
白花泡桐尿苷二磷酸葡萄糖焦磷酸化酶mRNA全序列克隆及序列分析
2012-05-10 02:07熊治国刘志刚李建祥裴海朝高福玲秦采风陈占宽叶金山
河南农业大学学报 2012年3期
熊治国,刘志刚,李建祥,裴海朝,高福玲,秦采风,陈占宽,叶金山,付 晓
(1.河南省绿士达林业新技术研究所,河南郑州450008;2.河南省经济林和林木种苗工作站,河南郑州450008;3.国家林业局泡桐研究开发中心,河南 郑州450003;4.新安县林业局,河南 新安471800)
白花泡桐(Paulownia fortunei(Seem.)Hemsl.),是一种速生优质用材落叶树种,生长快,成材早,繁殖容易,材质好,用途广,经济价值高.木材的材质是由其组分纤维素、木质素等的结构、性质及二者的总量和比率决定的[1].尿苷二磷酸葡萄糖焦磷酸化酶 (UDP-Glucose Pyrophosphorylase,UGPase),催化葡萄糖-1-磷酸(Glc-1-P)生成尿苷二磷酸葡萄糖(UDP-GLC).尿苷二磷酸葡萄糖是纤维素生物合成的基本原料与前体[2].UGP的活性决定着木材中纤维素含量及其与木质素的比率.目前,有些研究者已从一些植物如水稻[3]、黄芪[4]、大麦[5]、马铃薯[6]等中克隆了 UGPase.登录 GenBanK的泡桐mRNA只有8个,且无一与泡桐纤维素、木质素生物合成有关.这意味着目前研究泡桐纤维素生物合成代谢的表达调控尚无基础,搞清这些基因的结构、序列是研究泡桐纤维素生物合成代谢的基础与前提.本研究在对其他物种相关基因序列分析的基础上,采用RT-PCR及RACE(Rapid Amplification of cDNA Ends)方法,克隆并分析了UGP mRNA全序列,旨在丰富白花泡桐基因资源,为植物品质或材质改良提供理论依据.
1 材料与方法
1.1 材料与试剂
白花泡桐由河南农业大学提供.RNA提取试剂盒、AMV逆转录酶、高保真LA Taq、相关限制性核酸内切酶等试剂从宝生物工程(大连)有限公司(TAKARA)购得.
1.2 试验方法
1.2.1 白花泡桐总RNA的提取 取20 cm长的生长旺盛的当年生幼嫩白花泡桐枝条,剥去韧皮部,用清洁刀片刮取木质部外周形成层部分的细胞,液氮研磨成粉,加入试剂 RNAiso Reagent(TAKARA)提取总RNA.
1.2.2 UGP部分片段的获得 (1)RT-PCR:以提取的总RNA为模板,Oligo dT为引物,使用TAKARA生产的反转录酶XL(Reverse Transcriptase XL AMV)反转录得到cDNA.根据NCBI上发表的其他物种UGPase氨基酸的保守序列设计引物[7]BUF(5'-AARGAYGGNTGGTAYCCNCCNGG-3')和BUR(5'-GGRTTNGGDATDATYTCCATYTT-3'),cDNA为模板,TAKARA高保真Taq LA进行RTPCR,扩增UGP部分序列.反应结束后于琼脂糖凝胶中电泳检测结果.(2)序列测定:凝胶回收试剂盒回收目的片段.回收后的PCR产物送宝生物工程(大连)有限公司(TAKARA)进行序列测定.
1.2.3 UGP mRNA 5'上游未知序列的获得 使用TAKARA出品的5'-FULL RACE kit(Code No:D315)来克隆5'上游未知序列.依据获得的白花泡桐UGP部分编码序列,优选了2条特异的嵌套式PCR引物 U.5GSP1(5'-TGATTGCCTGTAAGTTGACCC-3')和 U.5GSP2(5'-GACATATTCTTTGCCCTGCGATA-3').克隆程序包括:1)泡桐总RNA的提取;2)泡桐总RNA去磷酸化处理;3)去帽子反应;4)5'RACE接头的连接;5)反转录反应;6)以外侧的特异性引物U.5GSP1与5'RACE Outer(5'-TACCGTCGTTCCACTAGTGATTT-3')引物进行PCR反应;7)以内侧的特异性引物U.5GSP2与5’RACE Inner(5'- CGCGGATCCTCCACTAGTGATTTCACTATAGG-3')引物进行PCR反应.琼脂糖凝胶电泳检测结果.凝胶回收扩增的片段,克隆于pMD20-T载体,转化E.coli JM109,菌落PCR鉴定的阳性克隆送样测序.
1.2.4 UGP mRNA 3'下游未知序列的获得 使用TAKARA 出品的3'-FULL RACE Core Set Ver.20(Code No:D314)来克隆3'下游未知序列.依据获得的白花泡桐UGP部分编码序列,优选了2条特异的嵌套式PCR引物U.3GSP1(5'-TTGTTATCGCAGGGCAAAG -3')和 U.3GSP2(5'-ACACTAGCTGATGTGAAAGGTGGT-3').具体的克隆程序包括:1)泡桐总RNA的提取;2)套式PCR反应,先以外侧的特异性引物U.3GSP1与3'RACE Outer(5'-TACCGTCGTTCCACTAGTGATTT-3')引物,进行PCR反应,再以内侧的特异性引物U.3GSP2与3'RACE Innerer(5'-CGCGGATCCTCCACTAGTGATTTCACTATAGG-3')引物进行 PCR反应.琼脂糖凝胶电泳检测结果.凝胶回收扩增的片段,克隆于pMD20-T载体,转化 E.coli JM109,菌落PCR鉴定的阳性克隆送样测序.
1.2.5 UGP mRNA全序列的获得及分析 经过以上步骤,分别获得了UGP mRNA的部分序列、5'上游未知序列、3'下游未知序列,且3个序列互有重叠,经Lasergene软件拼接、分析,得到UGP mRNA全序列,提交GenBank.
2 结果与分析
2.1 白花泡桐部分序列的获得
提取白花泡桐的总RNA,经电泳检测其浓度,反转录后以 BUF,BUR为引物,TAKARA高保真Taq LA RT-PCR,电泳结果见图1.将此片段回收,插入pMD19-T vector、转化,挑单菌落鉴定、测序.测序知扩增片段长413 bp,与预期相符.以此DNA片段序列推测可编码137个氨基酸,将其基因序列与NCBI中其它植物的UGPase进行比对,与烟草、马铃薯的UGP序列同源性分别为85%,84%.因此根据相似度可以断定此扩增的片段即为泡桐UGP基因的部分mRNA序列.
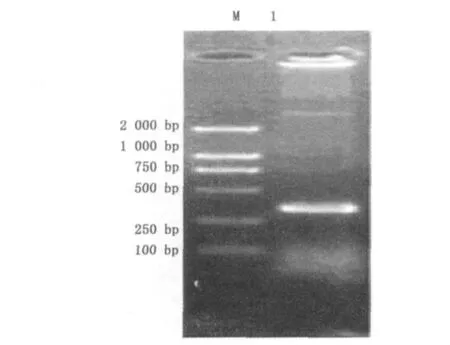
图1 UGP RT-PCR电泳图Fig.1 The UGP of RT-PCR
2.2 UGP mRNA 5'上游未知序列的获得
RACE法得到UGP mRNA 5'上游片段,经琼脂糖凝胶电泳检测结果.凝胶回收扩增的片段,克隆于pMD20-T载体,转化E.coli JM109,菌落PCR鉴定的阳性克隆送样测序,测序结果参见图2.序列中有效序列长777 bp.
2.3 UGP mRNA 3'下游未知序列的获得
RACE法得到UGP mRNA 3'上游片段,经琼脂糖凝胶电泳检测结果.凝胶回收扩增的片段,克隆于pMD20-T载体,转化E.coli JM109,菌落PCR鉴定的阳性克隆送样测序,测序结果见图2.序列中有效序列长815 bp.
2.4 UGP mRNA全序列的获得及分析
使用Lasergene软件,得到UGP mRNA全序列,见图3.UGP mRNA共1 694个碱基,CDS共1 428个碱基(112~1 539),编码氨基酸475(图3).
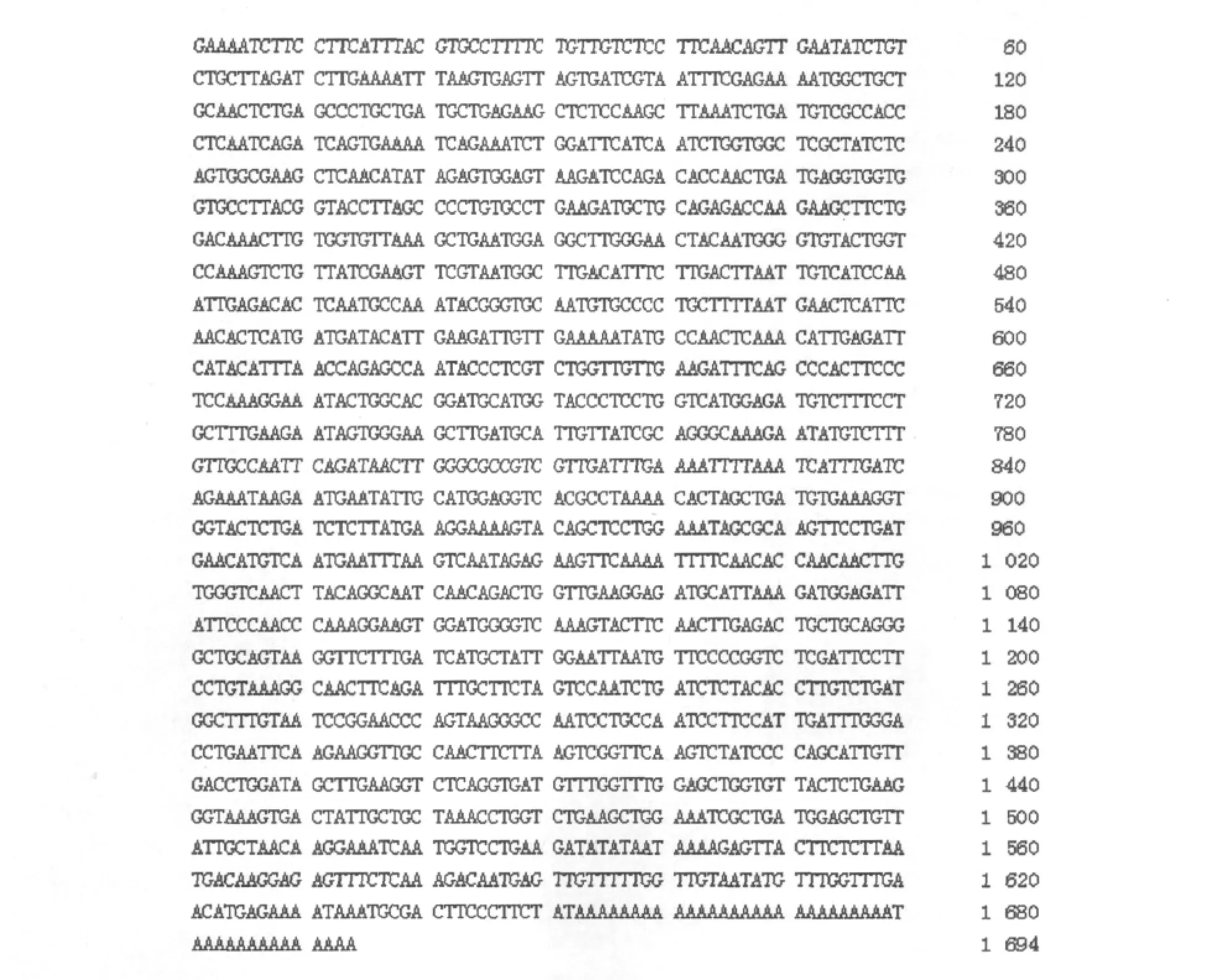
图2 白花泡桐UGP mRNA全序列Fig.2 The complete sequence of UGP mRNA
登陆NCBI网站,利用GenBank BLAST软件对泡桐UGP全长mRNA序列分析比对的结果显示,与烟草的同源性为84%,与马铃薯的同源性为83%,美洲山杨的同源性为 81%.利用 GenBank BLAST软件对泡桐UGP酶氨基酸序列分析比对的结果显示,与烟草相似性为89%,与美洲山杨的相似性为88%,与马铃薯的相似性为88%.

图3 白花泡桐UGP氨基酸全序列Fig.3 The amino acid sequence alignment of UGP
3 讨论
虽然UGP基因已从水稻、黄芪、大麦、马铃薯等几种植物中克隆出来,但用BLAST比较了已知的几种植物的UGP基因核苷酸序列,由于亲缘关系较远,很难找到可用来设计引物的同源性很高的核苷酸片段,只能依据UGP氨基酸保守序列,设计简并引物,通过RT-PCR扩增得到一段cDNA片段,以此片段序列为基础设计特异引物,再通过5'RACE扩增得到了含5'末端的cDNA片段,3'RACE扩增得到含3'末端的cDNA片段.根据以上2个片段的核苷酸序列,设计特异引物,扩增得到了包含完整编码区的全长cDNA.为提高PCR扩增的特异性,采用嵌套PCR扩增.由嵌套引物引导的第2轮PCR可显著提高扩增产物的特异性.RACE是1种以PCR为基础快速分离包含3'和5'末端的cDNA的方法.其优点在于快速简便,仅知少量序列信息时即可扩增得到全长cDNA,多数情况下无需构建cDNA文库.我们在仅知UGP部分氨基酸序列的情况下,未构建cDNA文库,应用RACE这种方法,首次得到了白花泡桐UGP全长cDNA.
对于植物UGPase是否存在多基因家族的问题,早期研究者认为植物中只有1种UGPase,EIMERT等从大麦叶、胚及胚乳的cDNA文库中克隆了11个UGPase序列,结果也表明大麦中只有1种UGPase[5].但后来的研究发现在马铃薯[8,9]、水稻[3]中存在2种相似性很高的UGPase.水稻中存在2种UGPase,其中UGP1位于第9条染色体上,而UGP2位于第2条染色体上.而白花泡桐中是否也存在2种UGPase,还有待进一步研究.
本研究首次克隆了白花泡桐UGP mRNA全序列,构建载体的后续工作正在进行中.希望通过抑制纤维素生物合成的关键酶UGPASE的活性,以此提高木质素的比率,从而提高泡桐木材的硬度.
[1] ANTEROLA A M,LEWIS N G.Trends in lignin modification:a comprehensive analysis of the effects of genetic manipulations mutations on lignification and vascular integrity[J].Phytochemistry,2002,61(3):221 -294.
[2] 连 玲,叶冰莹,何 炜,等.甘蔗 UGPasecDNA的克隆及序列分析[J].福建师范大学学报:自然科学版,2009,25(3):105-108.
[3] ABE T,NIIYAMA H,SASAHARA T.Cloning of cDNA for UDP-glucose pyrophosphorylase and the expression of mRNA in rice endosperm [J].Theor Appl Genet,2002,105(2/3):216-221.
[4] WU X J,DU M,WENG Y Q.UGPase of astragalus membranaceus:cDNA cloning,analyzing and expressing in Escherichia coli[J].Acta Botanica Sinica,2002,44(6):689-693.
[5] EIMERT K,VILLAND P,KILIAN A,et al.Cloning and characterization of several cDNA for UDP-glucose pyrophosphorylase from barley(Hordeum vulgare)tissues[J].Gene,1996,170(2):227 -232.
[6] KATSUBE T,KAZUTA T,TANIZAWA K,et al.Expression in Escherichia coli of UDP-glucose pyrophosphorylase cDNA from potato tuber and functional assessment of the five lysyl residues located at the substratebinding site[J].Biochemistry,1991,30(35):8546 -8551.
[7] 吴晓俊,刘涤,胡之璧.尿苷二磷酸葡萄糖焦磷酸化酶[J].植物生理学通讯,2000,36(3):193 -200.
[8] SOWOKINOS J R,THOMAS C,BURRELL M M.Pyrophosphorylases in potato V.allelic polymorphism of UDP-glucose pyrophosphorylase in potato cultivars and its association with tuber resistance to sweetening in the cold[J].Plant Physiology,1997,113:511-517.
[9] SOWOKINOS J R,VIGDOROVICH V,ABRAHAMSEN M.Molecular cloning and sequence variation of UDP-glucose pyrophosphorylase cDNAs from potatoes sensitive and resistant to cold sweetening[J].Journal of Plant Physiology,2004,161(8):947-955.
猜你喜欢
太原理工大学学报(2021年6期)2021-11-25
中国海洋大学学报(自然科学版)(2019年7期)2019-05-21
中国海洋大学学报(自然科学版)(2019年7期)2019-01-04
中成药(2018年1期)2018-02-02
现代园艺(2017年13期)2018-01-19
作文周刊·小学三年级版(2017年15期)2017-06-22
广州化工(2016年11期)2016-09-02
安徽林业科技(2015年6期)2015-09-25
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
中央民族大学学报(自然科学版)(2014年2期)2014-06-09
