均69.44 mg/L;-P=4.30~8.28 mg/L,平均5.71 mg/L。
1.3 静态试验
从2个SBR反应器好氧末端取4 L污泥,清洗3遍,将清洗后的污泥平均分装于4个反应瓶中,然后加入人工配水至1.5 L。人工配水中不含磷源和碳源,其余成分如表1所示。向4个反应瓶中分别加入10 m L 30 g/L的乙酸钠溶液,使其初始碳源浓度达到200 mg/L。同时投加不等量的亚硝酸钠,使得4个反应瓶中亚硝态氮的浓度分别达到0、10、20和30 mg/L。实验过程中在线监测p H,用0.5 mol/L的HCL和0.5 mol/L NaOH调节并维持混合液的p H=7.5±0.05,温度控制为25℃±0.5℃。
1.4 分析方法
采用荧光原位杂交技术(Fluorescence in-situ hybridization,FISH)分析聚磷菌的富集程度。试验中采用的16S r RNA寡核苷酸探针如表2所示。其中全菌探针EUBmix由EUB338、EUB338Ⅱ和EUB338Ⅲ混合而成;聚磷菌探针PAOmix由PAO462、PAO651和PAO846混合而成。
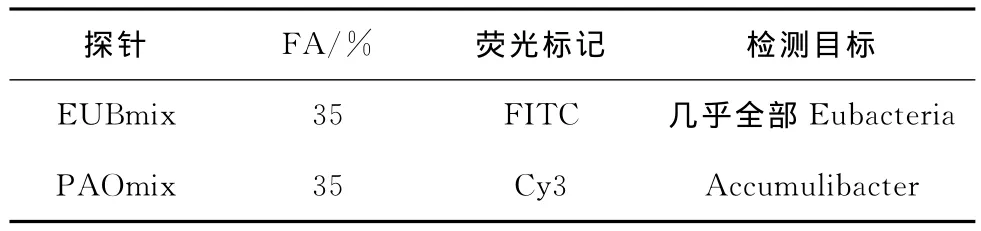
表2 FISH分析中采用的16S rRNA寡核苷酸探针[11-12]
2 实验结果和分析
2.1 亚硝酸盐对人工配水富集聚磷菌厌氧代谢的影响
图1表示不同亚硝酸盐浓度下活性污泥1中VFA、PO3-4—P、PHA和NO-2—N浓度的变化。由图1(a)可看出,与未投加亚硝酸盐的情况相比,投加亚硝酸盐的反应瓶中VFA的消耗速率明显减慢,直至反应进行至120 min反应瓶内仍有VFA剩余。反应进行至70 min时,未投加亚硝酸盐的反应瓶中VFA已经消耗完,而亚硝态氮浓度分别为10、20和30 mg/L的反应瓶中,VFA的浓度分别为73.88、92.93和98.25 mg/L。由图1(c)可看出一部分VFA被聚磷菌吸收以PHA的形式储存在体内,而由图1(d)可知还有一部分VFA被异养菌作为碳源用于亚硝酸型反硝化反应。亚硝态氮浓度分别为10、20、30 mg/L时,亚硝酸盐的利用速率基本一致,即反硝化速率没有明显的差异。由此可以看出,亚硝酸盐对聚磷菌吸收VFA的过程造成抑制,但对于亚硝酸盐的反硝化过程没有明显影响。
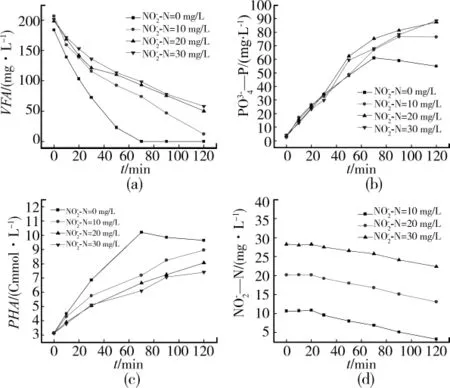
图1 不同亚硝酸盐浓度下活性污泥1中VFA、—P、PHA和—N浓度的变化
图1(b)反映了不同亚硝酸盐浓度下磷酸盐浓度的变化。与VFA的浓度变化不同,向反应瓶中投加亚硝酸盐不但没有减少磷酸盐的释放量,反而促进了聚磷菌对磷酸盐的进一步释放,在反应进行至70 min时,亚硝酸盐浓度为30 mg/L的反应瓶中磷酸盐的释放量达到67.8 mg/L,比未投加亚硝酸盐反应瓶中的浓度高6.9 mg/L。在未投加亚硝酸盐的反应瓶中,反应进行至70 min时,磷酸盐的释放量 出 现 了 下 降 的 现 象,由 60.9 mg/L 降 至54.9 mg/L。观察图1(c)亚硝态氮浓度为0时聚磷菌合成PHA的变化情况,发现70 min时PHA浓度也出现了不增加反而下降的现象,PHA浓度由10.20 Cmmol/L降至9.64 Cmmol/L。以上现象说明反应瓶中的VFA被消耗完之后,聚磷菌以体内储存的PHA为电子供体,以反应瓶中少量溶解氧为电子受体进行吸磷反应。
2.2 亚硝酸盐对短程脱氮除磷活性污泥中聚磷菌厌氧代谢的影响
图2表示不同亚硝酸盐浓度下活性污泥2中VFA、PO3-4—P、PHA和NO-2—N浓度的变化。从图2(a)可以看出,当亚硝态氮浓度分别为0、10、20、30 mg/L时,VFA 分别在120、90、50、50 min消耗完,投加亚硝酸盐的反应瓶中VFA的利用速率大于未投加亚硝酸盐的反应瓶中的VFA利用速率。从图2(c)和图2(d)可知,对于投加亚硝酸盐的反应瓶,VFA被聚磷菌吸收除了作为内碳源被储存于体内之外,还有一部分VFA作为碳源用于亚硝酸型反硝化反应,所以有亚硝酸盐存在的反应瓶中VFA的消耗速率较快。由图2(d)亚硝态氮的变化曲线可知,当亚硝态氮浓度为10、20、30 mg/L时,亚硝态氮的消耗速率没有明显的差异,说明亚硝酸盐没有抑制反硝化反应。

图2 不同亚硝酸盐浓度下活性污泥2中VFA、—P、PHA和—N浓度的变化
图2(b)表示了不同亚硝态氮浓度下聚磷菌释放磷酸盐的浓度变化,从曲线的变化可看出随着亚硝态氮浓度的增大,聚磷菌释放磷酸盐的量减少。另外,可观察到当亚硝态氮为20和30 mg/L时反应进行至50 min出现磷酸盐浓度没有升高反而降低的现象,而图2(c)中相应浓度下的PHA浓度也出现了一致的变化趋势。由图2(d)可看出50 min时,20 mg/L的反应瓶中亚硝态氮全部被反硝化,此时30 mg/L的反应瓶中仍存在5.5 mg/L亚硝态氮。以上现象说明,当反应瓶中碳源消耗完后,聚磷菌能够以PHA为电子供体,以亚硝酸盐为电子受体进行吸磷,这也说明该系统中存在反硝化聚磷菌。
由以上分析可知,30 mg/L的亚硝酸盐对聚磷菌吸收VFA、释放磷酸盐、合成PHA和亚硝酸盐的反硝化过程没有形成明显的抑制,但是亚硝酸盐的存在仍对系统造成一定的影响。厌氧条件下投加亚硝酸盐,系统中发生异养菌的反硝化作用,引起聚磷菌与普通异养反硝化菌之间对于VFA的竞争。而当系统中的碳源不充足时,聚磷菌不能获得足量的VFA,导致PHA的合成和磷酸盐的释放减少,进而影响好氧吸磷作用,若不补充碳源,最终将导致除磷系统的恶化。
2.3 微生物种群分析
图3(a)和图3(b)分别为活性污泥1和活性污泥2中全菌EUBmix和PAOmix合成图。经计算可知,活性污泥1中的聚磷菌占到全菌的55%,活性污泥2中的聚磷菌占到全菌的7.6%。由以上分析可知,对于处理实际生活污水的短程脱氮除磷活性污泥,由于生活污水的水质复杂,进水有机物负荷变化较大,污泥中的微生物种群较丰富且抗冲击负荷能力较强。当系统中有亚硝酸盐存在时,大量的异养菌能够利用碳源快速的将亚硝酸盐还原,从而在一定程度上减弱了亚硝酸盐作为抑制剂对聚磷菌厌氧释磷的抑制作用。而富集聚磷菌系统由于长期用人工配水培养聚磷菌,其污泥中微生物的组成较为简单,对环境变化的适应能力较弱,当系统中加入亚硝酸盐后,聚磷菌的厌氧代谢受到了抑制作用。

图3 2种活性污泥中聚磷菌的FISH合成效果图
2.4 亚硝酸盐抑制PAOs厌氧代谢的机理分析
2.4.1 亚硝酸盐对PAOs吸收 VFA的影响Mino[13]在1987年提出乙酸通过主动运输的方式进入聚磷菌体内,Smolders等[14]的实验结果也支持了此观点。Saunders等[15]在膜运输水平下对厌氧条件下PAOs吸收VFA的机制进行了更深入的研究,发现乙酸通过细胞膜的驱动力主要来自质子驱动力PMF,即次级运输。质子驱动力由Pit载体蛋白排出磷和质子而产生[16]。
主动运输的过程需要消耗能量,关于微生物体内ATP形成,目前最为公认的是Peter D Mitchell在1961年提出化学渗透假说[17],提出生成ATP的氧化与磷酸化之间起到偶联作用的是H+的跨膜梯度。现有研究认为亚硝酸盐是一种解偶联剂,能够提高质子通过穿梭机制透过细胞膜的性能,破坏跨膜的质子梯度[18]。在微生物体内,由于细胞膜具有选择性,H+不能自由的通过细胞膜,于是在细胞膜的两侧形成了质子跨膜梯度,而解偶联剂能够提高细胞膜对H+的通透性,从而破坏了细胞膜两侧的质子梯度,抑制ADP+Pi生成ATP的磷酸化作用。由此而推测出,在富集聚磷菌的反应瓶中投加的亚硝酸盐是解偶联剂,通过破坏质子驱动力,破坏VFA主动运输的过程,导致聚磷菌吸收VFA的量减少。
2.4.2 亚硝酸盐对PAOs释放磷酸盐的影响
poly-P存在于所有的细胞中,可见其对细胞功能有着重要的作用。在EBPR系统中,poly-P的作用还不十分清楚,但是目前被广泛认同的一个重要作用是胞内poly-P的水解能够为VFA的吸收和PHA的合成提供能量[19]。
厌氧条件下P的释放和VFA的吸收的比率P/VFA是考察PAOs性能的一个重要参数[14],表示了PAOs每吸收一个单位的VFA所释放的P的量,而吸收VFA所需的能量主要来自于poly-P的水解,所以说P/VFA的值体现的是吸收一个单位的VFA所需能量的多少。图4表示的是富集聚磷菌系统静态试验中不同亚硝酸盐浓度下P/VFA的变化情况。与未投加亚硝酸盐的情况相比,有亚硝酸盐存在的系统中P/VFA的值较大,说明当有亚硝酸盐存在时需要更多的能量用于VFA的吸收。
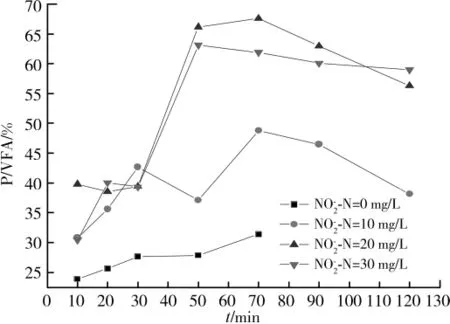
4 富集聚磷菌系统静态试验中P/VFA的变化
在厌氧条件下,有亚硝酸盐的除磷系统中P/VFA的比值较大,说明系统中可能发生了二次释磷现象。所谓二次释磷是指在厌氧条件下,不伴随有机物(如乙酸等)的吸收以及多聚物(如PHB等)的合成的磷释放。而引起二次释磷的原因主要有2个:一是厌氧条件下加入降低PMF的物质;二是细菌的内源呼吸引起了胞内多聚磷酸盐的水解[21]。
Bond等[20]研究表明在碱性条件下,聚磷菌能够通过降解体内储存的poly-P调节EBPR系统中的p H。Smolders等[14]研究认为随着p H值的升高,P/VFA的比值增大,即p H值的升高使得聚磷菌的释磷能力得到了增强。厌氧条件下,聚磷菌吸收VFA的过程需要消耗PMF,而p H的升高会降低PMF,为了维持PMF值的恒定,聚磷菌需要水解更多的poly-P,所以升高的p H使得聚磷菌释放磷酸盐的量增多。由此可推测出,在富集聚磷菌的反应瓶中,厌氧条件下亚硝酸盐的投加促进了磷酸盐的释放,是因为亚硝酸盐的存在破坏了聚磷菌细胞的PMF,为了维持PMF恒定,为聚磷菌吸收VFA提供能量,聚磷菌需要分解更多的poly-P,为厌氧代谢过程提供能量。
3 结 论
1)人工配水富集聚磷菌的活性污泥中,当亚硝态氮的浓度超过10 mg/L时,聚磷菌吸收VFA的过程受到抑制,使得PHA的合成量减少,释磷量增加;在SBR短程脱氮除磷活性污泥中,当亚硝态氮浓度达到30 mg/L时,聚磷菌的厌氧代谢没有受到抑制。
2)人工配水富集聚磷菌的活性污泥,有亚硝酸盐与未投加亚硝酸盐的情况相比,P/VFA明显增大,即有亚硝酸盐存在时需要更多的能量用于VFA的吸收。
3)SBR短程脱氮除磷活性污泥加入亚硝酸盐后发生异养菌的反硝化作用,引起异养反硝化菌与聚磷菌竞争VFA,在碳源不充足的情况下,造成聚磷菌合成PHA和释磷量的减少。
4)厌氧条件下,来自不同工艺的污泥对于亚硝酸盐的耐受能力不同,与活性污泥的微生物组成有关。采用FISH技术对2种活性污泥中的聚磷菌做定量分析,发现人工配水富集聚磷菌活性污泥中聚磷菌含量达到55%,而短程脱氮除磷活性污泥中的聚磷菌为7.6%。处理生活污水的活性污泥菌群比人工配水富集聚磷菌系统中的菌群更加复杂,其对亚硝酸盐的耐受能力更强。
[1]OEHMEN A,LEMOS P C,CARVALHO G,et al.Advances in enhanced biological phosphorus removal:From micro to macro scale[J].Water Research,2007,41:2271-2300.
[2]GUO J H,PENG Y Z,HUANG H J,et al.Short-and long-term effects of temperature on partial nitrification in a sequencing batch reactor treating domestic wastewater[J].J.Hazard.Mater.,2010,179(1-3):471-479.
[3]ZENG W,LI L,YANG Y,et al.Nitritation and denitritation of domestic wastewater using a continuous anaerobic-anoxic-aerobic (A2O)process at ambient temperatures[J].Bioresourch Technology,2010,101:8074-8082.
[4]ANTHONISEN A,LOEHR R,PRAKASAM T,et al.Inhibition of nitrification by ammonia and nitrous acid[J].Water Pollut Control Fed,1976,48:835-852.
[5]MUSVOTO E V,LAKAY M T,CASEY T G,et al.Filamentous organism bulking in nutrient removal activated sludge systems Paper 8:The effect of nitrate and nitrite[J].Water SA,1999,25(4):397-407.
[6]VADIVELU V M,KELLER J,YUAN Z G.Effect of free ammonia and free nitrous acid concentration on the anabolic and catabolic processes of an enriched nitrosomonas culture[J].Biotechnology and Bioengineering,2006,95(5):830-839.
[7]ZHOU Y,PIJUAN M,YUAN Z.Free nitrous acid inhibitionon anoxic phosphorus uptake and denitrification by poly-phosphate accumulating organisms[J].Biotechnology and Bioengineering,2007,98(4):903-912.
[8]WEON S Y,LEE C W,LEE S I,et al.Nitrite inhibition of aerobic growth of Acinetobacter sp.[J].Water Research,2002,36:4471-4476.
[9]PIJUAN M,YE L,YUAN Z G.Free nitrous acid inhibition on the aerobic metabolism of poly-phosphate accumulating organisms[J].Water Research,2010,44(20):6063-6072.
[10]国家环保局.水和废水检测分析方法[M].北京:环境科学出版社,1998.
[11]DAIMS H,BRUHL A,AMANN R,et al.The domain-specific probe EUB338 is insufficient for the detection of all bacteria:Development and evaluation of a more comprehensive probe set[J].Systematic and Applied Microbiology,1999,22(3):434-444.
[12]CROCETTI G R,HUGENHOLTZ P,BOND P L,et al. Identification of polyphosphate-accumulating organisms and design of 16S r RNA-directed probes for their detection and quantitation[J]. Applied and Environmental Microbiology,2000,66(3):1175-1182.
[13]MINO T.Effect of Phosphorus Accumulation on Acetate Metabolism in the Biological Phosphorus Removal Process[M].Biological Phosphate Removal from Wastewaters,Ramadori R,Oxford:Pergamon Press,1987:27-38.
[14]SMOLDERS G J F, VAN DER MEIJ J, VAN LOOSDRECHT M C M,et al.Model of the anaerobic metabolism of the biological phosphorus removal process: stoichiometry and p H influence[J].Biotechnology and Bioengineering,1994,43:461-470.
[15]SAUNDERS A T.The Physiology of Microorganisms in Enhanced Biological Phosphorus Removal[D].St.Lucia:University of Queensland,2005.
[16]MARTIN H G,IVANOVA N,KUNIN V,et al.Metagenomic analysis of two enhanced biological phosphorus removal(EBPR)sludge communities[J].Nature Biotechnol,2006,24(10):1263-1269.
[17]YARBROUGH J,RAKE J,EAGON R.Bacterial inhibitory effects of nitrite-inhibition of activ-transport,but not of group translocation,and of intracellular enzymes[J].Applied and Environmental Microbiology,1980,39(4):831-834.
[18]MITCHELL P D.Coupling of phosphorylation to electron and hydrogen transport by a chemiosmotic type of membrane possible function of chains of catalysts[J].Nature,1961,191:144-148.
[19]HU J Y,ONG S L,NG W J,et al.A new method for characterizing denitrifying phosphorus removal bacteria by using three different types of electron acceptors[J].Water Research,2003,37:3463-3471.
[20]SEVIOUR R J, MINO T, ONUKI M. The microbiology of biological phosphorus removal in activated sludge systems[J]. FEMS Microbiology Reviews,2003,27:99-127.
[21]王景峰,王暄,田淑媛.SBR强化生物系统处理高浓度含磷废水 [J].中国给水排水,2003,19(5):61-63.WANG JING-FENG,WANG XUAN,TIAN SHUYUAN.Phosphorus removal from wastewater using enhanced sequencing batch reactor[J].China Water and Wastewater,2003,19(5):61-63.
[22]BOND P L,KELLER J,BLACKALL L L.Anaerobic phosphate release from activated sludge with enhanced biological phosphorus removal:A possible mechanism of intracellular p H control [J].Biotechnology and Bioengineering,1999,63:507-515.
猜你喜欢
疯狂英语·新读写(2018年3期)2018-11-29 22:37:11
电镀与环保(2016年2期)2017-01-20 08:15:25
微生物学杂志(2016年2期)2016-12-21 07:46:07
环境污染与防治(2016年12期)2016-03-13 05:52:12
四川化工(2015年2期)2015-01-03 11:35:28
湖南大学学报(自然科学版)(2014年7期)2014-12-19 05:26:12
无机化学学报(2014年12期)2014-02-28 17:33:54
食品科学(2013年6期)2013-03-11 18:20:11
植物营养与肥料学报(2011年4期)2011-10-26 02:44:56
植物营养与肥料学报(2010年3期)2010-11-16 00:59:24
