
不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼生长性能、体组成以及血清生化指标的影响
2012-06-06 09:47赛陈3张健东3纪多亮吴灶和3周
动物营养学报 2012年1期
王 赛陈 刚,2,3张健东,2,3纪多亮吴灶和,2,3周 晖,2,3*
(1.广东海洋大学水产学院,湛江 524025;2.广东省水产经济动物病原生物学及流行病学重点实验室,湛江 524025;3.广东省普通高等学校南海水产经济动物增养殖重点实验室,湛江 524025)
饲料中的蛋白质主要来源于动、植物性蛋白质原料,由于鱼粉具有蛋白质含量高、必需氨基酸全面、易被水产动物消化吸收等优点,其一直是水产动物首选的优质蛋白质源[1]。但是,随着水产养殖业的迅速发展和渔获量的降低,鱼粉价格不断攀升,导致水产饲料价格居高不下。因此,寻找鱼粉的替代品成为水产动物营养研究的热点。现有的研究多是利用来源广泛、价格低廉的植物性蛋白质源来替代鱼粉,虽然植物性蛋白质源具有适口性差、氨基酸不平衡、存在抗营养因子等缺点,但通过适当的加工处理和科学的设计配方,可以成为鱼粉的优良替代品[2]。
褐点石斑鱼(Epinephelus fuscoguttatus),又称棕点石斑鱼,俗称老虎斑、虎头斑,在分类学上隶属鲈形目,鲈亚目,科,石斑鱼亚科,石斑鱼亚属,为暖水性岛礁性鱼类,在我国主要分布于东海和南海,资源量稀少。由于其肉味鲜美、生长快、市场潜力大,养殖前景看好,已成为福建、广东、海南等地海水养殖的重要名优品种之一。豆粕、玉米蛋白粉、啤酒酵母是动物饲料中常用的蛋白质饲料。已有研究发现,石斑鱼(Epinephelus coioides)[3]、军曹鱼(Rachycentron canadum)[4]、哲罗鱼(Hucho taimen)[5]、牙 鲆(Paralichthys olivaceus)[6]、金鲳鱼(Trachinotus ovatus)[7]饲料中大豆蛋白替代鱼粉蛋白的比例以不超过25%为宜。关于海水鱼饲料中玉米蛋白粉和啤酒酵母替代鱼粉的 研 究 目 前 较 少,对 部 分 虾 类[8-10]、淡 水 鱼类[11-12]和中华鳖(Pelodiscus sinensis)[13]的研究发现,饲料中玉米蛋白粉和啤酒酵母替代鱼粉比例不宜超过10%。为找到可部分替代鱼粉且适合褐点石斑鱼生长的蛋白质源,本试验拟分别采用豆粕、啤酒酵母、玉米蛋白粉部分替代饲料中的鱼粉,研究这3种蛋白质源部分替代鱼粉后对褐点石斑鱼幼鱼生长性能、体组成以及血清生化指标的影响,为今后褐点石斑鱼人工复合饲料的研发提供理论依据。
1 材料与方法
1.1 试验饲料
试验设计5种等氮等能的饲料,对照组饲料(D1)以鱼粉为唯一蛋白质源,试验组饲料(D2~D5)是在对照组饲料的基础上分别用豆粕替代10%的鱼粉、豆粕替代20%的鱼粉、啤酒酵母替代10%的鱼粉、玉米蛋白粉替代10%的鱼粉。试验饲料中添加0.5%的三氧化二铬(Cr2O3)作为营养物质表观消化率测定的指示剂,其组成及营养水平见表1。饲料原料均购自粤海饲料集团有限公司,经粉碎后过60目筛,再经制粒机加工成直径为3.0 mm的颗粒料,风干后置于4℃冰箱中备用。
1.2 试验动物与饲养管理
试验用褐点石斑鱼幼鱼由海南石斑鱼苗场提供。褐点石斑鱼幼鱼从育苗池运到实验室水槽后,采用对照组饲料进行驯化,驯化时间为10 d,在幼鱼完全适应实验室养殖环境,摄食活跃,且没有意外死亡的情况下进入正式试验。选取体重在15 g左右的褐点石斑鱼幼鱼180尾,随机分为5组,每组3个重复,每个重复12尾。试验鱼以重复为单位饲养于半自动充气循环水族箱(60 cm×52 cm×60 cm)中,每个水族箱装水150 L,24 h充气,每天09:00和15:00定时投喂饲料至饱食状态,投喂结束后捞出剩料。试验用水为经曝气、沉淀、沙滤及网滤的天然海水,盐度27‰~32‰。试验期间每天测1次水温,每周测1次水体溶氧和氨氮,水族箱内水温保持在(28±1)℃,氨氮低于0.5 mg/L,溶氧高于5 mg/L,pH 6.7 左右,自然光照,8周后结束试验。
1.3 测定指标和方法
分别在试验开始和结束时对各重复鱼进行计数和称重,并记录投料量和剩料量。试验结束后将鱼禁食1 d,从每组中随机选取12尾鱼(每个重复中随机选取4尾鱼),其中6尾烘干后用于测定全鱼组成,另6尾用于测定体重、肝脏重、肌肉组成以及血清生化指标。
试验饲料和鱼体营养成分的测定采用AOAC(1995)[14]的方法,即干物质含量采用105℃恒温干燥法测定;粗蛋白质含量采用凯氏定氮法测定,粗脂肪含量采用索氏抽提法测定,灰分含量采用高温(550℃)灼烧法测定。饲料和粪样中Cr2O3的测定参照Bolin等[15]的方法。
采用2 mL一次性无菌注射器从尾鳍后方的尾静脉采血,将采集的血液注入玻璃采血管中,于4℃条件下静置,待血液分层后,置于离心机中以3 000 r/min离心10 min,用微量移液枪将血清转入1.5 mL离心管中,用日立全自动生化分析仪进行血清生化指标的测定。
1.4 计算公式




表1 试验饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets(DM basis) %
1.5 数据统计分析
试验结果以平均值±标准差表示。采用SPSS 16.0对数据进行统计学分析,先对数据进行单因素方差分析(One-way ANOVA),再进行Duncan氏多重比较,以检验组间差异显著性,P<0.05表示差异显著。
2 结果与分析
2.1 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼生长性能的影响
由表2可知,D2、D3组的增重率、摄食率、特定生长率和粗蛋白质表观消化率与对照组差异不显著(P>0.05),而 D4、D5组的上述指标均显著低于对照组(P<0.05)。此外,D5组的增重率和粗蛋白质表观消化率还显著低于D2组(P<0.05);D4、D5组的特定生长率还显著低于 D2、D3组(P<0.05);D5组的摄食率还显著低于D2、D3、D4组(P <0.05)。D4、D5组的干物质表观消化率显著高于对照组和 D2组(P<0.05),与 D3组差异不显著(P>0.05)。试验鱼的饲料系数和肝体指数均以对照组最低,D5组最高。其中,饲料系数除D3、D4组间差异不显著(P>0.05)外,其余各组间均差异显著(P<0.05);而肝体指数除D2、D3组间差异不显著(P>0.05)外,其余各组间均差异显著(P<0.05)。各组试验鱼的成活率差异不显著(P>0.05)。
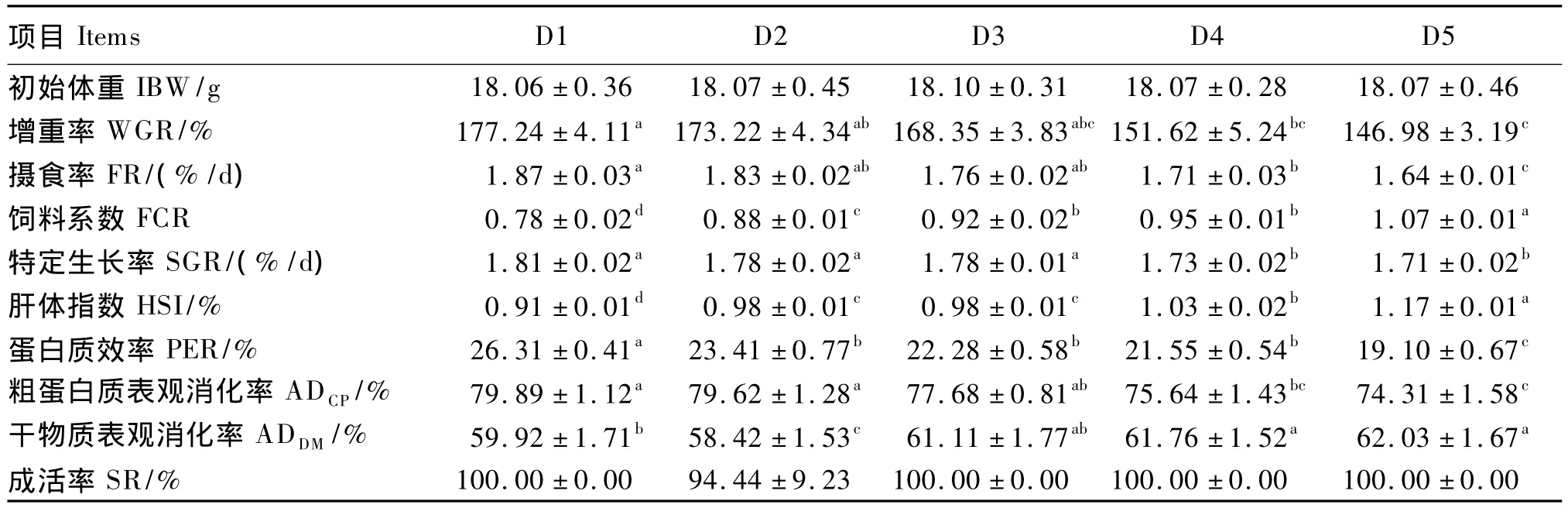
表2 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼生长性能的影响Table 2 Influence of partial replacement of fish meal by different protein sources on growth performance of juvenile Epinephelus fuscoguttatus
2.2 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼体组成的影响
由表3可知,全鱼组成中,D1、D2、D3组粗蛋白质含量显著高于D5组(P<0.05),与D4组无显著差异(P>0.05);D1组粗脂肪含量显著高于D5组(P <0.05),与 D2、D3、D4组无显著差异(P>0.05);水分含量在各组间均无显著差异(P >0.05)。肌肉组成中,D1、D2、D3 组粗蛋白质含量显著高于D5组(P<0.05),与D4组无显著差异(P>0.05);D1组粗脂肪含量显著高于D3、D4组(P<0.05),与 D2、D5组无显著差异(P >0.05);D1组水分含量显著高于 D3组(P<0.05),与 D2、D4、D5 组无显著差异(P >0.05)。

表3 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼体组成的影响(湿重基础)Table 3 Influence of partial replacement of fish meal by different protein sources on body composition of juvenile Epinephelus fuscoguttatus(wet weight basis) %
2.3 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼血清生化指标的影响
由表4可知,D1、D4组血清总蛋白含量显著高于D3组(P<0.05),与 D2、D5组无显著差异(P>0.05);D1组血清球蛋白含量显著高于D3组(P <0.05),与 D2、D4、D5 组无显著差异(P >0.05);D3组血清白球比显著高于其他各组(P<0.05);D1组血清葡萄糖含量显著高于 D3组(P<0.05),与 D2、D4、D5 组无显著差异(P >0.05);D1、D4组血清甘油三酯含量显著高于D3、D5组(P<0.05),与 D2组无显著差异(P>0.05)。各组试验鱼的血清白蛋白、总胆固醇含量以及谷丙转氨酶、谷草转氨酶活性均差异不显著(P >0.05)。
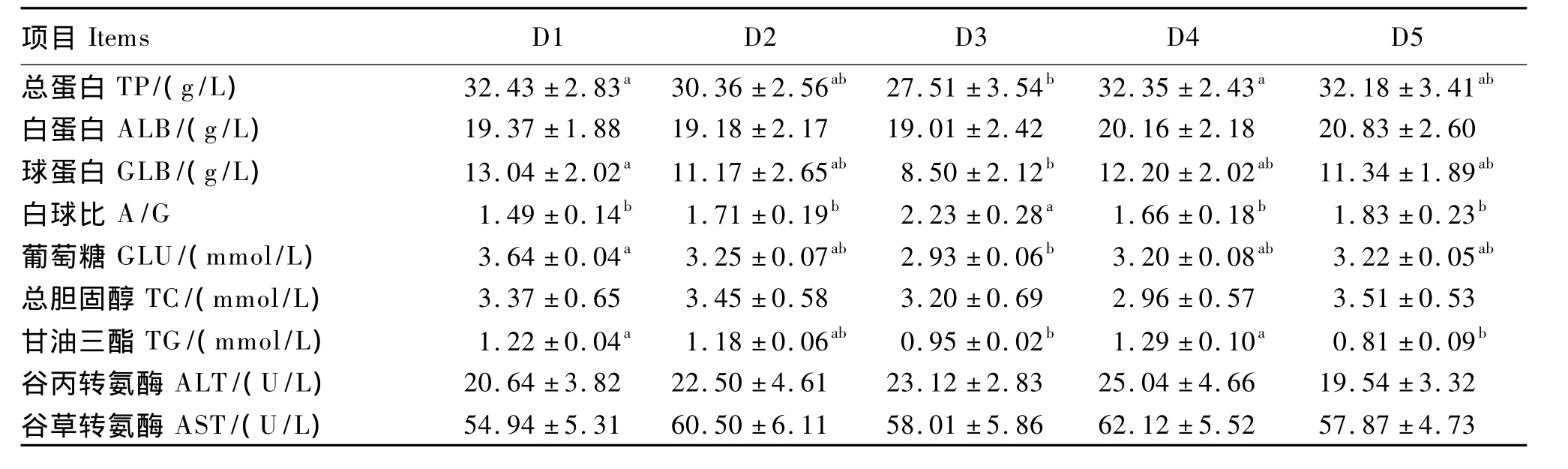
表4 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼血清生化指标的影响Table 4 Influence of partial replacement of fish meal by different protein sources on serum biochemical indices of juvenile Epinephelus fuscoguttatus
3 讨论
3.1 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼生长性能的影响
目前,对单种蛋白质源替代鱼粉的研究较多,而对不同蛋白质源之间替代效果对比的研究较少。水产动物配合饲料中适宜蛋白质源替代鱼粉的比例与养殖动物的种类、食性等多种因素有关,肉食性鱼类对蛋白质的需求量比草食性和杂食性鱼类要高。罗智等[3]研究表明,以发酵豆粕替代10%的鱼粉不影响石斑鱼幼鱼的生长;高荣兵等[16]研究发现,在具有杂食偏植食性特点的点篮子鱼(Siganus guttatus)饲料中,豆粕替代鱼粉比例不宜超过1/3;Chou等[4]研究得出,采用大豆蛋白替代40%的鱼粉蛋白不影响军曹鱼幼鱼的生长;Kaushik 等[17]认为,虹鳟(Oncorhynchus mykiss)饲料中豆粕替代鱼粉比例不宜超过25%;Shimeno等[18]研究发现,黄狮鱼饲料中豆粕替代鱼粉比例不宜超过20%。本试验中,D2、D3组试验鱼的饲料系数和蛋白质效率显著低于对照组,这可能是因为豆粕中存在的抗营养因子和氨基酸不平衡性影响了试验鱼对饲料中营养成分的吸收[1,4,6];但增重率、特定生长率、肝体指数与对照组相比无显著差异,这说明饲料中以豆粕替代20%的鱼粉并不影响褐点石斑鱼幼鱼的正常生长。一般认为,鱼类配合饲料中采用植物性蛋白质源替代鱼粉会降低饲料的适口性,本试验中的3种蛋白质源替代鱼粉后均降低了试验鱼的摄食率,因此应在进一步的试验中添加更适合的诱食剂。在淡水鱼中,陈然等[12]对异育银鲫(Carassius auratus gibelio)的研究发现,当饲料中玉米蛋白粉替代鱼粉比例达到16%时,试验鱼的粗蛋白质表观消化率、干物质表观消化率和增重率显著降低;李自金等[19]对草鱼(Ctenopharyngodon idellua)的研究发现,用啤酒酵母替代饲料中3%的鱼粉有助于草鱼的生长。在海水鱼中,Shiau等[20]对点带石斑鱼(Epinephelus malabaricus)的研究发现,随着饲料中玉米淀粉含量的增加,点带石斑鱼的粗蛋白质表观消化率和增重率降低,体组成中的粗脂肪含量升高;Laining 等[21]认为,老鼠斑(Cromileptes altivelis)对饲料中富含碳水化合物的植物成分难以消化,导致粗蛋白质表观消化率、干物质表观消化率降低。本试验中,D4、D5组试验鱼的生长性能显著低于对照组,这说明饲料中以玉米蛋白粉和啤酒酵母替代10%的鱼粉不适合褐点石斑鱼幼鱼的生长,其原因在于:与鱼粉和豆粕相比,玉米蛋白粉中赖氨酸、蛋氨酸、色氨酸等必需氨基酸缺乏[12,22];此外,玉米蛋白粉含有大量的多糖,而褐点石斑鱼对植物添加成分中的多糖难以消化,这与 Shiau 等[20]、Laining 等[21]、Chen 等[23]等对其他品种石斑鱼的研究结果相似。适量的啤酒酵母虽因含有益生素可通过改善肠道健康提高饲料消化率,但过高的用量(如本试验中9.25%的用量)则可能因为含有过量的核酸等非蛋白氮而影响营养物质的吸收。因此,本研究认为,玉米蛋白粉和啤酒酵母替代10%的鱼粉不能满足褐点石斑鱼幼鱼的营养需求,进一步的试验需要考虑降低玉米蛋白粉和啤酒酵母对鱼粉的替代比例,或者尝试以其他蛋白质源替代鱼粉。
3.2 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼体组成的影响
对军曹鱼[4]、虹鳟[24]的研究发现,大豆蛋白含量的增加未对鱼体体组成产生负面影响;韩斌等[8]研究发现,玉米蛋白粉替代鱼粉未对凡纳滨对虾(Litopenaeus vannamei)体组成产生显著影响;而Deng等[25]对牙鲆幼鱼的研究发现,大豆蛋白替代鱼粉比例的增加使鱼体粗蛋白质含量显著降低。本试验发现,饲料中以玉米蛋白粉替代10%的鱼粉时,褐点石斑鱼幼鱼全鱼粗蛋白质和粗脂肪含量显著降低;饲料中以豆粕替代20%的鱼粉时,褐点石斑鱼幼鱼肌肉粗脂肪和水分含量显著降低。上述结果说明,未经过高温膨化的大豆蛋白中存在胰蛋白酶抑制因子、大豆凝集素等抗营养因子,限制了鱼体对饲料中营养物质的吸收,导致了肌肉中粗脂肪含量的降低,这与哲罗鱼[5]、牙鲆[6]、点篮子鱼[16]的研究结果相似;而玉米蛋白粉中虽然抗营养因子较少,但其碳水化合物含量较多且氨基酸平衡性差,导致试验鱼全鱼粗蛋白质和粗脂肪含量都显著降低,这与试验鱼的生长性能所反映的情况相一致。
3.3 不同蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼血清生化指标的影响
研究表明,哲罗鱼血浆中总蛋白含量随大豆分离蛋白替代鱼粉比例的增加呈降低趋势[5],而牙鲆肝脏中总蛋白含量随豆粕替代鱼粉比例的增加呈升高趋势[6]。本试验中,在以豆粕替代20%的鱼粉时,显著降低了试验鱼血清总蛋白和球蛋白含量,这是因为豆粕中的蛋氨酸、赖氨酸含量均低于鱼粉,且含有某些抗营养因子,高豆粕含量饲料导致褐点石斑鱼幼鱼对饲料中蛋白质的吸收能力降低[1,5-6,24]。研究发现,随着饲料中豆粕含量的增加,牙鲆[6]、虹鳟[24]和异育银鲫[26]血清葡萄糖含量未产生显著变化。Wilson[27]认为,饲料中复杂的碳水化合物结构会降低鱼体对营养物质的利用。本试验中,以豆粕替代20%鱼粉的组褐点石斑鱼幼鱼血清葡萄糖含量显著低于对照组,结合试验鱼的生长情况可知,褐点石斑鱼幼鱼不易消化吸收豆粕中的碳水化合物成分。研究发现,牙鲆[6]、点带石斑鱼[20]、异育银鲫[28]血清胆固醇含量随豆粕替代鱼粉比例的增加呈降低趋势,而瘤棘鲆(Psetta maxima)血清胆固醇和甘油三酯含量随玉米蛋白粉替代鱼粉比例的增加而显著降低[29]。本试验中,3种蛋白质源部分替代鱼粉后对褐点石斑鱼幼鱼血清总胆固醇含量的影响并不显著,结合血清谷草转氨酶、谷丙转氨酶活性的无显著变化可以认为,本试验中的3种蛋白质源部分替代鱼粉对褐点石斑鱼幼鱼血脂的合成与分解代谢以及肝功能和肾功能未产生负面影响。
4 结论
本试验条件下:
①玉米蛋白粉和啤酒酵母替代10%的鱼粉显著降低了褐点石斑鱼幼鱼的生长性能。
②豆粕替代10%和20%的鱼粉虽然降低了褐点石斑鱼幼鱼的蛋白质效率,提高了饲料系数,但对其增重率、摄食率、特定生长率和粗蛋白质表观消化率并未产生显著影响。
③豆粕替代20%的鱼粉降低了褐点石斑鱼幼鱼血清中蛋白质和葡萄糖含量。
④从主要生长指标不受影响的角度考虑,褐点石斑鱼幼鱼饲料中可以用豆粕替代20%的鱼粉。
[1]周岐存,麦康森,刘永坚,等.动植物蛋白源替代鱼粉研究进展[J].水产学报,2005,29(3):404 -410.
[2]LISA D,SARA G,CARL F,et al.Feeding aquaculture growth through globalization:exploitation of marine ecosystems for fish meal[J].Global Environmental Change,2007,17:238-249.
[3]罗智,刘永坚,麦康森,等.石斑鱼配合饲料中发酵豆粕和豆粕部分替代白鱼粉的研究[J].水产学报,2004,28(2):175 -181.
[4]CHOU R L,HER B Y,SU M S,et al.Substituting fish meal with soybean meal in diets of juvenile cobia Rachycentron canadum [J]. Aquaculture,2004,229:325-333.
[5]徐奇友,王常安,许红,等.大豆分离蛋白替代鱼粉对哲罗鱼稚鱼生长、体成分和血液生化指标的影响[J].水生生物学报,2008,32(6):941 -946.
[6]刘襄河,叶继丹,王子甲,等.饲料中豆粕替代鱼粉比例对牙鲆生长性能及生化指标的影响[J].水产学报,2010,34(3):450 -458.
[7]赵丽梅,王喜波,张海涛,等.金鲳鱼饲料中发酵豆粕替代鱼粉的研究[J].中国饲料,2011(11):20-22.
[8]韩斌,华雪铭,周洪琪,等.玉米蛋白粉替代部分鱼粉对凡纳滨对虾摄食量、生长和肌肉成分的影响[J].水产学报,2009,33(4):658 -665.
[9]胡盼,黄旭雄,郭腾飞,等.玉米蛋白粉部分替代鱼粉对日本沼虾生长和肌肉组成的影响[J].上海海洋大学学报,2011,20(2):230 -237.
[10]程媛媛,周洪琪,华雪铭,等.玉米蛋白粉部分替代鱼粉对罗氏沼虾生长、氨基酸沉积率和肌肉营养成分的影响[J].中国水产科学,2009,16(4):572-579.
[11]张梁,周维禄,杨世平,等.饲料酵母替代部分鱼粉养殖淡水白鲳试验[J].郑州牧业工程高等专科学校学报,2003,23(1):4 -5.
[12]陈然,华雪铭,黄旭雄,等.玉米蛋白粉替代鱼粉对异育银鲫生长、蛋白酶活性及表观消化率的影响[J].上海交通大学学报:农业科学版,2009,27(4):358-367.
[13]周贵谭.啤酒酵母替代部分鱼粉对中华鳖生长的试验[J].广东饲料,2003,12(3):11 -12.
[14]AOAC.Official methods of analysis of AOAC international[S].16th ed.Arlington,V.A.:AOAC International,1995.
[15]BOLIN D W,KING R P,KLOSTERMAN E W.A simplified method for the determination of chromicoxide Cr2O3when used as an index substance[J].Science,1952,116:634-635.
[16]高荣兵,庄平,章龙珍,等.豆粕替代鱼粉对点篮子鱼生长性能的影响[J].水产学报,2010,34(10):1534-1540.
[17]KAUSHIK S J,CRAVEDI J P,LALLES J P,et al.Partial or total replacement of fish meal by soybean protein on growth,protein utilization,potential estrogenic or antigenic effects,cholesterolemia and flesh quality in rainbow trout,Oncorhynchus mykiss[J].Aquaculture,1995,133:257-274.
[18]SHIMENO S,KUMON M,ANDO H,et al.The growth performance and body composition of young yellowtail fed with diets containing defatted soybean meals for a long period[J].Nippon Suisan Gakkaishi,1993,59:821 -825.
[19]李自金,邱波,童运彬,等.啤酒酵母取代鱼粉在草鱼上的应用[J].饲料工业,2009,30(20):25 -26.
[20]SHIAU S Y,LAN C W.Optimum dietary protein level and protein to energy ratio for growth of grouper(Epinephelus malabaricus)[J].Aquaculture,1996,145:259-266.
[21]LAINING A,AHMAD R T,WILLIAMS K,et al.Apparent digestibility of selected feed ingredients for humpback grouper,Cromileptes altivelis[J].Aquaculture,2003,218:529-538.
[22]王成章,王恬.饲料学[M].北京:中国农业出版社,2003:215-217.
[23]CHEN H Y,TSAI J C.Optimal dietary protein level for the growth of juvenile grouper,Epinephelus malabaricus,fed semipurified diets[J].Aquaculture,1994,199:265-271.
[24]陆阳,杨雨虹,王裕玉,等.不同比例膨化豆粕替代鱼粉对虹鳟生长、体成分及血液学指标的影响[J].动物营养学报,2010,22(1):221 -227.
[25]DENG J M,MAI K S,AI Q H,et al.Effects of replacing fish meal with soy protein concentrate on feed intake and growth of juvenile Japanese flounder,Paralichthys olivaceus[J].Aquaculture,2006,258:503-513.
[26]王文娟,叶元土,蔡春芳,等.豆粕及其抗营养因子对异育银鲫血清生化和非特异性免疫指标的影响[J].中国饲料,2010(18):30 -33.
[27]WILSON R P.Utilization of dietary carbohydrate by fish[J].Aquaculture,1994,124:67 -80.
[28]王崇,雷武,解绶启,等.饲料中豆粕替代鱼粉蛋白对异育银鲫生长、代谢及免疫功能的影响[J].水生生物学报,2009,33(4):740 -747.
[29]REGOST C,ARZEL J,KAUSHIK S J.Partial or total replacement of fish meal by corn gluten meal in diet for turbot(Psetta maxima)[J].Aquaculture,1999,180:99-117.
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2022年1期)2022-04-26
当代水产(2020年3期)2020-06-15
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
当代水产(2019年5期)2019-07-25
当代水产(2019年1期)2019-05-16
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
