
米氏冰草种群种子产量与种子雨的时间动态
2012-03-13 05:37:24林海俊王宏光金晓明刘及东闫志勇
草业科学 2012年11期
林海俊,王宏光,金晓明,刘及东,闫志勇
(1.呼伦贝尔市教育局,内蒙古 呼伦贝尔 021008; 2.呼伦贝尔学院生命科学与化学学院,内蒙古 呼伦贝尔 021008)
种子或果实靠自身的重力和外界力量(如风力等)散布到地表的过程,被形象地称为种子雨(Seed Rain)[1]。以往对种子雨的研究多集中在木本植物[2-6],而对草本植物种子雨的研究相对较少,沙地植物种子雨的研究则更少[7-11]。种子雨是土壤种子库的主要来源,直接影响种群和群落的组成、结构和动态[12],是群落更新发展的重要环节。了解植物种子雨的时间格局,对于深入研究植物种群的更新、发展动态及植被恢复具有重要科学价值。国内外有关种子雨散布时空动态的研究已有大量报道[13-19]。米氏冰草(Agropyronmichnoi)别名为根茎冰草,属禾本科小麦族的多年生草本植物,多分布于欧亚大陆温带草原区,是呼伦贝尔草地的乡土种,具有很高的饲用价值。另外,在长期的自然选择过程中,米氏冰草形成了对沙地环境较强的适应性,既可以成为沙地植被次生演替的先锋种,又可以通过无性繁殖形成沙地植物群落的优势种。米氏冰草具有很强的抗旱性,在年降水量为200~380 mm地区生长良好。因此,在沙漠化治理实践中,米氏冰草是兼顾生态效益和经济效益的重要固沙植物。然而,有关米氏冰草在沙地生境中有性繁殖体散布特征及规律方面的研究鲜见报道。本研究以呼伦贝尔沙漠化草地米氏冰草种群为对象,开展种子雨时空动态及有性繁殖器官特性的研究,旨在为沙化草地植被的恢复和管理提供科学依据。
1 材料与方法
1.1研究区概况 研究区位于内蒙古呼伦贝尔市陈巴尔虎旗完工镇境内,地理位置为11°44′75″~118°54′76″ E,49°08′87″~49°11′48″ N,海拔588.4~618.0 m,具温带大陆干旱半干旱过渡气候特征。年均气温-1.5 ℃,降水量230~300 mm(70%集中于夏秋季),而蒸发量为降水量的4~6倍;年均风速4.5 m·s-1,最大风速为20 m·s-1,全年8级以上大风日数平均30 d以上。地带性植被以贝加尔针茅(Stipabaicalensis)和羊草(Leymuschinensis)为优势种群;沙漠化草地的植被主要以草本植物种为主,其中有米氏冰草、东北牡蒿(Artemisiamanshurica)、扁蓿豆(Melissitusruthenius)、狗尾草(Setariaviridis)、百里香(Thymusvulgaris)、猪毛菜(Salsolafoliosa)、沙蓬(Agriophyllumsquarrosum)等。群落中还零星分布着少量的灌木,如差不嘎蒿(A.halodendron)和小叶锦鸡儿(Caraganamicrophylla)[20]等。
研究样地在呼伦贝尔市陈巴尔虎旗呼和诺尔苏木境内的沙漠化草地内,为2005年进行人工补播的米氏冰草草地,经过了5年的自然恢复。其中,样地1(以下简称半固定沙地)是以米氏冰草为单优势种群的半固定沙地,位于沙丘上部,植被盖度为35%~45%;样地2(以下简称固定沙地)是以米氏冰草为单优势种群的固定沙地,位于沙丘下部,植被盖度为65%~75%。两种样地的面积均大于2 000 m2。
1.2研究方法 种子产量用单位面积内植物有性繁殖体所形成的种子数量来表示;种子雨强度用植物有性繁殖后代散布到周边环境中的单位面积内的数量来表示。试验期间每天的气候因子的测定数据由呼伦贝尔市陈巴尔虎旗气象局提供(表1)。
1.2.1种子雨强度 米氏冰草种子雨的采集时间选择在其成熟期进行,即2009年8月2日-9月27日。每隔5 d收集一次种子。种子雨收集器为直径0.2 m、高0.08 m、底部具铁纱网、上方无封闭的圆筒型竹箩筐(箩筐的底面积为0.031 4 m2)。将种子收集器随机摆放在半固定沙地和固定沙地样地内,每个样地10个重复,共20个种子收集器,在种子收集器外围用一次性筷子将其固定。每次取种子时,先将收集器内的枯枝落叶和其他植物的种子等杂物分离出去,收集近5 d内的米氏冰草种子,放入塑封袋中带回实验室并进行计数。
种子雨强度=种子雨数量/收集器底部面积。
1.2.2有性繁殖体性状及种子产量 在半固定沙地和固定沙地样地分别按草方格大小(草方格为用稻草建植的网格状人工沙障,由于地形和人工作业的差异,草方格面积存在一定误差,一般为1.0~2.5 m2),随机选取10个面积不等的样方,计数每个样方内出现的穗数。同时在半固定沙地和固定沙地样地分别随机抽取30个穗部,计数每穗上的种子数量。同时对随机抽取的穗部,测定其穗长、穗宽、每穗上的小穗数量等。然后计算出种子产量,即:
种子产量=样方内出现的穗数量×平均每穗上的种子数量/样方面积×100%。
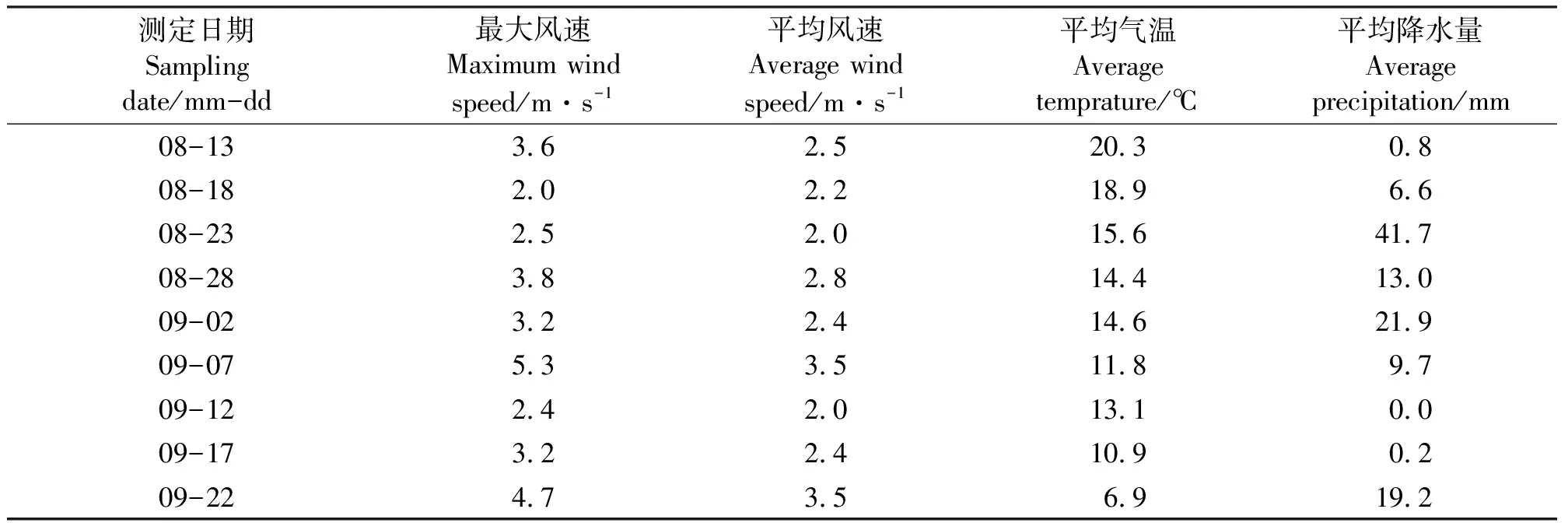
表1 米氏冰草种子雨期间的气候因子数据Table 1 Data of climatic factors during seed rain of Agropyron michnoi
1.2.3数据统计与分析 用Excel和SPSS软件对数据进行统计分析,用平均数(M)代表样本各性状整体水平特征,用标准差(SD)反映样本的变异度,对不同样地的相应指标进行方差分析(One-Way Anova),比较不同生境之间的种子产量及有性繁殖体的差异显著性。
种子雨期间,按每隔5 d的取样日期,分别取这5 d的平均风速、最大风速、平均气温和平均降水量为变量,与种子雨强度进行相关性分析,再以其中相关性最大的参数为变化,与种子雨强度进行回归分析。
2 结果与分析
2.1种子雨时间动态 研究结果显示(图1),半固定沙地和固定沙地的米氏冰草种群种子散布期基本一致,均为8月8日-9月27日。在种子散布期间,半固定沙地和固定沙地的米氏冰草种群的种子雨强度的波动变化也基本相似。种子雨的最大高峰期均出现在9月7日。另外,在8月13日、8月28日和9月22日时,半固定沙地出现了3次小高峰值,而固定沙地只呈小幅度的波动。各个时期的米氏冰草种子雨强度,基本上为半固定沙地高于固定沙地。在半固定沙地和固定沙地,米氏冰草种子雨总强度分别为3 158.2和1 146.5粒·m-2,半固定沙地的种子雨总强度是固定沙地的2.8倍。
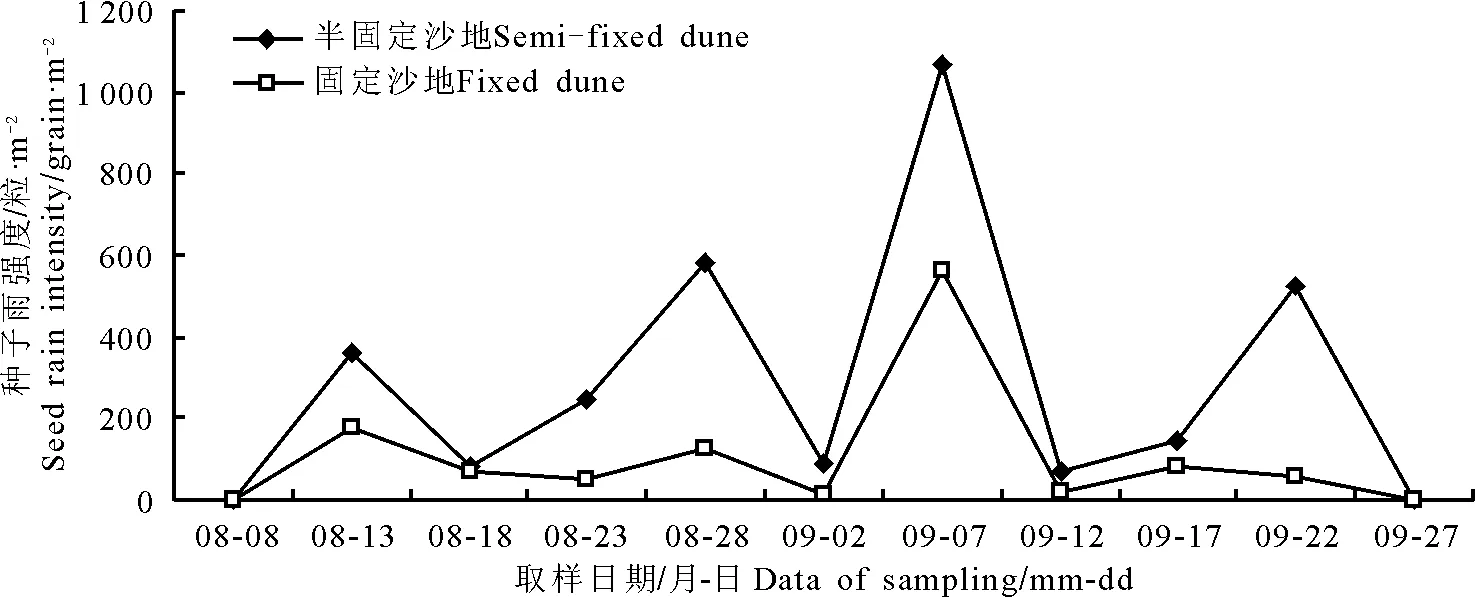
图1 不同生境米氏冰草种群种子雨时间动态Fig.1 Temporal dynamics of seed rain of Agropyron michnoi populations in different habitats
2.2种子产量与有性繁殖体的关系 对米氏冰草种群有性繁殖体的研究显示(表2),其穗长、穗宽、单穗质量及每穗上的小穗数半固定沙地上分别是固定沙地的1.3倍、1.5倍、1.5倍和1.2倍,且均表现出显著差异(P<0.05或P<0.001),表明沙地生境的变化对米氏冰草穗部形态特征产生一定影响。米氏冰草的种子百粒重为半固定沙地比固定沙地的高1.17倍,且存在显著差异(P<0.05),而每穗上的种子数及单位面积种子产量,在两样地之间却无显著差异,表明半固定沙地的米氏冰草种子比固定沙地的大,但在单株种子数量及单位面积种子数量上均相同。
在半固定沙地和固定沙地,米氏冰草种群种子产量分别为10 961和11 502粒·m-2(表2)。其中,在半固定沙地和固定沙地,米氏冰草种子雨数量分别占其种子产量的29%和10%,表明米氏冰草种子成熟后,种子雨进入土壤种子库的数量因生境的变化而有所差异。
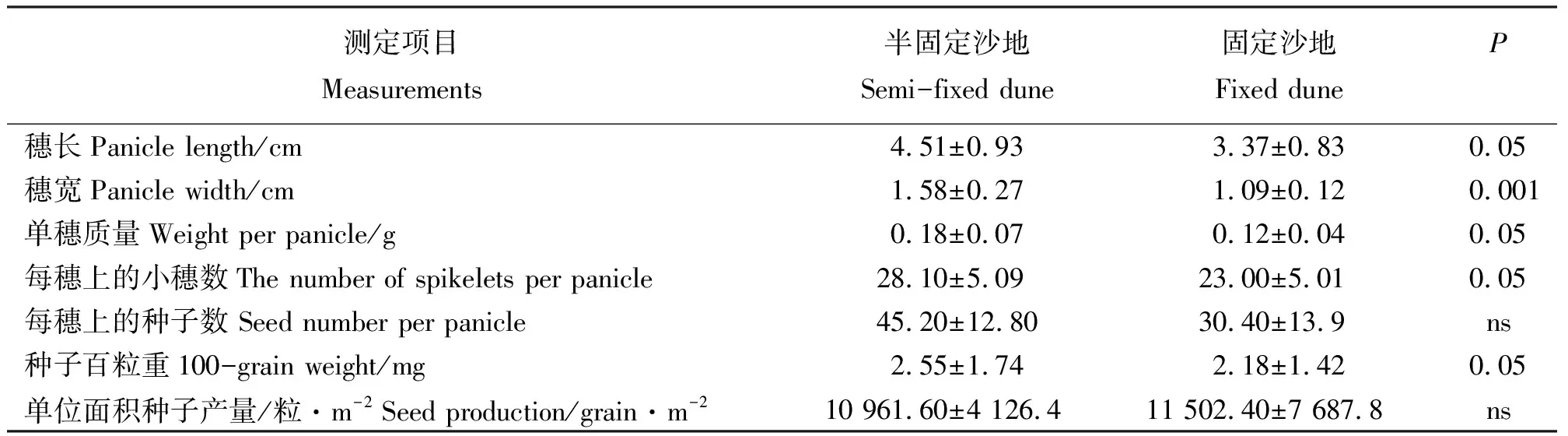
表2 不同生境米氏冰草有性繁殖体特征Table 2 Characteristics of sexual propagules of Agropyron michnoi populations in different habitats
2.3种子雨强度与最大风速的关系 以种子雨期间每5 d的最大风速为变量,与种子雨强度进行回归分析(图2),半固定沙地种子雨强度与风速之间以Y=-572.7+270.8X规律变化,固定沙地则以Y=-253.7+111.7X规律变化,且其回归方程均呈现出正态分布,显著性差异均小于0.001。
3 讨论与结论
种子扩散指种子离开母株的运动,是种子寻求萌发的最佳时间和空间的过程[14]。由于种子通过扩散可以减少幼苗与母株的竞争、幼苗之间的竞争以及母株附近的密度制约性死亡。远离母株的种子与母株附近的种子相比,出苗成活率较高[6,21]。因此,米氏冰草种子雨强度的大小及其分布格局对其在沙地的成功定居具有重要意义。种子扩散是发生在多个空间和时间尺度上的最基本的生物过程,包括种子雨的扩散及种子雨落地后的第2次扩散。以往的研究[22]表明,植物种之间的种子产量千差万别,不同种群之间的种子雨密度相差较大;由于环境条件如土壤深度、地形、土壤养分等的差异,相同植物种群内部不同个体之间的种子雨也有差异。本研究表明,在呼伦贝尔沙化草地,不同生境对米氏冰草种群的种子雨散布时期无影响,但却对其种子雨强度有显著影响。前者可能是由于两样地距离较近,气候条件基本相似所致,但较大尺度上的生境变化是否对米氏冰草的种子雨散布时期有影响,尚不清楚;后者可能是两样地的土壤养分和水分条件存在一定差异所致。但米氏冰草采取何种调整有性繁殖分配的策略,是否与生境土壤养分和水分条件的变化有关,还有待于进一步研究。
植物繁殖分配与植物的生长、植物的生活史、植物所处的演替状态以及植物的生存环境均有关系[23-27]。米氏冰草是无性系植物,既可通过地下根茎的生长进行无性繁殖,也可通过形成有性繁殖体进行种子繁殖,其穗部为有性繁殖器官[25]。
许多干旱地区的植物,如四合木(Tetraenamongolia)种群在环境恶劣的滩地,其生殖值、地上年净生物量的生殖分配比生境条件好的丘地要高[28]。本研究结果也与此相似,即米氏冰草种群的穗长、穗宽、单穗质量、每穗上的小穗数及种子百粒重等指标,在环境条件相对恶劣的半固定沙地均显著高于固定沙地。另外,质量较大的种子更有利于埋入土壤,有效地获取土壤中的水分,促进其萌发和幼苗的生长。
在本研究中,虽然不同生境米氏冰草种子单穗上的种子数量与单位面积的种子产量均无显著差异,但整个种子雨期间收集的累计种子数量只占其种子产量的10%~33%,且半固定沙地比固定沙地高3倍。这可能是因为随着沙地植被的恢复,动物数量也有所增加,致使大部分成熟脱落的种子被动物所采食。Gashwiller[29]发现,鼠类的破坏造成道格拉斯杉树(Pseudotsugamenziesii)种子的损失率达63%,仅有12%的种子在第2年萌发。沈泽昊等[30]对山地常绿落叶阔叶混交林种子雨的地形格局研究发现,地形对植被空间格局异质性的影响机制之一是作用于种子雨的空间分布。在呼伦贝尔沙地,半固定沙地一般位于沙丘的坡顶或坡中,风力较大,环境较干燥,而固定沙地位于沙丘的坡下,风力小,环境相对不算太干燥。这可能是半固定沙地的种子比固定沙地的种子更易散布的原因之一。
因此,沙地微生境的变化对米氏冰草种子雨散布的时期及波动规律无影响,而对其有性繁殖体的形态特征具有显著影响。
[1] Harper J L.Population Biology of Plants[M].London: Academic Press,1977:124-201.
[2] 马万里,荆涛,Kujansuu J,等.长白山地区胡桃楸种群的种子雨和种子库动态[J].北京林业大学学报,2001,23(3):70-72.
[3] 马杰,李庆芬,孙儒泳.啮齿动物和鸟类对东灵山地区辽东栎种子丢失的影响[J].生态学杂志,2004,23(1):107-110.
[4] 路纪琪,李宏俊,张知彬.啮齿动物分散贮食的影响因素[J].生态学杂志,2005,24:528-532.
[5] 肖治术,张知彬,王玉山.以种子为繁殖体的植物更新模型研究[J].生态学杂志,2003,22(4):70-75.
[6] Connell J H.On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees[A].In:Boer P J,Gradwell G R.Dynamics of Populations[M].Wageningen:Centre for Agricultural Publishing and Documentation,1971:298-312.
[7] Rabinowicz D,Rapp J K.Seed rain in a north American tall grass prairie:The resemblance of their abundance and compositions to dispersing seeds[J].Oikos,1980,36:191-195.
[8] Schott G W,Hamburg S P.The seed rain and seed banks of an adjacent tallgrass prairie old fields[J].Canadian Journal of Botany,1997,75:1-7.
[9] Watkinson A T,Harper J L.The demography of a sand dune annual,Vulpiafasciculate.Ⅲ.The dispersal of seeds[J].Journal of Ecology,1978,66:483-498.
[10] Westelaken I L,Maun M N.Spatial pattern and seed dispersal ofLithospermumcarolinienseon Lake Huron sand dunes[J].Canadian Journal of Botany,1985,63:125-132.
[11] Owen N W,Kent M,Dale M P.Spatial and temporal variability in seed dynamics of machair sand dune communities,Outer Hebriges,Scottland[J].Journal of Biogeograph,2001,28:565-588.
[12] Clobert J,Danchin E,Dhondt A A.Dispersal[M].Oxford:Oxford University Press,2001:103-132.
[13] Loiselle B A,Ribbens E,Vargas-Ramírez O R.Spatial and temporal variation of seed rain in a tropical lowland wet forest[J].Biotropica,1996,28(1):82-95.
[14] 班勇.植物生活史对策的进化[J].生态学杂志,1995,14(3):33-39.
[15] Barnes E M.Seed predation,germination and seedling establishment ofAcaciaeriolobain northern Botswana[J].Journal Arid of Environments,2001,49:541-554.
[16] Chambers J C.A day in the life of a seed: Movements and fates of seeds and their implications for natural and managed systems[J].Annual Review of Ecology and Systematics,1994,25:263-293.
[17] 杨允菲,祝廷成.松嫩平原大针茅群落种子雨动态的研究[J].植物生态学与地植物学学报,1991,15:46-55.
[18] 邹春静,徐文铎,刘广田.沙地云杉种群种子雨的时空分布规律[J].生态学杂志,1998,17(3):16-19.
[19] 胡星明,蔡永立,李恺.浙江天童常绿阔叶林栲树种子雨的时空格局[J].应用生态学报,2005,16(5):815-819.
[20] 金晓明,艾琳,卢欣石,等.不同群落中米氏冰草和羊草的年龄结构动态[J].生态学报,2011,31(18):5406-5413.
[21] Janzen D H.Herbivores and the number of tree species in tropical forests[J].American Naturalist,1970,104:501-528.
[22] Urbanska K W,Fattorini M.Seed rain in high altitude restoration plots in Switzerland[J].Restoration Ecology,2000,8:74-79.
[23] 钟章成.植物种群的繁殖对策[J].生态学杂志,1995,14(1):37-42.
[24] 操国兴,谢德体,钟章成.植物种群的生殖分配[J].四川林业科技,2003,24(2):25-29.
[25] 金晓明.米氏冰草繁殖生态学特征及种群竞争机制[D].北京:北京林业大学,2011:36-46.
[26] 黄世华,任媛媛,张世挺.根竞争对窄叶野豌豆生长的影响:公共的悲剧[J].草业科学,2011,28(2):266-271.
[27] 武建双,沈振西,张宪洲,等.藏北高原人工垂穗披碱草种群生物量分配对施氮处理的响应[J].草业学报,2009,18(6):113-121.
[28] 徐庆,刘世荣,臧润国.中国特有植物四合木种群的生殖生态特征—种群生殖值及生殖分配研究[J].林业科学,2001,37(2):36-41.
[29] Gashwiller J S.Conifer seed survival in Western Oregon clearcuts[J].Ecology,1967,48(3):431-438.
[30] 沈泽昊,吕楠,赵俊.山地常绿落叶阔叶混交林种子雨的地形格局[J].生态学报,2004,24(9):1981-1987.
猜你喜欢
莫愁(2023年25期)2023-09-14 06:34:56
海洋与湖沼(2022年2期)2022-03-25 10:32:14
现代苏州(2022年5期)2022-03-21 22:57:09
生态科学(2019年3期)2019-07-02 07:33:14
渔业研究(2017年4期)2017-08-30 13:34:36
新疆农业科学(2016年6期)2016-07-23 01:51:10
IT时代周刊(2015年9期)2015-11-11 05:51:51
海洋科学(2015年9期)2015-03-17 05:49:25
少儿科学周刊·儿童版(2014年1期)2014-03-10 06:23:18
草食家畜(2012年2期)2012-03-20 13:22:39
