
刈割对干热河谷海狮苏丹草生长的影响
2012-03-18 06:31:50史亮涛何光熊张明忠纪中华龙会英沙毓沧邵庆勇
草业科学 2012年11期
史亮涛,何光熊,张明忠,纪中华,龙会英,金 杰,沙毓沧,邵庆勇
(1.云南省农业科学院热区生态农业研究所,云南 元谋 651300; 2.云南省畜牧兽医科学院,云南 昆明 650224)
苏丹草(Sorghumsudanense)属禾本科高粱属,其饲用价值在国内外得到较高评价[1-2],在渔业中有“养鱼青饲料之王”的美誉;同时苏丹草具有较好的抗逆性[3],防风固沙、保持水土、维持生物多样性等多样化的生态服务功能[1,4-5],其内部生氰苷可作为电气化的持续源动力[6];苏丹草与高粱(S.bicolor)无生殖隔离[7],杂交种具有多种功能且效益明显[8-9],对发展畜牧业意义深远。
稳态(Stability)是指生物体在应对多变环境条件时在一定范围内保持自身状态的能力[10],生物体达到稳态状态的能力在很大程度上与该物种的适应能力及生态对策有关[11-12]。生物体的稳态有多种形式,已有众多学者从生物体维持体内N及P等元素平衡的角度就细菌[13]、真菌[14]、浮游动物[15]、水藻[16]、维管植物[17]等生命有机体对生物体的内稳态(Homeostasis)进行了研究,也有学者从生物体稳态的角度就生物多样性―生态系统功能(Biodiversity effects on ecosystem function,BDEF)展开了研究。有证据显示,豆科和禾本科草地牧草空间生态位互补实现稳态是草地生态系统实现生态构建和功能的主要机制[18],多样性高的草地生态系统可通过单种或多种植物群落空间和时间的生态位互补使系统内植物群落的总生物量提高[19-21],同时物种丰富度及均匀度高的草地生态系统具有较高的生产力[22]。但单一物种的草地,如苏丹草[8]同样具有较高的生产力,而刈割管理可以通过牧草的补偿性生长作用和均衡性生长特性影响牧草个体再生性能和群落生物量,从而影响群落的生产力[23-24]。依据最终产量恒值法则(Law of constant final yield)[25],单作群落在一定条件下能形成稳定的产量。那么单作植物群落通过平衡个体生长的动态作用从而使单一植物群落的总生物量在一定时间和范围内达到相对稳定的状态是极有可能的,已有学者从单作群落有限资源的利用分配策略进行了论述[26]。遗憾的是目前鲜有单作群落稳态及其稳态策略的机制等方面的报道。针对这一问题,本研究以苏丹草为试验对象在不同刈割条件下对这一设想进行验证,并探寻这一机制形成的可能途径,旨在丰富生物多样性与生态系统生产力关系的相关研究内容,同时为单作优质牧草草地管理提供理论参考。
1 材料与方法
1.1试验地自然概况 试验地位于金沙江干热河谷典型区元谋县云南省农业科学院热区生态农业研究所牧草试验研究基地,地处101°35′ E,25°23′ N,属南亚热带干热季风气候;光热资源充足,年均日照时间2 670.4 h,年均气温21.9 ℃,极端最高气温42 ℃,极端最低气温-0.8 ℃,年积温7 378~8 418 ℃·d;雨季集中于5―10月,7月降水最多,年均降水量616.7 mm,蒸发量为降水量的6.4倍,年均相对湿度53%。
1.2试验设计与测定指标
1.2.1试验设计 采用完全随机区组设计,设计留茬5、10、15 cm 3个刈割水平。因单位面积分蘖数和单株高度均会影响苏丹草单位面积产草量,该试验根据密度自疏原则[27],采用高密度撒播种植建立试验群落,规避单位面积分蘖数对试验结果的影响。当建立的单作群落生长到育穗期时刈割留茬;将苏丹草育穗期刈割处理后至下一育穗期时定义为第1个生长阶段,记录每一生长阶段期间苏丹草株高生长动态、分蘖、达到育穗期所需时间及单位面积产量变化;每处理至其单位面积产量低于第1生长阶段平均产量1/5时停止试验。小区面积2 m×2.5 m,小区间设置1 m宽隔离带;每个样方四周去除50 cm边际植株后均匀取样5株标记作为固定观测样本,分别观测单株分蘖数和株高。
1.2.2试验材料 试验采用海狮苏丹草作为研究对象,种子由百绿国际草业(天津)有限公司提供。
1.2.3田间管理 2010年6月20日采用条播法播种,行距20 cm,播种量约3.0 g·m-2,播种后覆盖一层浅土,在苗期保持土壤湿润;试验分别于2010年8月12日、9月4日、10月4日、11月23日和2011年2月5日进行5次刈割,前4次刈割2~3 d后人工去除田间杂草,浇水1次,施农用复合肥(1 g·m-2) 1次。
1.2.4测定指标
补偿生长期:每次刈割后到下一次刈割时为一个补偿生长期。
单位面积生物量:海狮苏丹草孕穗期进行刈割,留茬高度设5、10、15 cm 3个处理。测产时先去除小区四周各50 cm内植株,将余下部分刈割称量,按实际面积计算地上生物量。
株高:在每个生长期内每隔3 d于08:00对固定观测样本进行植株生长高度测定,测量从地面至植株的自然状态最高部位的绝对高度,测量5株求其平均值,记录植株生长高度。第1次株高测定于2010年8月16日进行,第1生长期共进行6次株高测定,第2生长期共测定10次株高,第3生长期共测定10次株高,第4生长期共测定15次株高。
单株分蘖数:在每个小区内随机选5株植株,每次刈割后统计每株分蘖数,求其平均值,得出每次刈割后每个处理单株分蘖数。
密度:每次刈割后测定各小区内苏丹草(株数)丛数。
1.3数据处理 原始数据分析采用SPSS 17.0统计软件进行相关分析、线性拟合和方差分析,多重比较采用最小显著差数法(LSD)检验,绘图采用Microsoft Excel软件辅助。
数据分析中取得的原始数据中植株个体生长高度随时间变化的情况以SPSS 17.0 Curve Estimation模块进行Logistic、Linear、Growth 3种模型线性拟合,结果显示Linear Model能较好地反映苏丹草株高生长的动态(R2>0.92,P<0.5),故选择其建立最优模型分析体系。
Linear-Model分析模型的构建(图1):植物的生长复合Logistic模型的变化规律,对于特定生物个体而言,其达到最大生物量积累和最大生物量所经历的时间是有界限的,二者之间相互作用的范围可用效应区(Effect cage,E)来表示(图1)。若对植物的某个阶段生长情况用线性模型(Liner-Model)来描述,可建立植物生物量y随时间变量x变化的回归曲线y=ax+b(图1)。由线性函数y=ax+b的特征及植物的生长情况可知,在这个模型中,斜率a的正负情况反映植物生物量积累随时间变化的增减,而其大小则反映出植物生物量积累随时间变化的快慢,代表了某个生物量性状的平均累积速率;在a确定的情况下,函数截距b决定了直线y=ax+b在效应区范围内到达某一特定生物量(y值)的表达效果(图1线段l1、l2、l3),即反映出生物达到某一特定生物量y′所需要的生长时间x′,代表植物实现这个生物量性状的潜在生长能力;a和b两个参数共同决定生物个体在效应区(E)内生长的动态变化情况。

图1 Linear-Model分析模型示意图Fig.1 Sketch map of Linear-Model
2 结果与分析
2.1单一牧草群落地上生物量的稳定性 将不同刈割处理条件(留茬5、10、15 cm)及单一牧草群落年生物量统计数据进行方差分析,结果显示(表1):虽然10 cm刈割处理的单位平均产量较高,但不同刈割处理条件下单一牧草群落地上生物量之间差异不显著(P>0.05)。将不同刈割处理条件下同一生长期3个刈割水平总生物量数据进行单因素方差分析,所得结果显示:在第1-3个补偿生长期同一(刈割茬数)生长时期单一牧草群落地上生物量之间差异不显著,而在第4个补偿生长期总生物量急剧减少,且与前3个生长期之间总生物量差异显著(P<0.05)。第1-3个补偿生长期总生物量的变化情况可证明,在特定时期和条件下,单一植物群落的总生物量在一定范围内达到相对稳定的状态。
2.2个体株高和单株分蘖数与地上生物量的关系 在不考虑刈割作用影响的条件下,将单个试验小区内样本植株单株分蘖数作为控制变量分析植物平均株高与群落总地上生物量的关系,可知:在去除单株分蘖数的影响后,海狮苏丹草个体平均株高与群落总生物量呈显著正相关(P<0.01,r=0.626)。同样,在考虑刈割作用的条件下,海狮苏丹草个体平均株高与群落总生物量呈显著正相关(P<0.001,r=0.665)。

表1 不同刈割管理水平及不同补偿生长时期海狮苏丹草地上生物量的变化情况Table 1 Biomass of Sorghum sudanense (Hay-King) in different mowing management and compensatory growing period
2.3个体株高生长动态与群体地上生物量的Linear-Model模型分析 将所得苏丹草植株生长高度随时间变量变化的回归方程y=ax+b中的a、b值进行相关分析,结果表明,在一个生命周期内,不同刈割处理条件下海狮苏丹草单一植物群落个体在生长过程中表现出平衡自身生长速率与生长潜力关系的策略。相关分析表明:海狮苏丹草个体株高的平均生长积累速率a与海狮苏丹草个体在特定刈割管理条件下及补偿生长期(图2、图3)内达到最大高度的潜力b均呈显著负相关(r<-0.87,P<0.05)。
将x取值为1分析海狮苏丹草植株生长高度y′随时间变量x′变化的个体回归方程y′=ax′+b在群体生长高度y随时间变量x变化的回归方程y=ax+b两侧的分布情况,结果显示,海狮苏丹草个体株高生长曲线y′=ax′+b在群体株高生长曲线y=ax+b两侧的分布比率为0.478,这一结果显示出通过平衡自身生长速率与生长潜力的关系适应单一植物群落竞争的海狮苏丹草个体在群体内有近似于均匀分布的格局。
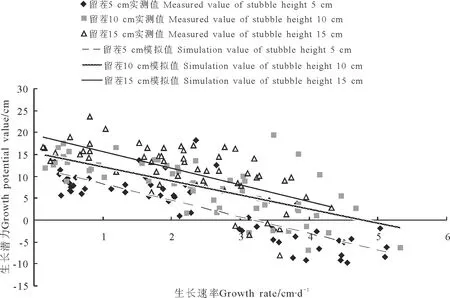
图2 不同刈割处理海狮苏丹草株高平均生长积累速率(a)与积累潜力(b)的关系Fig.2 Relationship between average growth rate (a) and growth potential value (b) of Sorghum sudanense (Hay-King) under different clipping treatments

图3 不同补偿期海狮苏丹草株高平均生长积累速率(a)与积累潜力(b)的关系Fig.3 Relationship between average growth rate (a) and growth potential value (b) of Sorghum sudanense (Hay-King) in different compensatory periods
3 讨论与结论
本研究证明,海狮苏丹草地单作群落的总生物量在刈割条件下有达到相对稳定的趋势,但是在第4补偿生长期产量急剧下降,可能是由于刈割强度过大或受物候期的影响。海狮苏丹草留茬高度为10 cm可获得较高生物量,这一结论与刘景辉等[28]、沈艳等[29]相关研究的结论接近;但不同留茬高度对海狮苏丹草总生物量的影响效果不显著(P>0.05),一方面可能是试验地气候、土壤肥力等因素的差异造成的,因为植物的生理周期及物候特性极易受到干扰;另一方面,本试验在去除了海狮苏丹草地处于生物量急剧退化的第4刈割补偿期生物量的影响后,留茬高度为10 cm可获得显著的累积生物量,即在特定时期对草地植物进行合理的刈割管理的确有助于提高单作海狮苏丹草地生产力。
海狮苏丹草个体平均高度与群落总生物量呈显著正相关,这与Cox[30]关于禾本科牧草群落个体主要通过纵向生长影响群体地上生物量的形成理论一致。海狮苏丹草单株平均分蘖数与群落总地上生物量呈显著负相关,但在去除植株高度的影响后,海狮苏丹草单株分蘖数与群落总生物量的相关不显著。初步分析,可能由于植物均具有在分蘖密度和分蘖大小,即个体大小和个体数量之间的权衡,植物的生物量由个体大小或个体数量或二者共同决定,而本试验仅研究了单株的分蘖数而不是分蘖密度或单位面积的分蘖数,因而不能准确分析出海狮苏丹草个体分蘖数与生物量之间的相关性。
生物体达到稳态状态的能力在很大程度上与该物种的适应能力及生态对策有关[10-12]。试验建立的Linear-Model分析模型结果显示:海狮苏丹草个体株高的平均生长积累速率a与海狮苏丹草个体在特定刈割管理条件下及不同补偿生长期内达到最大高度的潜力b呈显著负相关,且在不同时期,这些个体在群落中的分配大致为1∶1。由于单一植物群落的总生物量在一定范围内达到相对稳定的状态,这种负相关的存在及均匀分布的情况证明,单一植物群落个体可通过调剂自身生长速率与生长潜力的关系适应单一植物群落的生存环境,并使群落总生物量在一定范围内维持相对稳定。
综上所述,单一植物群落个体通过平衡自身生长速率与生长潜力的关系适应群体的生存环境,是实现单作植物群落总生物量在一定范围内维持相对稳定状态的策略之一。
[1] 徐玉鹏,武之新,赵忠祥.苏丹草的适应性及在我国农牧业生产中的发展前景[J].草业科学,2003,20(7):23-25.
[2] 赵全仁,徐廷荣,邢梅兰.美国苏丹草引种比较试验[J].草与畜杂志,1997(3):57-60.
[3] 梁其建.苏丹草在景泰灌区盐碱地的适应性观察[J].草与畜杂志,1998(3):27-31.
[4] 奎嘉祥,钟声,匡崇义.云南牧草品种与资源[M].昆明:云南科技出版社,2003:45-47.
[5] 陈艳瑞.苏丹草在塔里木河下游地区的栽培试验[J].干旱区研究,2004(4):379-383.
[6] Easty D B,Blaedel W J,Anderson L.Continuous electrochemical determination of cyanide.Application to cyanogenic glycosides in Sudan grass[J].Analytical Chemistry,1971,43(4):509-514.
[7] 詹秋文.高粱与苏丹草的遗传及其杂种优势利用的研究[D].南京:南京农业大学,2007:4-7.
[8] 詹秋文,林平,李军.高梁-苏丹草杂交种研究与利用前景[J].草业学报,2001,10(2):56-61.
[9] Wu H,Tang S,Zhang X,etal.Using elevated CO2to increase the biomass of aSorghumvulgare×Sorghumvulgarevar.sudanense hybrid andTrifoliumpratenseL.and to trigger hyperaccumulation of cesium[J].Journal of Hazardous Materials,2009,170(2-3):861-870.
[10] Kooijman S.The stoichiometry of animal energetics[J].Journal of Theoretical Biology,1995,177:139-149.
[11] Frost P C,Evans-White M A,Finkel Z V,etal.Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world[J].Oikos,2005,109:18-28.
[12] Jeyasingh P D,Weider L J,Sterner R W.Genetically-based trade-offs in response to stoichiometric food quality influence competition in a keystone aquatic herbivore[J].Ecology Letters,2009,12:1229-1237.
[13] Makino W,Cotner J B,Sterner R W,etal.Are bacteria more like plants or animals? Growth rate and resource dependence of bacterial C∶N∶P stoichiometry[J].Functional Ecology,2003,17:121-130.
[14] Levi M P,Cowling E B.Role of nitrogen in wood deterioration Ⅶ.Physiological adaptation of wood-destroying and other fungi to substrates deficient in nitrogen[J].Phytopathology,1969,59:460-468.
[15] Andersen T,Hessen D O.Carbon,nitrogen,and phosphorus content of freshwater zooplankton[J].Limnology and Oceanography,1991,36:807-814.
[16] Rhee G Y.Effects of N∶P atomic ratios and nitrate limitation on algal growth,cell composition and nitrate uptake[J].Limnology and Oceanography,1978,23:10-25.
[17] Yu Q,Elser J J,He N,etal.Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland[J].Oecologia,2011,166:1-10.
[18] Yu Q,Chen Q S,Elser J J,etal.Linking stoichiometric homoeostasis with ecosystem structure,functioning and stability[J].Ecology Letters,2010,13:1390-1399.
[19] Diaz S, Cabido M.Vive la difference:Plant functional diversity matters to ecosystem processes[J].Trends in Ecology and Evolution,2001,16:646-655.
[20] Loreau M,Hector A.Partitioning selection and complementarity in biodiversity experiment[J].Nature,2001,412:72-76.
[21] O’Dell R E,Young S L,Claassen V P.Native roadside perennial grasses persist a decade after planting in the Sacramento Valley[J].California Agriculture,2007,61(2):79-84.
[22] Darwin C.The origin of species by means of natural selection[M].London:John Murray,1859:381-412.
[23] 朱珏,张彬,谭支良,等.刈割对牧草生物量和品质影响的研究进展[J].草业科学,2009,26(2):80-85.
[24] 覃宗泉,雷会义,娄秀伟,等.刈割次数对热性草丛草地地面植被的影响[J].草业科学,2010,27(1):103-108.
[25] Harper J L.Population biology of plants[M].New York:Academic Press,1977,44:421-425.
[26] 黎磊,周道玮.红葱种群地上和地下构件的密度制约调节[J].植物生态学报,2011,35(3):284-293.
[27] Yoda K,Kira T,Ogawa H,etal.Self-thinning in overcrowded pure stand under cultivated and natural conditions[J].Journal of Biology of Osaka City University,1963,14:107-129.
[28] 刘景辉,赵宝平,焦立新,等.刈割次数与留茬高度对内农一号苏丹草产草量和品质的影响[J].草地学报,2005,13(2):93-96.
[29] 沈艳,杨鹏,覃强.不同刈割处理对苏丹草生产性能的影响[J].草原与草坪,2009(2):25-28.
[30] Cox C B.Vertebrate palaeodistributional patterns and continental drift[J].Journal of Biogeography,1974,1:75-94.
猜你喜欢
宝藏(2022年10期)2022-03-06 07:25:00
幼儿画刊(2021年10期)2021-10-20 01:44:40
趣味(语文)(2019年10期)2020-01-14 03:26:02
环球时报(2019-04-12)2019-04-12 08:18:57
娃娃乐园·综合智能(2018年11期)2018-07-10 09:24:32
农家科技中旬版(2018年2期)2018-05-05 10:09:54
环球人物(2018年7期)2018-04-23 07:58:42
现代农业(2018年7期)2018-02-18 10:58:09
小星星·阅读100分(低年级)(2016年11期)2016-11-18 22:27:00
新农村(2015年10期)2015-09-28 00:37:29
