
木质层孔菌产锰过氧化物酶条件的优化及酶学性质研究
2012-02-09 07:18:34崔艳红韩庆功常魁珍魏龙龙张贝贝胡志明
饲料工业 2012年12期
崔艳红 韩庆功 常魁珍 魏龙龙 张贝贝 胡志明
(河南科技学院动物科学学院,河南新乡453003)
木质素是一类结构复杂且稳定的生物大分子物质,在秸秆类物质中约占20%,其产量仅次于纤维素。由于动物体内缺乏降解木质素的酶类,因此很难将其作为有效的饲料来源。自然界中,木质素的完全降解是真菌、细菌、放线菌及相应微生物群落共同作用的结果,其中真菌起着主导作用。赵红霞等(2002)[1]报道,在适宜的条件下,白腐真菌的菌丝首先利用其分泌的超纤维素酶溶解表面的蜡质,然后菌丝进入秸秆内部,并产生锰过氧化物酶(MnP)、木质素过氧化物酶(LiP)、纤维素酶、半纤维素酶,降解秸秆中的木质素和纤维素,其中起关键作用的是LiP和MnP。目前,关于木质素降解酶的研究主要集中于白腐真菌,其对木质素的降解主要通过分泌胞外降解酶完成,主要有3种:①木质素过氧化物酶(Lignin peroxidase,LiP);②锰过氧化物酶(Manganese peroxidase,MnP);③漆酶(1accase,Lac)[2]。
木质层孔菌(F.lignosus)是白腐真菌的一种,产生的胞外酶有木质素过氧化物酶(LiP)、锰过氧化物酶(MnP)、木聚糖酶、β-葡聚糖酶、漆酶等氧化酶类。Pease等[3]报道,锰过氧化物酶对木质素的降解起关键作用。据调查,目前在研究木质素降解酶的领域对锰过氧化物酶研究尚少,而且大多集中在黄孢原毛平革菌和变色栓菌中的锰过氧化物酶。董旭杰等(2006)[4]研究指出,将3种白腐菌产锰过氧化物酶的能力进行比较,发现木质层孔菌产锰过氧化物酶量最高,可达880.2 U/l。
MnP是一种含铁血红素的糖基化过氧化物酶,在Mn2+和H2O2存在时,MnP能氧化分解芳香环多聚体,被认为是木质素降解的关键酶之一。相对于其它过氧化物酶,MnP的特别之处在于它的底物是有机酸。其可以氧化Mn2+为Mn3+,而Mn3+可以鳌合有机酸,这种鳌合物可通过扩散离开酶活性中心,去催化氧化二级底物,如酚化合物、木质素相关复合物及大分子量氯代木质素、氯代酚和许多其它物质。其对木质素的降解有赖于大量稳定的Mn3+的存在,与其他酶一起能把木质素彻底降解为CO2。因为MnP的底物专一性弱,具有氧化降解各类芳香族化合物的独特能力,因此,在纸浆的生物漂白、有机污染物的降解、染料的脱色、褐煤的生物降解、农业废弃物的处理和催化聚合反应等方面得到了较多的研究与应用[5]。
本试验选用白腐真菌中能产生较高锰过氧化物酶的菌株木质层孔菌5.132,研究了不同温度、碳源、氮源和培养时间对菌株5.132产锰过氧化物酶的影响,以确定其最佳培养条件,并对锰过氧化物酶的部分酶学性质进行了研究,为锰过氧化物酶的高效利用奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌种
木质层孔菌[Fomes lignosus(Berk)Coke],购自中国科学院微生物研究所,河南科技学院动物科学学院营养教研室保存。
1.1.2 培养基
PDA固体培养基:马铃薯提取液1 000 ml、葡萄糖20.0 g、琼脂15.0 g;
PDA液体培养基:马铃薯提取液1 000 ml、葡萄糖20.0 g。
1.1.3 仪器设备
电热蒸馏水器(北京市光明医疗仪器厂),数显超级恒温水浴锅(金坛市瑞尔电器有限公司),电热恒温培养箱(上海实验仪器厂有限公司),SW-CJ-2FD型双人单面净化工作台(苏州净化设备有限公司),722型光栅分光光度计(山东高密分析仪器厂),分析天平AB240-N(上海梅特勒-托利多仪器有限公司),电热鼓风干燥箱(上海实验仪器有限公司),802-型离心机(江苏金坛大地自动化仪器厂)。
1.2 方法
1.2.1 孢子悬液的制备
将木质层孔菌接种于灭菌后的斜面PDA固体培养基上活化,向培养5 d的斜面试管内加入无菌水,将菌丝洗出到三角瓶内,加玻璃珠打碎后,进行孢子计数。然后接入到250 ml三角瓶中,三角瓶装液量50 ml、接种量0.2 ml(以孢子计1.0×106个/ml)。
1.2.2 粗酶液制备
取木质层孔菌液体培养物于离心管中,3 000 r/min离心10 min,上清液即为粗酶液。
1.2.3 锰过氧化物酶活力测定
Mn3+是MnP作用的直接产物,在238 nm处测Mn3+的吸光值是测定MnP活力的一个较为灵敏简便的方法,此方法称乳酸钠法。乳酸钠法还可用于连续测定,具体步骤为:在4 ml反应体系中含50 mmol/l(pH值4~5)的乳酸钠缓冲液3.4 ml、1.6 mmol/l的硫酸锰溶液0.1 ml、粗酶液0.4 ml,预热至37℃时,加1.6 mmol/l的H2O2溶液0.1 ml启动反应。在238 nm紫外光处,测定反应4 min内的吸光光度值(OD)。在对照组中,以煮沸灭活15 min酶液代替原酶液,以蒸馏水代替H2O2溶液,其他反应物不变。锰过氧化物酶活力定义为每分钟使1 μmo1/l的Mn2+转化为Mn3+所需的酶量为1个酶活力单位(U/l)。
2 结果与分析
2.1 优化产酶条件的建立
2.1.1 培养温度对木质层孔菌产锰过氧化物酶的影响
将分装好的基础液体培养基高压灭菌后,加入5.132孢子悬液,分别在20、25、30、35、40、45℃的恒温条件下培养,培养5 d后测定酶活,结果如图1所示。从图1中可以看出,木质层孔菌在25~30℃范围内产酶量急剧上升,在30~35℃范围内产酶量增加缓慢并达到高峰为185~215 U/l。35℃时的产酶量是25℃
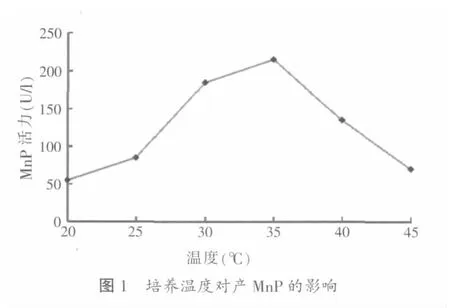
时产酶量的2.5倍,温度高于35℃时产酶量开始下降,到45℃时产酶量为70 U/l,与低温状态下的产酶量几乎无差异。综合比较,35℃为木质层孔菌合成锰过氧化物酶的最佳培养温度,温度过高或过低都不利于酶的合成。
2.1.2 不同碳源对木质层孔菌产锰过氧化物酶的影响
本试验在基础液体培养基的基础上分别选用不同的碳源:葡萄糖(单糖)、蔗糖(二糖)、可溶性淀粉(多糖)、麸皮和小麦秸粉各1.5 g/100 ml,分别接入菌种,静置培养至第5 d测定酶活力,发现酶活力随不同碳源而异,如图2所示。
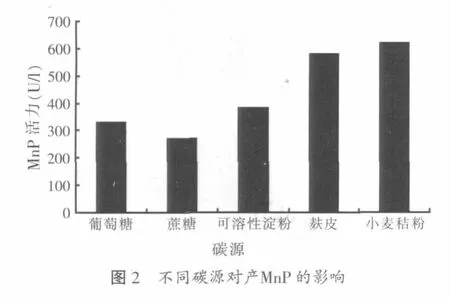
由图2结果可知,麸皮和小麦秸粉最适于木质层孔菌产锰过氧化物酶,酶活力分别高达578.4 U/l和620.7 U/l,产量高于其他碳源。所以,在5种碳源中,对木质层孔菌锰过氧化物酶合成最有利的是小麦秸粉,其次是麦麸,接下来依次为可溶性淀粉、葡萄糖和蔗糖。
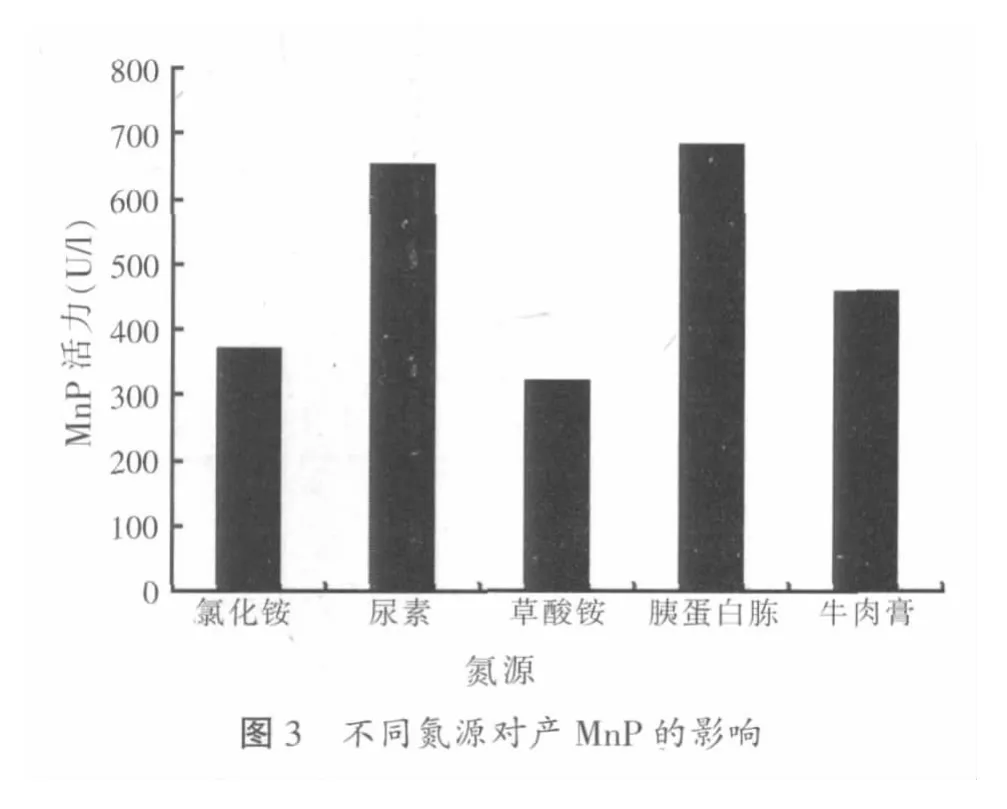
2.1.3 不同氮源对木质层孔菌产锰过氧化物酶的影响
本试验在100 ml PDA基础液体培养基中加小麦秸粉1.5 g做碳源,然后分别加尿素、草酸铵、氯化铵、胰蛋白胨、牛肉膏各0.5 g,接种木质层孔菌,培养5 d后进行酶活力测定,考察不同氮源对木质层孔菌产锰过氧化物酶的影响,结果见图3。
由图3可看出,木质层孔菌在5种不同氮源培养条件下均有锰过氧化物酶的合成,但锰过氧化物酶产酶活力的差异比较大。氮源为胰蛋白胨时,锰过氧化物酶的活力最高,为680.5 U/l;以尿素为氮源时的锰过氧化物酶活力为650.3 U/l,与胰蛋白胨为氮源时相比酶的活力稍微下降。以牛肉膏为氮源时,锰过氧化物酶的活力为458.9 U/l;以氯化铵为氮源时,锰过氧化物酶的活力为369.7 U/l;以草酸铵为氮源时,锰过氧化物酶的活力最低,为320.5 U/l。由此可见,以胰蛋白胨为氮源时的锰过氧化物酶的活力最高,尿素次之,随后为牛肉膏、氯化铵和草酸铵。
2.1.4 培养时间对木质层孔菌产锰过氧化物酶的影响
用基础液体产酶培养基培养木质层孔菌,每隔24 h测酶活力,连测10 d以考察不同培养时间对木质层孔菌合成锰过氧化物酶的影响,结果如图4。
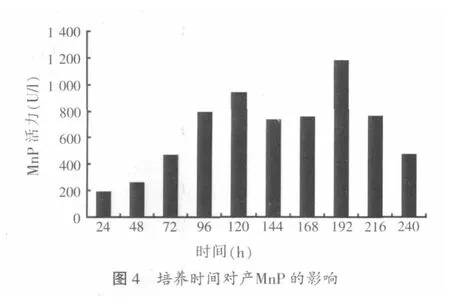
由图4可知,木质层孔菌产锰过氧化物酶的量随时间的延长而平稳上升,在120 h时出现一个产酶高峰,酶活力为938 U/l。此高峰过后,酶活力稍有下降,并在192 h时酶活力突然升高,此阶段为测定期间的酶活最高峰值,酶活力高达1 180.3 U/l,高峰过后锰过氧化物酶的活力大幅度下降。
2.2 酶作用特性的研究
2.2.1 酶作用最适温度
分别将优化条件下提取的粗酶液在32、37、42、47、52、57、62℃进行酶解反应,测定不同温度下锰过氧化物酶的相对活力(原粗酶液相对酶活为100%),结果见图5。
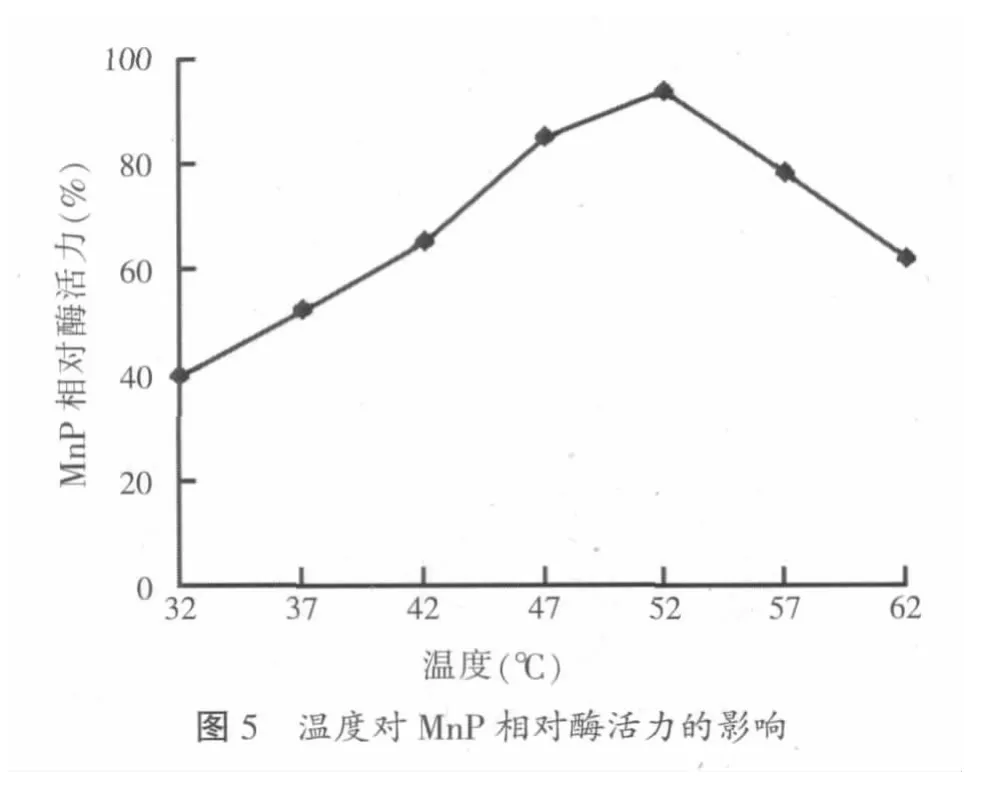
一般而言,在酶没有发生变性之前,其酶活会随反应温度的增加而增强,从图5中的曲线图可以看出,在32~47℃温度范围内,锰过氧化物酶的相对酶活随温度的升高而逐渐增大,在52℃时,相对酶活达到最大,继续升高温度,锰过氧化物酶的相对酶活呈快速下降趋势,说明温度高于52℃时会导致锰过氧化物酶发生部分结构的变性,从而导致相对酶活大大降低。
2.2.2 酶作用最适pH值
真菌在酸性条件下木质素降解酶活性最高,但不同的菌种有不同的最适pH值范围。将提取的粗酶液分别于pH值3.5、4.0、4.5、5.0、5.5的50 mmol/l醋酸-醋酸钠缓冲液中进行酶解反应,测定不同pH值情况下的相对酶活力,结果如图6所示。
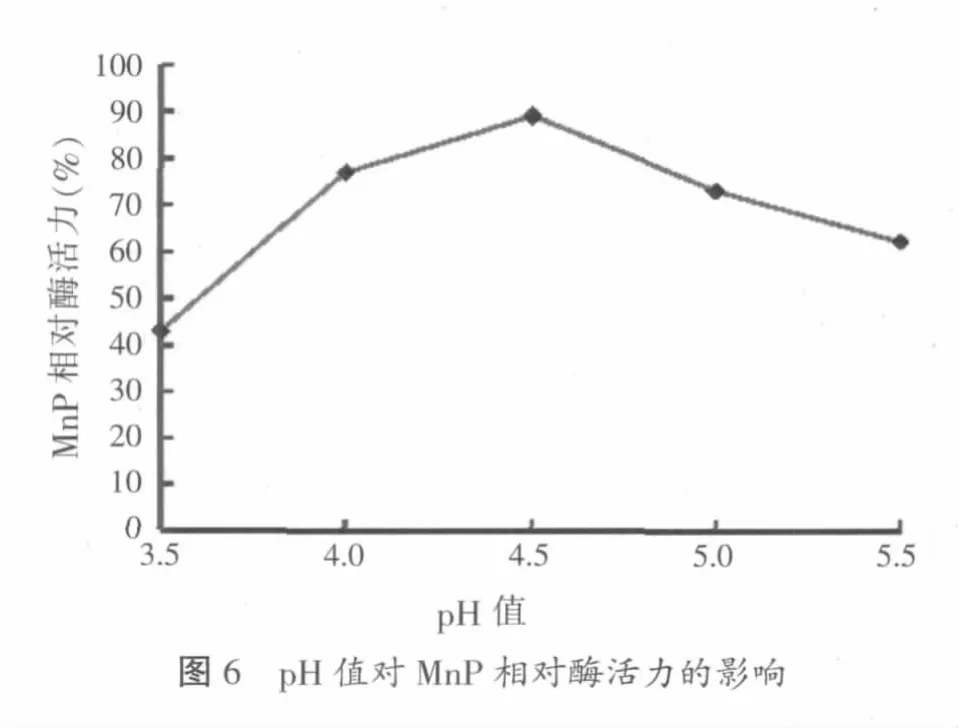
图6结果显示,木质层孔菌产锰过氧化物酶和大多数白腐真菌木质素降解酶系一样,有相对狭窄的最佳pH值范围。锰过氧化物酶在pH值4~5范围内酶活力比较稳定,其相对酶活力均能维持在70%以上,并在pH值4.5时相对酶活力最高,达89%。
2.2.3 金属离子对酶作用特性的影响
本试验在粗酶液中分别加入1 mmol/l的CuSO4·5H2O、MgSO4、FeSO4·7H2O、CaCl2、ZnSO4、AgNO3各1 ml,考察在不同金属离子存在的条件下锰过氧化物酶的相对酶活力,结果如图7所示。
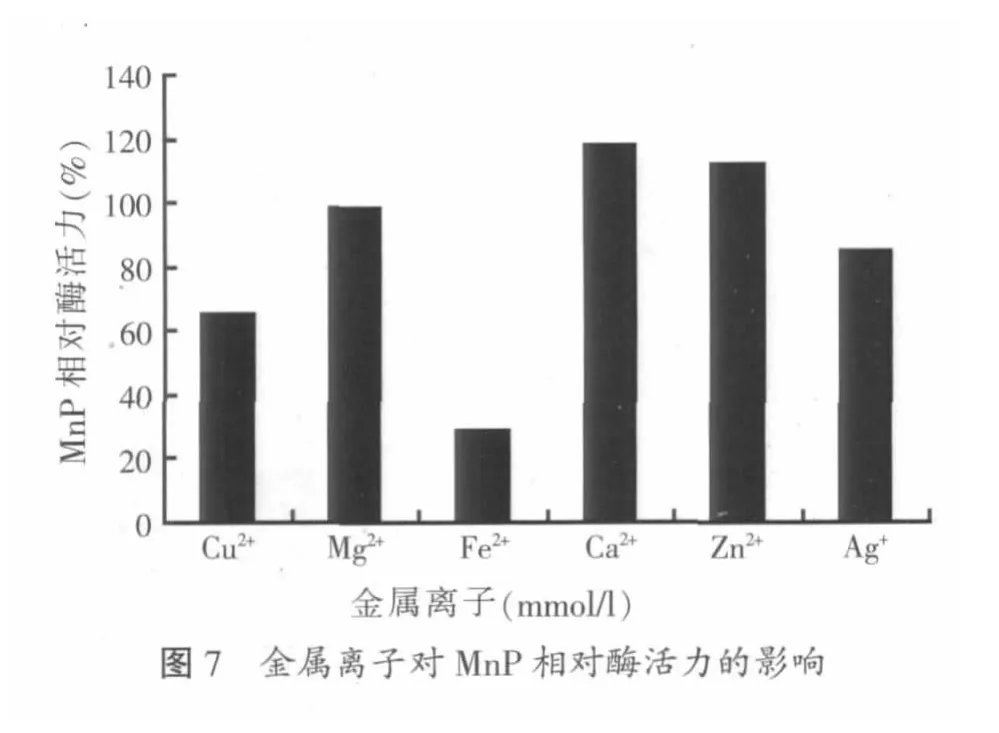
由图7可知,在有Zn2+和Ca2+存在的环境下,锰过氧化物酶的相对酶活力大于100%,其对MnP的活性有促进作用;Mg2+存在的环境中相对酶活达98.7%,对MnP的活力影响不是很大;而在Cu2+、Fe2+和Ag+存在的环境下,可使锰过氧化物酶的相对活力下降,对锰过氧化物酶活力具有抑制作用,其中Fe2+的抑制作用最强。
3 讨论
3.1 产酶条件的优化
碳源是构成菌体碳架及能量的来源,真菌细胞干重的一半是由碳组成的,由此可见,碳源在真菌生长过程中占有很重要的地位。小麦秸粉作为最佳碳源,其锰过氧化物酶的产量与以麸皮为碳源时的产量相差不大,比以可溶性淀粉为碳源时高1.6倍,比以葡萄糖为碳源时高1.8倍,比以蔗糖为碳源时高2.3倍。本试验结果与杨晓宽等(2004)[6]报道的白腐菌产锰过氧化物酶最佳碳源为葡萄糖的试验结果不一致,有待进一步的研究。
氮源是合成菌体蛋白、核酸等含氮物质的重要组成成分。不同氮源对真菌菌丝体的生长有不同程度的促进作用。本试验结果表明,以胰蛋白胨为氮源时的锰过氧化物酶活力最高,尿素次之,随后为牛肉膏、氯化铵和草酸铵。由此可推出有机氮源比无机氮源更有利于锰过氧化物酶的合成,但考虑到蛋白胨实际含氮量不恒定,并且利用尿素代替胰蛋白胨为氮源,可节约生产成本,更有利于工业化生产。此试验结果与董旭杰等[7]报道的木质层孔菌产锰过氧化物酶的氮源优劣顺序相似。
在培养时间对产酶的影响中,试验结果显示,随培养时间延长酶量逐渐提高,培养至第5 d时出现一个产酶高峰,之后有所下降,至第8 d又出现一个产酶高峰,之后酶活力大幅度下降。分析在最初培养基中的营养物质主要用于菌株的生长,所以酶活力比较低,随菌体的生长,其产酶量开始上升,并在第8 d达到最高峰为1 180.3 U/l,之后,菌体生长进入稳定期,随着营养物质的消耗,黏度上升,溶氧不足,酶活开始下降。
3.2 木质层孔菌锰过氧化物酶酶学性质
酶的温度耐受性和其本身的结构特点有很大关系,在不同温度条件下酶反应速率也大不相同。从试验结果和实际生产的角度考虑,52℃为木质层孔菌产锰过氧化物酶的最适反应温度,此结果与吴会广等[8]报道的锰过氧化物酶最适酶解温度为55℃的结论基本相符。
金属离子对酶的影响是多方面的,离子种类、离子电荷数、离子半径大小以及离子浓度等,都会对酶的活性产生不确定的影响。吴会广[8]研究表明,铁离子的浓度越低其抑制作用越小,本试验结果Fe2+的强抑制作用与所用浓度有很大关联。金属离子对酶的作用有两种:一是作用酶的辅助因子,二是作为激活剂起作用,但是金属离子与酶如何相互作用改变生物活性的分子机理仍然有待于进一步研究。
4 结论
由试验结果可知,木质层孔菌产锰过氧化物酶的最佳条件,即培养温度35℃、碳源为小麦秸粉1.5 g/100 ml、氮源为胰蛋白胨0.5 g/100 ml、最佳培养时间为8 d。通过对产酶条件的优化,最高酶活力可达1 180.3 U/l,这比优化前的木质层孔菌锰过氧化物酶产量提高了5.5倍。
酶作用特性研究结果显示,木质层孔菌产锰过氧化物酶的最适反应温度为52℃;最佳pH值范围在4~5之间;在金属离子的影响条件下,Zn2+和Ca2+对MnP的活力有促进作用;Mg2+对MnP的活力影响不是很大;Cu2+、Fe2+和Ag+可使锰过氧化物酶的相对活性下降,具有对锰过氧化物酶活性的抑制作用,其中Fe2+的抑制作用最强。
[1] 赵红霞,杨建军,詹勇.白腐真菌在秸秆作物资源开发中的研究[J].饲料工业,2002,23(11):40-42.
[2] 苏小军,熊兴耀,谭兴和,等.黑曲霉AF-1固态发酵产生淀粉酶的条件优化[J].湖南农业大学学报,2009,35(2):208-212.
[3] Pease E,Tien M.Heterogene and regulation of manganese peroxidases from phanerochaete ehrysos-Porium[J].Baeteriol.,1992,174:3532-3540.
[4] 董旭杰,曹福祥,陈静,等.3种白腐菌木质素降解酶的比较[J].中南林业科技大学学报,2006,27(3):131-135.
[5] 刘梦茹,付时雨,詹怀宇,等.锰过氧化物酶应用的研究进展[J].林产化学与工业,2006,26(20):112-116.
[6] 杨晓宽,杜连祥,路福平.白腐菌产锰过氧化物酶培养基的优化[J].生物技术,2004,14(3):49-50.
[7] 董旭杰,曹福祥,龙绛雪.木质层孔菌产锰过氧化物酶条件的优化[J].江西农业学报,2009,21(3):94-95.
[8] 吴会广.Rhizoctoniasp-SYBC-M3产MnP发酵条件的优化及酶学性质研究[D].江苏无锡:江南大学,2008.
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:24
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:52
河北科技师范学院学报(2022年2期)2022-08-26 08:55:46
中国食用菌(2021年10期)2021-12-01 11:21:10
山东工业技术(2016年15期)2016-12-01 05:31:50
中国塑料(2016年3期)2016-06-15 20:30:00
中国酿造(2016年12期)2016-03-01 03:08:19
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:36
应用海洋学学报(2015年3期)2015-11-22 07:39:30
食品工业科技(2014年13期)2014-03-11 18:16:49
