
3个油茶新品种的光合特性
2012-05-30 03:29谭晓风姜志娜杨巧丽
浙江农林大学学报 2012年4期
袁 军,谭晓风,姜志娜,杨巧丽,李 泽
(1.北京林业大学 研究生院,北京 100083;中南林业科技大学 经济林育种与栽培国家林业局重点实验室,湖南 长沙 410004;)
油茶Camellia oleifera是中国南方重要的木本食用油料树种[1],栽培面积占木本油料树种栽培面积的80%以上,已有2300多年的栽培利用历史。目前,中国共有油茶林3.02×106hm2,但长期以来,油茶籽产量低、品质差,制约了油茶产业的可持续发展。因此,选育优良品种,并加以快速繁育与推广,是油茶产业中的一项基础性工作和亟待解决的课题。目前,高光效育种已经成为育种工作者关注的主要问题。不同品种之间由于遗传因子的影响,光合潜能差异很大[2]。研究油茶不同品种间的光合生理特性不仅能为选育高光效品种提供理论依据,还能为良种的栽培和推广提供参考。国内外对油茶光合作用的研究曾有报道[3-4],但主要集中在光合速率及其影响因素等方面,对新培育的高产品种之间光合特性的差异和需光特点的研究尚少。为此,本研究通过对2009年国家新审定的3个油茶品种(‘华硕’‘华金’‘华鑫’)的光合生理特性和叶绿素质量分数等生理生化指标的测定和对比分析,揭示3个品种的基本光合生理特征和规律,为其作为高光效育种材料和丰产栽培提供参考资料。
1 材料与方法
1.1 试验材料
试验材料选自中南林业科技大学在长沙市望城区东城镇新建的高接换冠油茶示范林,以2009年国家审定的3个油茶新品种‘华硕’‘华金’‘华鑫’1年生的换冠林为试材。选择生长良好,长势基本一致的健康植株,每个品种各3株,每株选择南向中等大小、颜色正常的叶片(顶端第5~8片叶)3片,采用美国LI-COR公司生产的LI-6400便携式光合测定系统进行叶片瞬时光合速率测定。
1.2 测定方法
光合速率日变化测定。光合速率日变化于2011年7月上旬进行,天气晴朗,测定时间从7:00-19:00,隔2 h测定1次油茶的净光合速率(Pn),气孔导度(Gs)和胞间二氧化碳体积分数(Ci),同时测定光合有效辐射(PAR),大气温度(T)和相对湿度(HR)。测定前先测自然光照强度,然后将红蓝光源设定为与自然光相同的光强进行测定,当系统稳定后进行记录,数据记录5次·叶片-1,重复测定3次的平均值为光合速率的日变化。
光响应曲线的测定。光响应曲线于2011年7月上旬进行,测定时间为8:30-10:30,天气晴朗,利用LI-6400自动light-curve曲线测定功能,将红蓝光源设定一系列光合光通量密度,考虑到植物对弱光到强光的不敏感性,其梯度设定为:2500,2200,1900,1600,1300,1000,700,400,200,100,80,60,40, 20, 0μmol·m-2·s-1。 其测量参数包括: 净光合速率(Pn),气孔导度(Gs), 胞间二氧化碳体积分数(Ci),光合有效辐射(PAR),大气温度(T)和相对湿度(HR)等生理生态因子。利用非直角双曲线拟合[5],根据拟合曲线计算光饱和点和光补偿点。非直角双曲线模型理论公式为:

其中:Pn为净光合速率,Pm为最大净光合速率,φ为表观量子效率,PAR为光合有效辐射,k为光响应曲线曲角,Rday为暗呼吸速率。
叶绿素质量分数的测定。测定完以上2个指标后,采摘标记的叶片,立刻带回实验室测定,测定方法为丙酮提取法[6]。叶绿素浸提液用721-PC分光光度计测定光密度值,并按Arnon公式计算叶绿素质量分数。
所有数据均通过Excel 2003进行整理,并绘制图表,方差分析、相关分析以及其他统计分析处理均借助于SPSS 17.0软件。
2 结果与分析
2.1 3个油茶品种光合速率日变化
由图1可知:3个油茶品种胞间二氧化体积分数日变化均呈先降低后维持平衡又升高的特点,基本与净光合速率相反,王春清等[7]发现1 d中净光合速率与胞间二氧化碳呈极显著负相关关系(r=-0.97,n=13,P<0.01)。3个品种的气孔导度变化趋势基本一致,7:00-15:00一直维持在较高水平,15:00以后随着气温和光强的降低迅速下降。大气温度和光合有效辐射的变化趋势相似,从早晨逐步上升,13:00达到高峰,峰值分别为39℃和1800μmol·m-2·s-1,然后缓慢下降。相对湿度的变化则呈V字形,13:00最低,此时的相对湿度为53%。
3个油茶品种晴天的净光合速率随时间变化趋势相似,均呈宽大的单峰型,不存在明显的“光合午休”现象。从7:00开始,随着时间的推移,光合有效辐射逐渐增加,气温也逐渐升高,胞间二氧化碳体积分数迅速下降,净光合速率迅速增加,到9:00净光合速率达到全天的最大值,‘华硕’‘华金’‘华鑫’的净光合速率分别为8.42,9.33和8.97μmol·m-2·s-1;而在9:00以后,光合有效辐射和大气温度进一步升高,空气湿度逐渐下降,净光合速率开始缓慢下降,但下降幅度较小,15:00时的净光合速率与9:00相比,分别降低了24.4%,28.2%和19.4%;15:00-19:00,随着光合有效辐射和气温的逐渐降低,叶片气孔导度逐渐下降,净光合速率开始迅速下降,19:00净光合速率分别降为0.97,0.80和2.07μmol·m-2·s-1。除9:00,1 d中同一时刻‘华鑫’的净光合速率均高于‘华硕’和‘华金’。‘华金’的最大净光合速率最高,‘华鑫’次之,‘华硕’较低。
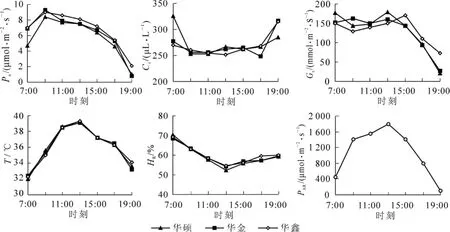
图1 油茶叶片净光合速率(Pn),胞间二氧化碳体积分数(Ci),气孔导度(Gs),叶面温度(T),空气湿度(HR)和光合有效辐射(PAR)的日变化Figure1 Diurnal variation of net photosynthetic rate (Pn), intercellular CO2concentration (Ci) , stomatal conductance (Gs), leaf temperature (T), relative humidity (HR) and Photosynthetically active radiation (PAR) of three cultivars of Camellia oleifera
结合表1可以看出:7:00‘华鑫’和‘华金’的净光合速率极显著高于‘华硕’,19:00‘华鑫’的净光合速率极显著高于‘华硕’和‘华金’,说明‘华鑫’对弱光的利用率较高。11:00-15:00,3个品种的净光合速率没有显著差异。在这期间,光合有效辐射和气温较高(均在1400μmol·m-2·s-1和35℃以上),而油茶的净光合速率变化很小,并保持在一个相当高的水平,说明这3个油茶品种对强光和高温的适应性较强。

表1 净光合速率日变化方差分析Table1 Comparison of the diurnal variation of net photosynthetic rate
2.2 3个油茶品种光合参数的差异
由图2的光响应曲线中可知:在光合有效辐射为0时,净光合速率均为负值;随着光合有效辐射的升高,净光合速率逐渐升高,在光合有效辐射为0~200μmol·m-2·s-1时, 净光合速率几乎呈线性增加;随着光合有效辐射继续升高,各品种的净光合速率增加速率减缓并维持在一定水平;当光合有效辐射继续升高到一定程度后,各品种的净光合速率呈现不同程度的下降,这可能是由于光合有效辐射强度过强,多余的光能造成光合速率下降或者引起光抑制[8]。 光合有效辐射达到 200μmol·m-2·s-1后,3个品种的净光合速率为‘华鑫’>‘华金’>‘华硕’。
表2可以看出:3个油茶品种的光合参数存在差异。最大净光合速率从大到小的顺序为‘华鑫’>‘华金’>‘华硕’,与净光合速率日均值基本相同。3个油茶品种中‘华鑫’的表观量子效率最大,表明其对弱光的利用效率较高。植物的光饱和点和光补偿点分别代表了光照强度与光合作用关系的上限和下限的临界指标,反映了叶片对强光和弱光的利用能力。3个油茶品种的光补偿点顺序为‘华金’>‘华鑫’>‘华硕’,光饱和点顺序为‘华硕’>‘华鑫’>‘华金’。从3个油茶品种的光补偿点和光饱和点来看,光补偿点较低而光饱和点较高,说明其对光照的适应性较强,为较耐荫的喜光树种。
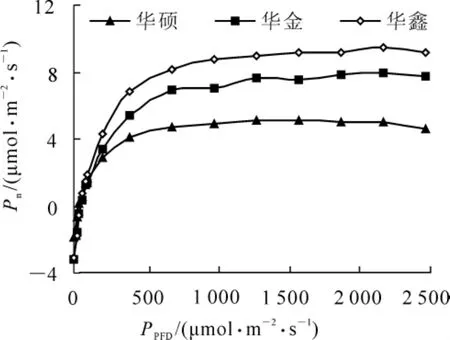
图2 3个品种净光合速率—光量子通量密度(Pn-PPFD)响应曲线Figure2 Responses of the net photosynthetic rate (Pn) to different photosynthetic photon flux densities (PPFD)
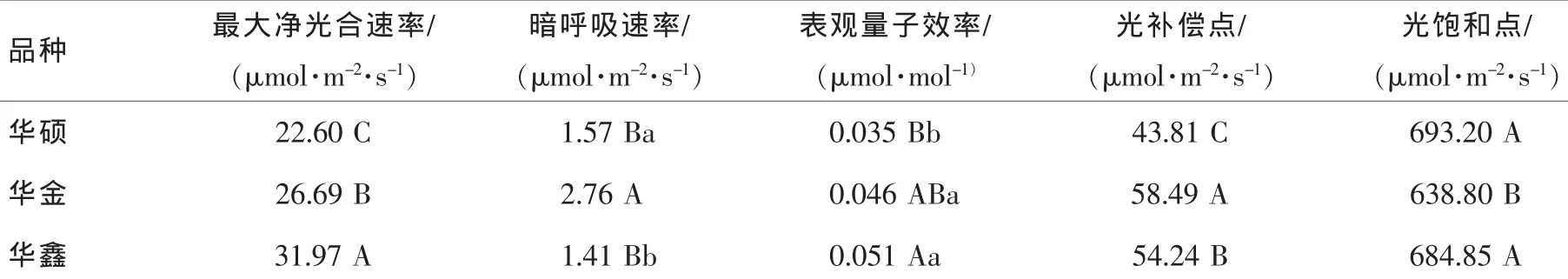
表2 3个油茶品种光合参数的差异Table2 Some differences of photosynthetic parameters in leaves of three Camellia oleifera cultivars
2.3 3个油茶品种叶绿素质量分数的差异
光合色素质量分数将直接影响叶绿素对光能的吸收、传递以及在PSⅡ和PSⅠ之间分配和转换合成三磷酸腺苷(ATP)和还原性辅酶Ⅱ(NADPH),从而影响植物的光合作用。由表3可以看出,3个油茶品种光合色素质量分数存在一定的差异,其中叶绿素a,叶绿素b和总叶绿素均为‘华硕’最高。‘华硕’与‘华金’‘华鑫’的叶绿素a和总叶绿素质量分数之间的差异均达到了极显著水平,而‘华金’和‘华鑫’之间没有显著差异;‘华金’与‘华鑫’的叶绿素b之间的差异达到了显著水平,而与‘华硕’之间没有显著差异。

表3 3个油茶品种叶绿素质量分数方差分析Table3 Comparison of the chlorophyll content of three Camellia oleifera cultivars
由表4可以看出:净光合速率与叶绿素a呈极显著负相关关系,与叶绿素b、总叶绿素和叶绿素a/叶绿素b之间相关性不显著。叶绿素b和总叶绿素间呈极显著正相关关系,是因为叶绿素总量=叶绿素a+叶绿素b,叶绿素a和叶绿素b之间不存在相关性。

表4 叶绿素和净光合速率相关性分析Table4 Correlation of chlorophyll and photosynthetic rate of Camellia oleifera
3 结论与讨论
油茶光合作用日变化有单峰和双峰2种不同的报道[9]。我们的测定结果表明:3个油茶新品种的光合速率日变化均呈宽大的单峰型,这与陈永忠等[10]的报道一致。即使光合有效辐射在1200μmol·m-2·s-1以上,气温在35℃以上,光合速率也一直维持在较高水平,有利于生成大量有机物,提高产量,这可能是3个油茶品种果实大,丰产、稳产的原因。从3个油茶品种的光合日变化可以看出9:00到15:00是1天中光合作用的主要时段。
光饱和点高、光补偿点低的植物对光强的生态适应范围较大,反之较小。由拟合的3个油茶品种的光饱和点、光补偿点结果可知,它们对光强的生态适应范围大小为‘华硕’>‘华鑫’>‘华金’。光响应曲线分析表明:3个油茶品种的光饱和点高,说明其耐高温、喜光,栽培时应选择高温、强光照地段造林,有利于其光合生产力的提高。
对叶绿素和净光合速率进行了相关分析,叶绿素总质量分数和净光合速率之间不存在显著相关性,这与骆琴娅等[10]通过试验得出的研究结果一致,即当叶片发育成熟,叶绿素稳定后,光合速率与叶绿素质量分数之间并没有明显的相关性。但也有研究表明:叶绿素和净光合速率之间存在显著相关性[12-13],其原因可能是研究分析的生长发育阶段不同,其叶绿素质量分数不同,不一定所有的叶绿素都参与了光合作用的进行。至于叶绿素a与净光合速率之间存在极显著负相关关系的原因还需要进一步探讨。
3个油茶品种的光合速率日变化均为宽大的单峰型,与双峰型相比,不存在光合午休现象,即使在中午高温、高光合有效辐射条件下,净光合速率一直维持在较高水平,说明它们对光能有较强的吸收能力,叶片的光合作用强,有利于积累较多的有机物,为花芽分化和果实后熟阶段的油脂转化提供营养,从而提高坐果率和出油率,使油茶稳产高产。‘华硕’具有最低的光补偿点和最高的光饱和点,对光强的适应范围最大,并且叶绿素质量分数最高;‘华鑫’的最大光合速率最高,暗呼吸速率最低、表观量子效率最高;相比而言,‘华硕’和‘华鑫’的光合特性较强。
[1]庄瑞林.中国油茶[M].北京:中国林业出版社,1988:1.
[2]陈隆升,陈永忠,彭邵峰.油茶光合特性研究进展与高光效育种前景[J].湖南林业科技,2011,37(3):33-39.CHEN Longsheng, CHEN Yongzhong, PENG Shaofeng.Research progress on photosynthetic characteristics of Camellia oleifera and the prospect of its high photosynthetic efficiency breeding [J].J Hunan For Sci Technol, 2011, 37 (3):33-39.
[3]唐炜,谭晓风,袁德义.山茶属3个物种光合特性日变化[J].北方园艺,2010(9):5-8.TANG Wei, TAN Xiaofeng, YUAN Deyi.The daily changes of photosynthesis about three species of Camellia genus[J].North Hortic, 2010 (9): 5-8.
[4]李建安,何志祥,孙颖,等.油茶林分光合特性的研究[J].中南林业科技大学学报,2010,30(10):56-61.LI Jian′an, HE Zhixiang, SUN Ying, et al.Study of photosynthetic characteristics in Camellia oleifera stand [J].J Central South Univ For&Technol, 2010, 30 (10): 56-61.
[5]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用 [J].农业基础科学,2005, 21 (8): 76-79.LIU Yufeng, XIAO Langtao, TONG Jianhua, et al.Primary application on the non-rectangular hyperbola model for photosynthetic light-response curve [J].Chin Agric Sci Bull, 2005, 21 (8): 76-79.
[6]郝建军,康宗利,于洋.植物生理学实验技术[M].北京:化工工业出版社,2006:68-70.
[7]王春清,祖容,张贤泽,等.葡萄幼树若干光合特性的研究[J].园艺学报,1989,16(4):279-284.WANG Chunqing, ZHU Rong, ZHANG Xianze, et al.Study on some photosynthetic characteristics of young guapevine [J].Acta Hortic Sin, 1989, 16 (4): 279-284.
[8]POWLES C B.Photo inhibition induced by visible light[J].Annu Rev Plant Physiol, 1984, 35: 15-44.
[9]王瑞,陈永忠,杨小胡,等.油茶光合作用及其影响因素研究进展[J].经济林研究,2007,25(2):78-83.WANG Rui, CHEN Yongzhong, YANG Xiaohu, et al.Literature review of researches on photosynthesis in Camellia oleifera L.and its affecting factors [J].Nonwood For Res, 2007, 25 (2): 78-83.
[10]陈永忠,王瑞,王湘南,等.油茶无性系果实成熟期光合特性的研究[J].经济林研究,2010,28(2):1-6.CHEN Yongzhong, WANG Rui, WANG Xiangnan, et al.Photosynthetic characteristics in Camellia oleifera clones at fruit maturation stage [J].Nonwood For Res, 2010, 28 (2): 1-6.
[11]骆琴娅,漆龙霖,方晰,等.山茶属植物5个物种光合作用的研究[J].林业科学研究,1993,6(3):311-316.LUO Qinya, QI Longlin, FANG Xi, et al.A study on photosynthesis of five species of Camellia genus [J].For Res,1993, 6 (3): 311-316.
[12]SHIGESABURO J.Photosynthetic efficiency in rice and wheat[J].Rice Breed, 1972, 22: 475-479.
[13]EEHIDE M, SHIGESABURO J.Photosynthetic heterosis in maize [J].Jpn J Breed, 1979, 29 (2): 159-165.
猜你喜欢
江苏教育·书法教育(2020年12期)2020-09-10
农业与技术(2020年12期)2020-07-04
农学学报(2019年1期)2019-02-21
民间故事选刊·上(2018年5期)2018-05-18
上海故事(2018年1期)2018-03-02
新课程·下旬(2017年7期)2017-08-14
电脑爱好者(2016年1期)2016-01-14
中国计算机报(2009年34期)2009-09-14
现代计算机(2009年8期)2009-08-12
足球俱乐部(2008年23期)2008-01-09
