
不同类型牡丹花期蛋白质和脯氨酸的含量变化
2012-06-06 06:57李亚杰施江胥华伟张淑玲霍志鹏王建章
河南科技学院学报(自然科学版) 2012年1期
李亚杰,施江,胥华伟,张淑玲,霍志鹏,王建章
(1.河南科技大学农学院,河南洛阳471000;2.洛阳国际牡丹园,河南洛阳471000)
牡丹Paeonia suffruticosa属芍药科Paeoniaceae芍药属Paeonia,多年生落叶小灌木,又名洛阳花、富贵花,具有很高的观赏价值和栽培价值[1].但是牡丹的自然花期较短,最佳观赏期仅2~3 d,远远不能满足人们旅游观赏的需要,所以探讨牡丹花期的衰老机理和如何延长牡丹花期显得非常重要.牡丹为乙烯敏感型花卉[2],内源激素的改变常常会带来一系列的生理生化变化,蛋白质作为生命活动的体现者,常在植物衰老过程中变化显著,因此被认为是重要的衰老指标之一[3].而脯氨酸作为一种渗透调节物质,一方面可以缓解胁迫带来的渗透压,另一方面还可以清除O2-、OH-等活性氧,维持蛋白质的正常结构,此外,它还可以作为一种生长发育信号,调控植物的生长发育[4].因此本文选用不同花期类型牡丹,探讨其叶和花中可溶性蛋白和脯氨酸含量在开花及衰老过程中的变化规律,希望能够进一步丰富牡丹衰老机理的研究,为牡丹花期调控提供理论依据.
1 材料与方法
1.1 试验材料
供试材料为牡丹早花品种‘凤丹白’、中花品种‘洛阳红’和晚花品种‘玉楼点翠’,于2011年4-5月采自洛阳国际牡丹园.参照王荣花等的标准[5],将花朵发育时期划分为露色期(Ⅰ期)、绽口期(Ⅱ期)、初开期(Ⅲ期)、半开期(Ⅳ期)、盛开期(Ⅴ期)和开始衰败期(Ⅵ期).从牡丹植株上采切不同发育阶段叶、花,用冰盒带回实验室,用于各项生理指标的测定.
1.2 试验方法
1.2.1 可溶性蛋白含量的测定 称取牡丹鲜样0.2~0.5g,分3次总共加入5mL蒸馏水研磨匀浆,然后3000r/min离心10 min,取上清液0.1 mL于试管中,加入5 mL考马斯亮蓝G-250溶液,充分混合;放置5 min后在595 nm下比色,以5 mL考马斯亮蓝G-250溶液为CK,测定其吸光度,并通过以牛血清蛋白所作标准曲线求值,重复3次.根据以下公式计算其可溶性蛋白含量:
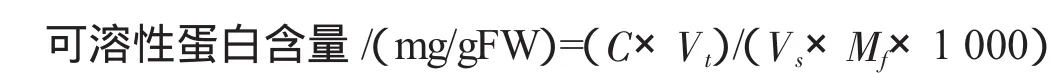
式中:C为查标准曲线值/μg,Vt为提取液总体积/mL,Vs为测定时的加样量/mL,Mf为样品鲜重/g.
1.2.1 脯氨酸含量的测定 将待测叶片擦拭干净,剪碎混匀,分别称取0.5 g,放入试管中,加入体积分数为3%的磺基水杨酸5 mL,在沸水浴中提取10 min,提取过程中要不停地摇动.取出试管冷却以后过滤到干净的试管中,滤液即为脯氨酸的提取液.吸取滤液2 mL于另一干净的带玻璃塞的试管中,并加入2 mL冰醋酸和2 mL体积分数为2.5%的酸性茚三酮,沸水浴中加热30 min,溶液即呈红色.冷却后加入5 mL甲苯,震荡30 s,静置片刻,取上部甲苯萃取物质,以甲苯为空白对照,在分光光度计上520 nm波长下比色,测定其吸光值.测定2 mL待测液中脯氨酸的质量浓度(μg/2 mL).根据以下公式计算脯氨酸含量:
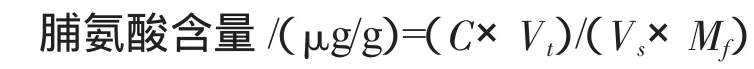
式中:C为2 mL测定液中脯氨酸的质量浓度/(μg/g),Vt为提取液总体积/mL,Vs为测定时的加样量/mL,Mf为样品鲜重/g.
2 结果与分析
2.1 牡丹花期可溶性蛋白含量的变化
牡丹叶片中可溶性蛋白含量测定结果见图1.
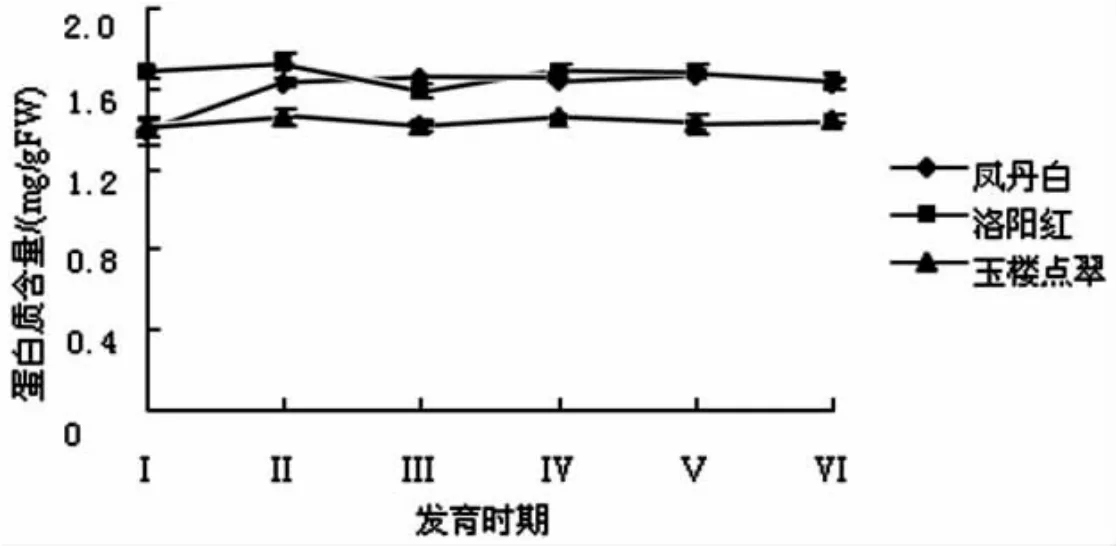
图1 牡丹叶片中可溶性蛋白含量的变化
由图1可知,在开花前期,3种牡丹叶片中可溶性蛋白含量略有上升,总体浮动不大,这是由于叶片自身的生长需要维持较高的蛋白质,以满足其生理功能;在开花后期叶片蛋白质含量波动不大,说明牡丹叶片在花期没有出现衰老的显著特征,这与观察现象一致.
牡丹花瓣中可溶性蛋白含量测定结果见图2.
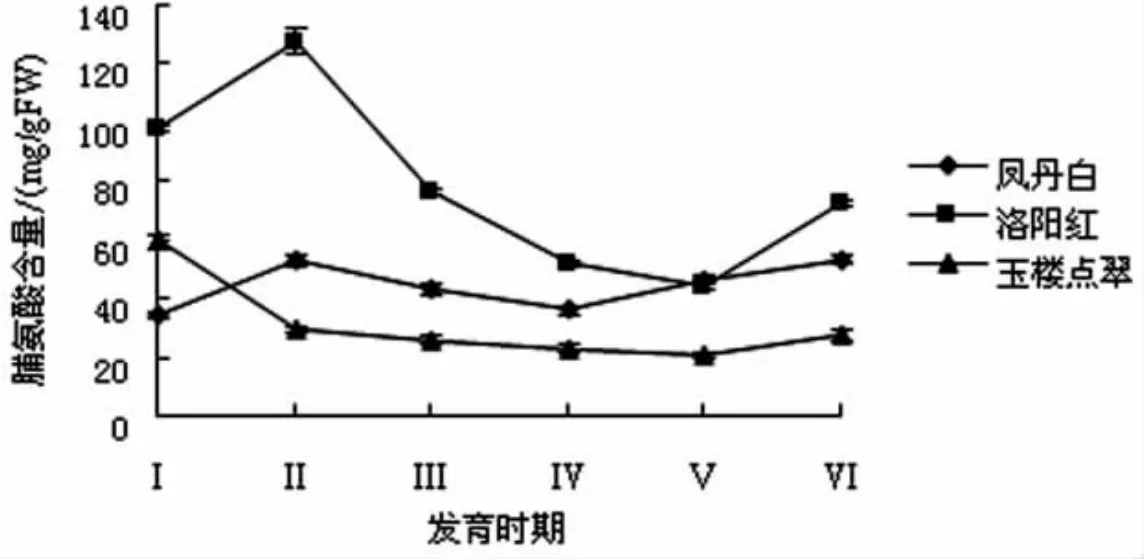
图2 牡丹花瓣中可溶性蛋白含量的变化
由图2可知,‘洛阳红’和‘玉楼点翠’花瓣蛋白质含量略有上升,说明花瓣有蛋白质的合成,满足其强烈的开花需求及细胞增长,而从Ⅱ期后开始逐渐下降,‘凤丹白’花瓣蛋白质含量则随花期持续下降,表明随牡丹花瓣的衰老,有机质大量分解,异化作用加强,花瓣可溶性蛋白含量降低.‘凤丹白’和‘玉楼点翠’末期出现上升趋势,可能是随着花的衰败细胞活性降低,降解蛋白质的能力下降,水分蒸发导致含量上升.
2.2 牡丹花期脯氨酸含量的变化
牡丹叶片中脯氨酸含量的测定结果见图3.
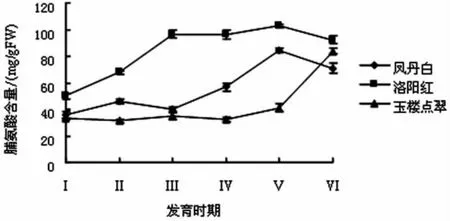
图3 牡丹叶片中脯氨酸含量的变化
由图3可知,3种牡丹叶片在露色期到始衰期的脯氨酸含量不同,变化趋势较为一致,呈逐渐上升状态,‘凤丹白’和‘洛阳红’叶片升高较快,开花末期略有下降,‘玉楼点翠’脯氨酸含量则是前期较为平缓,开花末期急剧上升.分析认为在牡丹开花期,生殖生长较为旺盛,会抑制叶片的营养生长,而且叶片合成的有机物要向代谢旺盛的花瓣转移,导致本身渗透势降低,从而诱导脯氨酸合成系统,提高脯氨酸含量,缓解低渗透势带来的伤害,随着花朵的衰败,生殖生长的抑制逐渐解除,脯氨酸含量出现下降趋势.另外,进入4月下旬以后,温度升高较快,最高达到37℃左右,晚花品种‘玉楼点翠’的叶片很可能受到高温的胁迫,致使脯氨酸含量增加.
牡丹花瓣中脯氨酸含量的测定结果见图4.
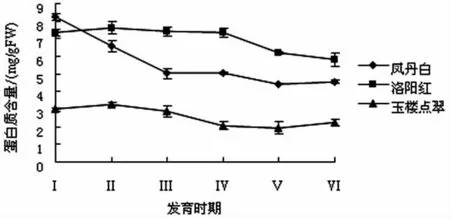
图4 牡丹花瓣中脯氨酸含量的变化
由图4可知,不同花期‘洛阳红’花瓣脯氨酸含量较高,‘凤丹白’次之,‘玉楼点翠’较少,3种牡丹花瓣脯氨酸含量整体呈下降趋势,在开花前期‘凤丹白’和‘洛阳红’花瓣脯氨酸含量略有升高,然后逐渐下降,开花末期不同品种都略有上升.较高的脯氨酸含量可以降低细胞的渗透势,促进细胞吸水和开花生长,另外,脯氨酸可以作为能量物质,为开花提供能量.
3 结论与讨论
在植物的成熟器官中,蛋白质维持一种合成和分解的平衡状态,含量相对稳定,但是随着植物的衰老蛋白质就会降解,细胞结构逐渐破坏,最终走向死亡[6].牡丹为多年生小灌木,叶片和花瓣表现出不同步的衰老现象,牡丹花期一般在四、五月份,从开花到衰败只有一周左右,而叶片生长周期较长,一般在十一月份左右脱落.许多研究认为蛋白质的含量是在植物衰老过程重要指标之一[3],本研究发现在牡丹花期叶片蛋白含量基本稳定,没有出现衰老迹象,这与观察现象相一致,而花瓣蛋白质含量则呈下降趋势,蛋白质逐渐降解会破坏细胞的膜结构,增加膜透性,进一步导致细胞内代谢紊乱,清除O2-等氧自由基的功能受到抑制,膜质过氧化加剧,最终导致了细胞的死亡.
脯氨酸是植物在冰冻、干旱、高温等胁迫条件下累积的主要有机分子,是一种重要的渗透调节物质.在植物的光合器官和生殖器官中,往往有较多的脯氨酸积累,既有助于降低渗透压,提高细胞持水能力,还可以参与氧自由基的清除.此外,1分子脯氨酸氧化可以提供32个ATP,可以作为能量底物,脯氨酸有助于维持蛋白质结构的稳定,这可能是植物对胁迫的生态适应[7].本试验结果表明,叶片脯氨酸含量随花期呈上升趋势,而花瓣脯氨酸含量呈下降趋势,后期略有上升.分析认为,在牡丹花期,花瓣的剧烈活动抑制叶片的营养生长,而叶片营养物质向花瓣的运输进一步增长,诱导了脯氨酸的合成,开花后期随着花瓣的衰败生殖抑制逐渐解除,脯氨酸含量下降,晚花品种‘玉楼点翠’在开花后期叶片脯氨酸快速升高可能是受较高气温的影响[8];花瓣在前几个时期有较高的脯氨酸含量,说明脯氨酸促进了牡丹花瓣的吸水生长,在清除系统中发挥重要作用,减轻活性氧对细胞的伤害,并可能参与蛋白质的合成以及剧烈的呼吸作用,在线粒体中氧化提供能量并逐渐消耗,脯氨酸含量随花期开始下降,半开期和盛花期脯氨酸含量较少,对细胞的持水作用减弱,活性氧积累,并加剧了蛋白质的降解,推进细胞结构的破坏,液泡当中的脯氨酸释放加剧,而且蛋白质的降解一定程度上增加了开花后期脯氨酸的含量.研究认为外源脯氨酸可以提高植物的抗逆性,改变开花时间和延长花期[9],是否对牡丹有效有待进一步研究.
蛋白质的合成和脯氨酸的积累被认为是两个相对独立的过程,并不存在直接关系[10].本试验发现叶片和花瓣中蛋白含量和脯氨酸含量没有显著相关性,与上述结论相一致.另外,在牡丹开花和衰老的过程中,不同时期叶片和花瓣中蛋白质和脯氨酸含量发生明显的变化.这种变化与品种类型、源-库关系、以及花瓣的程序性死亡、乙烯等激素的调控等相关联,并且与牡丹花的衰老相对应,因此蛋白质和脯氨酸含量变化可能在牡丹花衰老过程中起重要作用.
[1]王莲英.中国牡丹品种图志[M].北京:中国林业出版社,1997:28-45.
[2]史国安,郭香凤,李春丽,等.牡丹花枝不同发育时期各器官乙烯释放和ACC含量的变化[J].园艺学报,2010,37(1):77-82.
[3]高勇,吴绍锦.月季切花瓶插期生理变化与衰老关系的研究[J].园艺学报,1990,17(1):71-75.
[4]谢虹,杨兰,李忠光.脯氨酸在植物非生物胁迫耐性形成中的作用[J].生物技术通报,2011(2):23-27.
[5]王荣花,刘雅莉,李嘉瑞.不同发育阶段牡丹和芍药切花开花生理特性的研究[J].园艺学报,2005,32(5):861-865.
[6]杨淑慎,高俊风,李学俊.高等植物叶片的衰老[J].西北植物学报,2001,21(6):1271-1277.
[7]焦蓉,刘好宝,刘贯山,等.论脯氨酸累积与植物抗渗透胁迫[J].中国农学通报,2011,27(7):216-221.
[8]骆俊,韩金蓉,王艳,等.高温胁迫下牡丹的抗逆生理响应[J].长江大学学报:自然科学版,2011,8(2):223-226.
[9]Lehmann S,Funk D,Szabados L,et a1.Prnline metabolismandtransport in plant devement[J].AminoAcids,2010,39(4):949-962.
[10]班青宇,耿建峰,侯喜林,等.不结球白菜叶片脯氨酸与可溶性蛋白含量的QTL分析[J].南京农业大学学报,2010,33(2):375-381.
猜你喜欢
特产研究(2022年6期)2023-01-17
中老年保健(2022年3期)2022-11-21
数学小灵通(1-2年级)(2021年9期)2021-10-12
学生天地(2020年2期)2020-08-25
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
小天使·五年级语数英综合(2018年6期)2018-06-13
现代园艺(2017年13期)2018-01-19
连环画报(2017年1期)2017-07-13
都市(2017年6期)2017-07-03
