
内蒙古不同草原类型土壤呼吸对放牧强度及水热因子的响应
2012-04-12 03:41杨阳韩国栋李元恒陈志芳王成杰
草业学报 2012年6期
杨阳,韩国栋,李元恒,陈志芳,王成杰
(内蒙古农业大学生态环境学院,内蒙古 呼和浩特010018)
*土壤呼吸(soil respiration)是指未受扰动的土壤中产生CO2的所有代谢过程,它包括3个生物学过程(植物根呼吸、土壤微生物呼吸和土壤动物呼吸)和1个非生物学过程(含碳物质化学氧化过程)。碳素以CO2的形态从土壤向大气流动是土壤呼吸作用的表现形式,它是生态系统碳素循环的主要过程之一,对于调节陆地生态系统碳库和循环起着重要的作用。随着全球气候变化研究成为公众和科学界关注的热点,CO2作为一种最重要的温室气体,其源、汇及通量的精确测定格外得到重视,而土壤呼吸作为土壤碳库碳平衡的一个重要相关过程更是不容忽视。其中,草地生态系统的土壤呼吸是全球土壤呼吸的重要组成部分,对于调节全球C循环具有重大作用。
从20世纪70年代以来,国外对土壤呼吸的相关研究就开始了。我国开展的较晚,早期测定是针对农田排放的CO2,对自然生态系统森林、草原研究的很少。近些年李凌浩等[1]、陈四清等[2]、崔骁勇等[3]对草原土壤呼吸的研究比较多。Fang和Wang[4]对中国土壤呼吸总碳量作了估算(4.2×109t/a),但由于缺乏土壤呼吸的直接观测数据,只能依靠国外的数据类推。研究土壤呼吸的复杂性在于它的变异性与生物、非生物因子的关系上,自然发生的事件和人为扰动都会引起土壤呼吸的变化。目前,学者们对土壤呼吸的研究涉及的草原类型较多,研究方向多集中在不同时间尺度[5-7](日尺度、季节尺度、年际间)和控制因子[8,9](土壤温度、土壤含水量、降水和土壤碳氮等非生物因子)上,对不同空间尺度以及放牧干扰的草原土壤呼吸作用的比较研究较少。
内蒙古草原是欧亚大陆草原的重要组成部分,也是我国畜牧业的重要生产基地。放牧是内蒙古草原的主要利用方式,放牧强度、放牧制度、放牧季节和放牧家畜种类都在很大程度上影响着草地生态系统的物种组成、草地生产力及土壤的理化性质[10]。放牧导致生物因子和非生物因子发生变化,进而导致了土壤呼吸速率的变化。许多研究表明[1,7,11],适度放牧使草地生物量、根系和凋落物数量、土壤动物和微生物的多样性提高,从而促进土壤呼吸。过度放牧导致草地退化、盖度降低、草地地上和地下生物量下降,草地碳汇功能减弱[8,12],从而降低土壤呼吸速率。
为了充分认识草地生态系统和土壤呼吸的关系,需要针对实际情况,在不同的草原群落进行大量的野外定位实验研究工作,立足碳循环过程的整体[7],综合考虑未来全球气候变化和人类活动干扰的可能影响。鉴于此,本研究选取内蒙古具有代表性的草甸草原(锡林郭勒盟西乌珠穆沁旗)、典型草原(赤峰市克什克腾旗)和荒漠草原(乌兰察布市四子王旗)典型牧户的不同放牧强度梯度草场,探讨不同类型草原在不同放牧强度下土壤呼吸变化规律以及与生物和非生物因子的相关性,为内蒙古草原放牧管理方式的准确评估和草原碳增汇减排对策的制定提供参考依据。
1 材料与方法
1.1 研究区概况
草甸草原试验区位于锡林郭勒盟西乌珠穆沁旗境内(东经117°43′20″,北纬44°36′45″),海拔1 090 m。试验区属于温带半干旱气候,年平均气温1℃,年平均降水量390 mm,年平均蒸发量1 600 mm。植物群落由40多种植物组成,建群种为羊草(Leymuschinensis),伴生植物种为贝加尔针茅(Stipabaicalensis)和线叶菊(Filifolium sibiricum)等。土壤类型为暗栗钙土。
典型草原试验区位于赤峰市克什克腾旗西部达里诺尔国家级自然保护区境内(东经116°38′~116°41′,北纬43°25′~43°27′),海拔1 370 m。试验区属于中温型大陆性气候,年平均气温为1~2℃,年平均降水量350 mm,年平均蒸发量1 643 mm,植物种类丰富,建群种是大针茅(Stipa grandis),优势种有羊草、糙隐子草(Cleistogenes squarrosa)及冷蒿(Artemisiafrigida)等。土壤类型为典型栗钙土。
荒漠草原试验区位于乌兰察布市四子王旗王府一队(东经111°53′46″,北纬41°47′17″),海拔1 456 m。试验区属于典型的中温带大陆性气候,年平均气温3.4℃,年平均降水量为248 mm,年平均蒸发量2 947 mm,植被类型为短花针茅+冷蒿+无芒隐子草,草层低矮,且植被较稀疏,建群种为短花针茅(Stipabreviflora),优势种为冷蒿、无芒隐子草(Cleistogenessongorica)。土壤类型为淡栗钙土。
以上3个草原类型的盖度和生物量概况见表1。试验区的放牧方式为原始连续自由放牧,放牧时间为5月上旬-11月上旬,放牧家畜以绵羊为主。
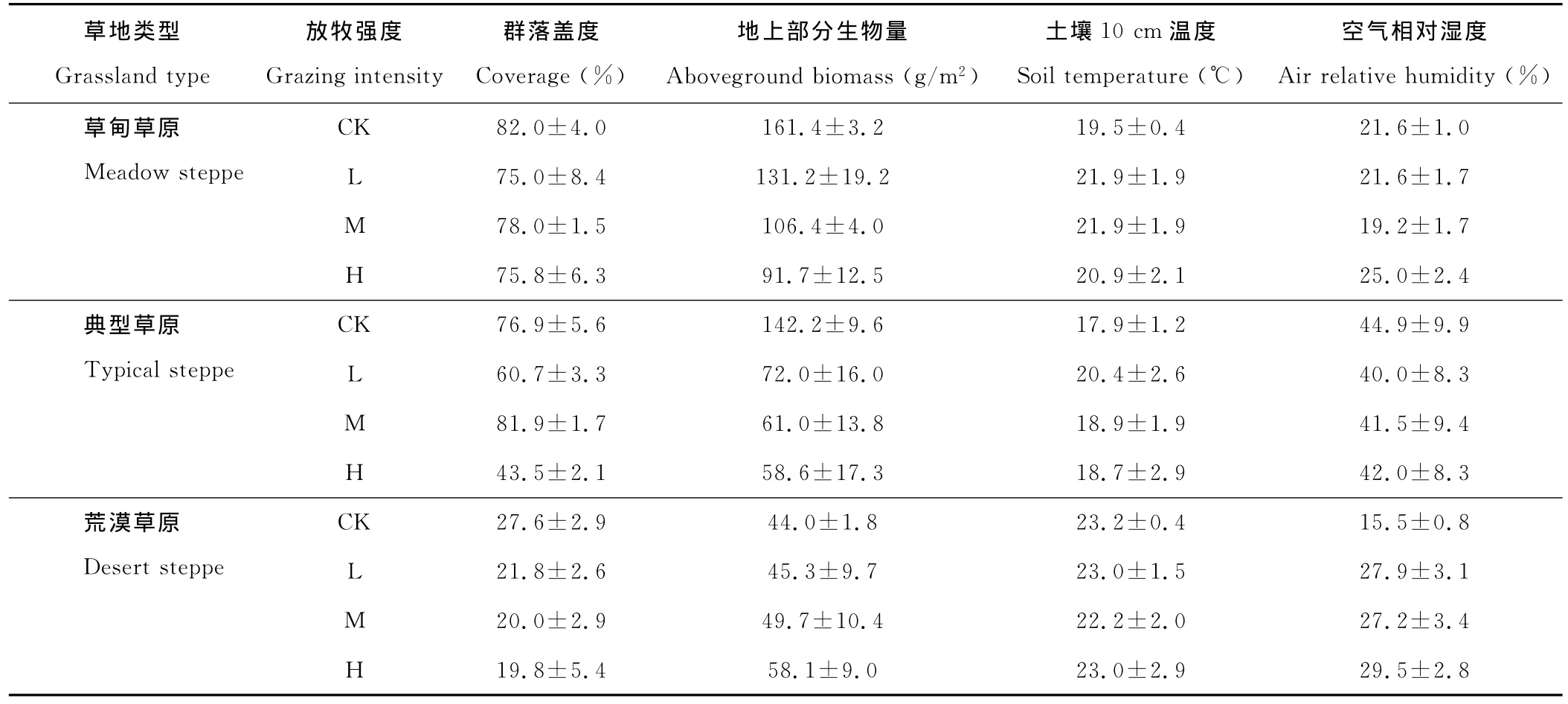
表1 试验样地植被群落盖度和地上部分生物量的本底调查(2009年)和土壤10cm温度与空气相对湿度(2010年)Table 1 Coverage and aboveground biomass of plant communities(2009),10cm soil temperature and air relative humidity(2010)in experimental sites
1.2 研究设计
不同放牧强度的划分是按照李博[13]退化草地分级方法,分为4个放牧强度梯度:零放牧的围封区为对照(un-grazing,CK)样地、轻度放牧样地(light grazing,LG)、中度放牧样地(moderate grazing,MG)和重度放牧样地(heavy grazing,HG)。具体设计方法是按照草原群落随着放牧强度的变化而进行划分,沿半径方向构成草原群落的不同放牧梯度:居民点或家畜饮水点周围放牧强度较大,远离居民点或家畜饮水点的另一端放牧强度较轻。采样时,以居民点一端为起点至零放牧的对照区,设置3条样带(样带长约5 000 m),作为3次重复样带,每个样点进行3次重复取样。
1.3 数据采集
本实验采用Li-8100便携式土壤呼吸测定仪(Li-Cor,Inc.,Lincoln,NE,USA)测定土壤呼吸。在实验进行前1 d埋入一个聚氯乙烯(polyvinylchloride,PVC)土壤呼吸环(内径22 cm,高度5 cm),一端削薄,露出地面部分高度为2~3 cm,保持各样地PVC套环地上部分环内高度一致。如果土壤环内有绿色植物即用剪刀齐地剪去地上部分,以消除测定土壤呼吸时植物自养呼吸的影响。测定时间在2010年8月15日-8月30日,每次测定在上午9:00-11:00完成[14]。土壤10 cm层的温度、空气相对湿度利用Li-8100附件探头自动测定记录。
1.4 数据处理
统计分析采用SAS 9.0(SAS Institute Inc,2002,Cary,NC)软件完成,使用Sigmaplot 10.0和 Microsoft Excel软件作图。采用两因素方差分析(two-way ANOVAs)测试不同草原类型、不同放牧强度及它们交互作用对土壤呼吸的影响,同时采用单因素方差分析(one-way ANOVA)和LSD最少显著差异法检验各草原类型不同放牧强度间土壤呼吸的差异。土壤呼吸与温湿度间的相关性,采用非线性相关分析。
2 结果与分析
2.1 土壤呼吸速率对草原类型和放牧强度的响应
通过以草原类型和放牧强度为主效应的两因素方差分析,不同的草地类型和放牧强度及交互作用都对土壤呼吸有显著影响(表2,P<0.05)。运用单因素方差分析对每一类草地类型中不同放牧强度对土壤呼吸的分析得出,显著性差异主要表现在荒漠草原(P<0.001)和典型草原(P<0.05),而草甸草原不同放牧强度间土壤呼吸差异不显著(表3,P>0.05)。其中,荒漠草原、典型草原和草甸草原生态系统不同放牧强度处理平均土壤呼吸速率分别为2.72,2.12,3.49μmol CO2/(m2·s),草甸草原生态系统整体土壤呼吸速率最强,荒漠草原生态系统次之,典型草原生态系统最弱。
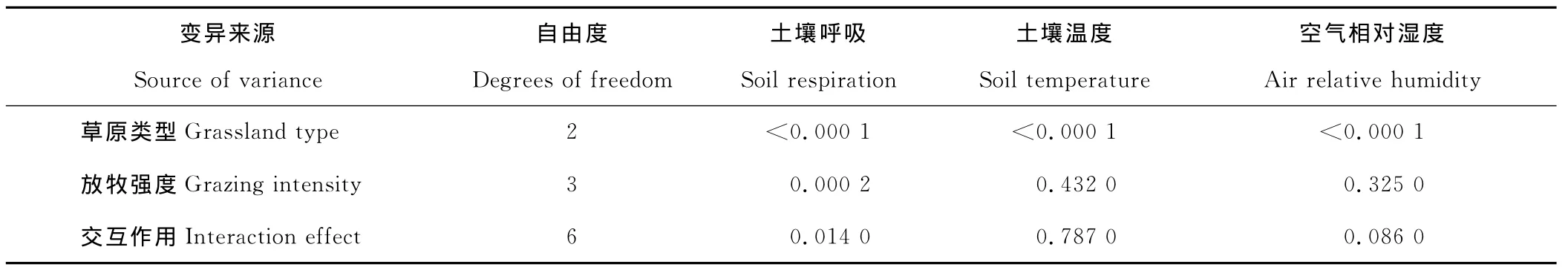
表2 不同草原类型、放牧强度对土壤呼吸、土壤温度、空气相对湿度的两因素方差分析Table 2 Two-way ANOVAs of soil respiration,soil temperature and air relative humidity in different grassland types and grazing intensities

表3 不同放牧强度对土壤呼吸、土壤温度、空气相对湿度的单因素方差分析Table 3 One-way ANOVA of soil respiration,soil temperature and air relative humidity in different grassland types under different grazing intensities
经对不同草原生态系统在4个放牧强度下的方差分析,得出适度的放牧强度能增加土壤呼吸速率。随着放牧强度的增加,都呈先升高后降低的变化规律(图1)。在荒漠草原生态系统中,轻度和中度放牧强度下土壤呼吸显著高于对照和重度放牧区(P<0.05,图1),土壤呼吸速率最大是轻度放牧区,达3.91μmol CO2/(m2·s),最低是对照区,是轻度放牧区的40%。在典型草原生态系统中,仅中度放牧和重度放牧之间土壤呼吸有显著差异(P<0.05,图1),中度放牧下土壤呼吸最高(2.61μmol CO2/m2·s),重度放牧最低(1.56μmol CO2/m2·s),它们之间土壤呼吸相差1.05μmol CO2/(m2·s)。在草甸草原生态系统中,不同放牧强度间土壤呼吸无显著差异,轻度放牧土壤呼吸最高,为4.02μmol CO2/(m2·s)。
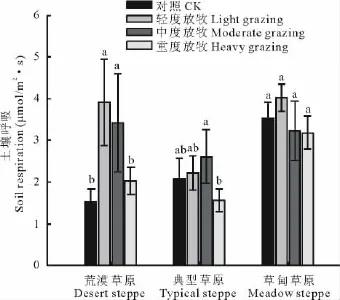
图1 各草地类型不同放牧强度下土壤呼吸的变化Fig.1 Changes of soil respiration under different grazing intensities in different grassland types
2.2 不同草原类型和放牧强度对土壤呼吸及水热因子的影响
以草原类型和放牧强度为主效应作两因素方差分析(表2)和以放牧强度为主效应,对每一草原类型作单因素方差分析(表1,3)。结果显示,不同草原类型对土壤10 cm层温度和空气相对湿度有极显著性影响(P<0.001);不同放牧强度对土壤10 cm层温度和空气相对湿度无显著性影响(表2,P>0.05)。荒漠草原中,放牧强度对土壤10 cm层的温度没有影响,各强度土壤温度都保持在22~23℃。而典型草原和草甸草原中土壤温度对不同放牧强度有显著影响(P<0.05,表1,3),零放牧强度都表现出最低的土壤温度,分别为17.9和19.5℃,轻度放牧土壤温度最高,分别为20.4和21.9℃;典型草原中,放牧强度对空气相对湿度没有显著影响(P>0.05),零放牧强度空气相对湿度最高,达到44.9%,最低为轻度放牧强度,为40%,而荒漠草原和草甸草原中均有显著性影响(P<0.05,表1,3)。
2.3 土壤呼吸和土壤温度以及空气相对湿度的关系
在进行相关分析时,考虑到放牧强度的影响,具体分析每一类草原生态系统中各放牧强度与土壤温度和空气相对湿度的关系时,同一类草原生态系统,不同放牧强度呈现的回归关系不同。相同放牧强度下,与不同环境因子的回归关系不同(表4)。在荒漠草原中,4个放牧强度的土壤呼吸与土壤温度都呈二次相关,但只有中度放牧达到显著水平(表4,P<0.05);与空气相对湿度都呈指数相关,只有重度放牧下没有达到显著差异(P>0.05)。在典型草原中,4个放牧强度的土壤呼吸与土壤温度呈二次相关和指数相关,其中对照和中度放牧下相关性达到显著水平(P<0.05),轻度放牧和重度放牧下二者呈二次相关,其中重度放牧达到显著水平(表4,P<0.05)。与空气相对湿度的关系中,除轻度放牧呈显著二次相关,其他3个放牧强度下都呈指数相关,其中重度放牧下相关性没有达到显著水平。在草甸草原中,除中度放牧的土壤呼吸与土壤温度呈显著二次相关,其他都呈指数相关,重度放牧下达到显著水平(P<0.05)。与空气相对湿度都呈二次相关,但只有重度放牧下相关性达到显著水平(P<0.05)。
3 讨论
土壤呼吸的变异性受到自然条件和人为干扰的影响,不同群落类型的植物组成、气象条件和土壤环境会导致土壤中产生CO2的量也不尽相同[15],人类活动也日益显著的改变了土壤呼吸的特征,影响着土壤CO2产生和传输的各个环节,其中土地利用方式的改变对其影响十分明显。
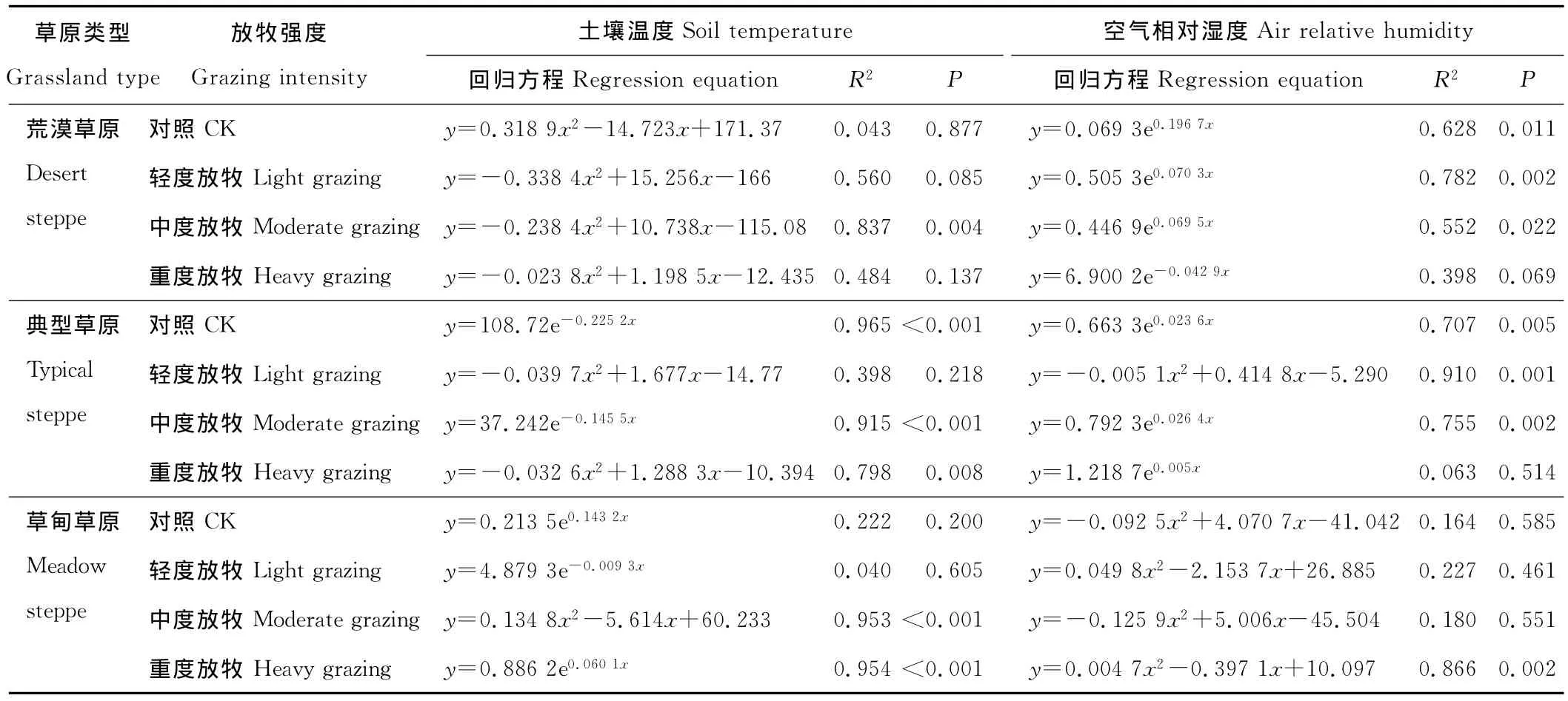
表4 各草原类型下不同放牧强度的土壤呼吸与土壤温度和空气相对湿度的回归关系Table 4 Regression relations among soil respiration of different grassland types,10 cm soil temperature and air relative humidity under different grazing intensities
土壤呼吸释放的CO2中,大部分来自植物光合作用固定的碳。目前在关于土壤呼吸的调控方面,大多学者的实验证明来自冠层光合作用的底物供应是影响土壤呼吸的主要因素。Li等[16]等对锡林河流域羊草群落地上活生物量与土壤呼吸间的相关分析得出二者为显著乘幂的结论。Janssens和Lankreijer[17]通过对18个欧洲森林系统进行研究发现,在区域尺度上,土壤呼吸与生态系统生产力关系密切。Verburg等[18]的温室试验历经2个生长季的观测表明地上部分光合作用直接控制着土壤呼吸。也有一些间接研究[19,20]证明了以上观点,如土壤的环境调控着对糖类的需求,而光合作用决定了地上部分糖类的供应能力。内蒙古草原属于温带草原,植被状况从大兴安岭湿润、半湿润的森林区,以及靠近森林一侧的半湿润的草甸草原,向西过渡到典型草原、荒漠草原,直至极旱荒漠,导致植被覆盖度和植物多样性的差异,使不同草原类型下的草地生态系统整体土壤呼吸速率有显著差异,本实验结论支持了此观点。
不同放牧强度对土壤呼吸有显著影响。Wan和Luo[21]在整个1年的研究期间持续剪掉地上部分生物量,使刈割地保持裸地状态,测得年平均CO2通量降低33%。在美国大平原的草地中,研究人员用刈割和遮荫的方法,发现刈割和遮荫使土壤呼吸在1周内降低了近70%。Bremer等[22]研究了3个刈割处理(早期刈割、整个季节均进行刈割、不刈割)下的土壤呼吸,发现刈割后的第2天土壤呼吸降低了21%~49%。通常,放牧在生长季移走活的生物量,改变了植物冠层结构,进而改变了植物根系分泌物的量;另外,牲畜的践踏以及尿液和粪便输入会改变土壤微环境,从而使土壤呼吸速率降低。也有部分学者根据生态学中经典的“中度干扰假说”,结合“植物-动物间互作的补偿性”观点,提出放牧优化假设,认为草地植被生物量与放牧强度之间的关系是非线性的,即随着放牧强度的增加,草地的生物量先增加[23],然后随放牧强度的增加而下降,中间拐点之后即是过度放牧。本研究所选取的3个草原类型,均以禾本科植物为优势种或建群种,适当强度的放牧可以促进草地植物的分蘖和繁殖,有利于补偿性生长[24]。但长期高强度的放牧会导致草地植物的退化和死亡,可以解释本研究中“随着放牧强度增加,土壤呼吸呈先高后低”的变化规律。
土壤温度对呼吸的影响是多方面的。通常土壤呼吸存在一个温度响应曲线,45~50℃为临界值,在此之前,土壤呼吸速率随温度升高而呈指数升高,随后开始下降。在此响应过程中,呼吸酶最大活性(Vmax)被认为是主要限制因子[25]。也有学者通过控制性实验,证明温度间接影响根系的伸长生长、植物的物候和在土壤团聚体水平上的底物、氧气的运输[26]。土壤湿度是影响土壤呼吸的另一重要的影响因子,很多野外研究结果表明土壤湿度只有在最高和最低的情况下才会抑制土壤CO2通量[27]。本实验从不同草地类型来看,土壤呼吸与地下土壤10 cm层温度、空气相对湿度间的关系密切。在水分成为主要限制因子的荒漠草原,土壤呼吸速率与空气相对湿度多表现为正相关关系,即随着湿度的增大,土壤呼吸速率有所增强。多数控制性实验证明,土壤呼吸通常对最限制它的因子产生响应,即当土壤含水量较低时,土壤呼吸对温度的反应不敏感;在水分不受限制的草甸草原,土壤呼吸速率多随着地下10 cm层温度的升高而升高;在典型草原中,水热配置较好,土壤地下10 cm层温度、空气相对湿度二者交互影响着土壤呼吸。本研究表明在相同草地类型下,不同放牧强度的土壤呼吸对土壤10 cm层温度和空气相对湿度响应各异,但均与土壤呼吸呈指数方程或二次方程进行拟合,与周广胜[28]总结的近年来国内外科学家开展的大量的土壤呼吸作用定量评估方面的研究一致。
值得补充说明的是,土壤呼吸的各影响因素之间并不是孤立的,不仅同时对土壤呼吸产生影响,而且它们之间也有相互影响,同时,在呼吸通量测量方面也不能只对短期流量、季节动态及相应的影响因子进行分析,应结合测定和计算出的土壤呼吸的年度总量数据资料,更合理、更准确地描述内蒙古不同草原类型,乃至全球范围内的土壤呼吸对人为干扰及水热因子的响应问题。
[1] 李凌浩,王其兵,白永飞,等.锡林河流域羊草草原群落土壤呼吸及其影响因子的研究[J].植物生态学报,2000,24:680-686.
[2] 陈四清,崔骁勇,周广胜,等.内蒙古锡林河流域大针茅草原土壤呼吸和凋落物分解的CO2排放速率研究[J].植物学报,1999,41(6):645-650.
[3] 崔骁勇,王艳芬,杜占池.内蒙古典型草原主要植物群落土壤呼吸的初步研究[J].草地学报,1999,7(3):245-250.
[4] Fang J Y,Wang W.Soil respiration as a key belowground process:issues and perspectives[J].Journal of Plant Ecology(Chinese Version),2007,31:345-347.
[5] 陈海军,王明玖,韩国栋,等.不同强度放牧对贝加尔针茅草原土壤微生物和土壤呼吸的影响[J].干旱区资源与环境,2008,22:165-169.
[6] 白哈斯.不同放牧率对羊草+苔草低地草甸草地再生和净初级生产力的影响[J].干旱区研究,2007,24(6):826-829.
[7] 鲍芳,周广胜.中国草原土壤呼吸作用研究进展[J].植物生态学报,2010,34(6):713-726.
[8] 徐海红,侯向阳,那日苏.不同放牧制度下短花针茅荒漠草原土壤呼吸动态研究[J].草业学报,2011,20(2):219-226.
[9] 周萍,刘国彬,薛萐.草地生态系统土壤呼吸及其影响因素研究进展[J].草业学报,2009,18(2):184-193.
[10] 郭明英,卫智军,运向军,等.放牧对草原土壤呼吸的影响[J].草业科学,2011,28(5):729-736.
[11] 陈全胜,韩兴国,阎志丹,等.典型温带草原群落土壤呼吸温度敏感性与土壤水分的关系[J].生态学报,2004,24(4):831-836.
[12] 陈全胜,李凌浩,韩兴国,等.温带草原11个植物群落夏秋土壤呼吸对气温变化的响应[J].植物生态学报,2003,27:441-447.
[13] 李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[14] Xu M,Qi Y.Soil-surface CO2efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California[J].Global Change Biology,2001,7:667-677.
[15] 刘伟,王继明,王智平.内蒙古典型草原植物功能型对土壤甲烷吸收的影响[J].植物生态学报,2011,35(3):275-283.
[16] Li L H,Han X G,Wang Q B,etal.Correlations between plant biomass and soil respiration in aLeymuschinensiscommunity in the Xilin River Basin of Inner Mongolia[J].Acta Botanica Sinica,2002,44(5):593-597.
[17] Janssens I A,Lankreijer H.Productivity overshadows temperature in determining soil and ecosystem respiration across European forests[J].Global Change Biology,2001,7(3):269-278.
[18] Verburg P J,Arnone III J A,Obrist D,etal.Net ecosystem carbon exchange in two experimental grassland ecosystems[J].Global Change Biology,2004,10:498-508.
[19] Nadelhoffer K J.The potential effects of nitrogen deposition on fine-root production in forest ecosystems[J].New Phytologist,2000,147:131-139.
[20] Burton A J,Pregitzer K S,Hendrick R L.Relationships between fine-root dynamics and nitrogen availability in Michigan northern hardwood forests[J].Oecologia,2000,125(3):389-399.
[21] Wan S,Luo Y.Substrate regulation of soil respiration in a tallgrass prairie:Results of a clipping and shading experiment[J].Global Biogeochemical Cycles,2003,17:1054,doi:10.1029/2002GB001971.
[22] Bremer J D,Ham J M,Owensby C E,etal.Responses of soil respiration to clipping and grazing in a tallgrass prairie[J].Journal of Environmental Quality,1998,27:1539-1548.
[23] 陶冶,张元明.3种荒漠植物群落物种组成与丰富度的季节变化及地上生物量特征[J].草业学报,2011,20(6):1-11.
[24] 彭祺,王宁,张锦俊.放牧与草地植物之间的相互关系[J].宁夏农学院学报,2004,25(4):76-79.
[25] Atkin O K,Tjoelker M G.Thermal acclimation and the dynamic response of plant respiration to temperature[J].Trends in Plant Science,2003,8:343-351.
[26] Fu S,Cheng W X,Susfalk R.Rhizosphere respiration varies with plant species and phenology a greenhouse pot experiment[J].Plant and Soil,2002,239:133-140.
[27] Xu L,Baldocchi D D,Tang J.How soil moisture rain pulses and growth alter the response of ecosystem respiration to temperature[J].Global Biogeochemical Cycles,2004,18,doi:10.1029/2004GB002281.
[28] 周广胜.土壤呼吸作用普适性评估模型构件的设想[J].中国科学,2008,38(3):293-302.
猜你喜欢
小学生优秀作文·时尚版·中年级(2022年2期)2022-02-18
散文诗(2021年22期)2022-01-12
散文诗(2021年22期)2022-01-12
幼儿100(2020年31期)2020-11-18
疯狂英语·初中版(2019年4期)2019-09-10
小太阳画报(2018年6期)2018-05-14
辽宁林业科技(2017年4期)2017-06-22
浙江大学学报(工学版)(2016年2期)2016-06-05
大气科学(2015年5期)2015-12-04
儿童故事画报(2015年6期)2015-08-17
