
Ⅰ型囊泡膜谷氨酸转运体在生后发育大鼠三叉神经运动核中的表达
2012-02-08 06:26庞有旺郑季南洪庆南
中国组织化学与细胞化学杂志 2012年5期
庞有旺 郑季南 洪庆南 方 钧
(解放军泉州第180医院骨科,福建泉州362000)
三叉神经中脑核神经元传递口面部空间位置信息和闭口肌运动信息[1]。三叉神经中脑核向三叉神经运动核的直接投射组成咀嚼运动的单突触反射环路,在调控咀嚼运动中发挥重要作用[2-4]。以往国内、外研究表明大鼠三叉神经运动核内Ⅰ型囊泡膜谷氨酸转运体神经终末为三叉神经中脑核中枢突终末[5-7]。本研究应用免疫组织化学染色技术,在光镜下观察了大鼠三叉神经运动核Ⅰ型囊泡膜谷氨酸转运体阳性纤维终末在生后发育过程中变化。
材料和方法
本实验共用了18只Sprague-Dawley幼年大鼠和3只成年大鼠。把幼鼠出生日作为newborn、然后依次为生后1d、2d等。newborn、生后3、7、10、14、21d和成年每个年龄段各取3只(非同窝)。在戊巴比妥钠(60mg/kg)腹膜腔注射深麻醉下,先用0.025mol/L磷酸缓冲液配制的生理盐水(PBS,pH7.3)经心脏冲洗血液,至流出的液体无血色,再用50-300ml含0.2%多聚甲醛和75% (v/v)饱和苦味酸的0.1mol/L的磷酸缓冲液(pH7.3)灌注固定15-30min。灌毕立即取脑并浸于含2%多聚甲醛和75%饱和苦味酸的0.1mol/L PBS后固定24-48h(4℃),然后移入含25%蔗糖的0.1mol/L的PB中,4℃过夜。冰冻连续冠状切片,片厚40μm,切片分2套,分别收集于0.05mol/L的PBS中。
第1套切片用于VGluT1免疫组织化学染色。具体步骤如下:(1)用兔抗 VGluT1IgG(0.8μg/ml,日本京都大学金子武嗣先生惠赠)孵育切片过夜;(2)生物素标记的驴抗兔IgG(1∶100,Jackson)室温孵育4h;(3)移入 ABC-Elite(1∶100,Vector)孵育切片3h。其中步骤(1)和(2)用含5%驴血清、0.05%叠氮钠、0.5%Triton X-100和0.25%角叉菜胶的0.05mol/L 的 PBS稀 释,步 骤(3)用 含 0.3%Triton X-100的0.05mol/L的PBS稀释。然后进行DAB反应和双氧水呈色。裱片、脱水透明、DPX封片。光镜下观察并拍照。
第2套切片用于对照实验。省略一抗血清或用正常血清替代一抗血清进行对照实验,结果为阴性。
结 果
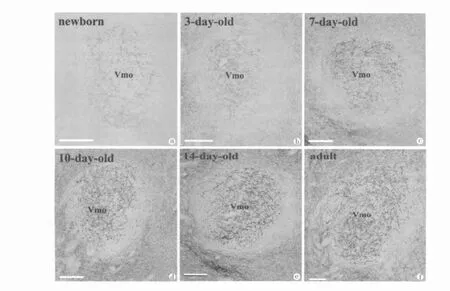
新生幼鼠的三叉神经运动核就可观察到Ⅰ型囊泡膜谷氨酸转运体的表达,呈低强度,分布在背外侧部,此时一个显著的特征是三叉神经运动核周区不能探查到Ⅰ型囊泡膜谷氨酸转运体的阳性结构(图a)。出生后的第一周是幼鼠三叉神经运动核内Ⅰ型囊泡膜谷氨酸转运体表达快速增长期,新生、生后3d、生后7d每阶段都有明显增加(图a-c),生后7d已经接近成年表达水平(图a-f)。生后第二周,幼鼠三叉神经运动核内Ⅰ型囊泡膜谷氨酸转运体表达也有逐渐增加(图d-e),生后14d幼鼠三叉神经运动核内的Ⅰ型囊泡膜谷氨酸转运体表达达到成年模式(图e-f)。
讨 论
三叉神经中脑核中枢突与三叉神经运动核内的运动神经元形成单突触反射通路,该通路以谷氨酸作为递质[8]。作为一种氨基酸,谷氨酸在神经元内既是神经递质,又是中间代谢产物。在神经元内,谷氨酸只有被转运至突触囊泡内才可以发挥神经递质功能。近年已经证明分布在囊泡膜的谷氨酸转运体有三种亚型,负责转运谷氨酸到突触囊泡内[5-7]。在大鼠三叉神经运动核内,Ⅰ型囊泡膜谷氨酸转运体表达于三叉神经中脑核神经元的中枢突[9]。大鼠发育过程中Ⅰ型囊泡膜谷氨酸转运体在三叉神经中脑核神经元的胞体和外周突中的表达变化以往已经进行了研究报道[10]。本文观察了大鼠生后发育过程中三叉神经运动核内Ⅰ型囊泡膜谷氨酸转运体的表达变化。我们观察到Ⅰ型囊泡膜谷氨酸转运体在新生大鼠三叉神经运动核内已经表达,表明胚胎发育过程中已建立了三叉神经中脑核神经元发挥兴奋性突触传递功能。出生后Ⅰ型囊泡膜谷氨酸转运体在三叉神经运动核内表达快速增进,1w内已接近成年水平,2w后达到成年水平,为幼鼠的咀嚼活动奠定了基础。
以往的研究表明三叉神经运动神经元表达多种谷氨酸受体,包括AMPA和NMDA受体,有多种AMPA和NMDA受体亚型在刚出生幼鼠的三叉神经运动神经元表达[11-13]。生理学研究表明单突触反射环路中通过AMPA和NMDA受体引起三叉神经运动核神经元的兴奋[14-15]。显然Ⅰ型囊泡膜谷氨酸转运体在生后大鼠三叉神经运动核内的表达是与受体的表达是匹配的。发育过程中,神经环路的建立依赖神经活动[16],因而尽管哺乳期幼鼠以吮吸、吞咽等活动为主,咀嚼活动较少,Ⅰ型囊泡膜谷氨运体在三叉神经运动核内表达可能也具有重要生理意义,比如参与神经环路的建立。但详细的作用需要进一步研究。
[1]Luo PF,Moritani M,Dessem D.Jaw-Muscle Spindle Afferent Pathways to the Trigeminal Motor Nucleus in the Rat.J Comp Neurol,2001,435:341-353
[2]Nozaki S,Iriki A,Nakamura Y.Localization of central rhythm generator involved in cortically induced rhythmical masticatory jaw-opening movement in the guinea pig.J Neurophysiol,1986,55:806-825
[3]Nakamura Y,Katakura N.Generation of masticatory rhythm in the brainstem.Neurosci Res,1995,23:1-19
[4]Sahara Y,Hashimoto N,Nakamura Y.Hypoglossal premotor neurons in the rostral medullary parvocellular reticular formation participate in cortically-induced rhythmical tongue movements.Neurosci Res,1996,26:119-131
[5]Katakura N,Chandler SH.An iontophoretic analysis of the pharmacologic mechanisms responsible for trigeminal motoneuronal discharge during masticatory-like activity in the guinea pig.J Neurophysiol,1990,63:356-369
[6]Turman JE Jr,Chandler SH.Immunohistochemical localization of glutamate and glutaminase in guinea pig trigeminal premotoneurons.Brain Res,1994,634:49-61
[7]Ni B,Wu X,Yan GM,et al.Regional expression and cellular localization of the Na+-dependent inorganic phosphate cotransporter of rat brain.J Neurosci,1995,15:5789-5799
[8]Copray JCVM,Ter Horst GJ,Liem RSB,et al.Neurotransmitters and neuropeptides within the mesencephalic trigeminal nucleus of the rat:an immunohistochemical analysis.Neurosci,1990,37:399-411
[9]Pang YW,Ge SN,Li JL,et al.Axon Terminals Expressing Vesicular Glutamate Transporter VGLUT1or VGLUT2Within the Trigeminal Motor Nucleus of the Rat:Origins and Distribution Patterns.J Comp Neurol,2009,512:595-612
[10]Pang YW,Li JL,Nakamura K,et al.Expression of vesicular glutamate transporter 1immunoreactivity in peripheral and central endings of trigeminal mesencephalic nucleus neurons in the rat.J Comp Neurol,2006,498:129-141
[11]Turman Jr JE,Ajdari J,Chandler SH.NMDA receptor NR1and NR2A/B subunit expression in trigeminal neurons during early postnatal development.J Comp Neurol,1999,409:237-249
[12]Turman Jr JE,MacDonald AS,Pawl KEW,et al.AMPA receptor subunit expression in trigeminal neurons during postnatal development.J Comp Neurol,2000,427:109-123
[13]Turman Jr JE,Hiyama L,Castillo M,et al.Expression of group I and II metabotropic glutamate receptors in trigeminal neurons during postnatal development.Dev Neurosci,2001,23:41-54
[14]Chandler SH.Evidence for excitatory amino acid transmission between mesencephalic nucleus of V and jawcloser motoneurons in the guinea pig.Brain Res,1989,477:252-264
[15]Katakura N,Chandler SH.An iontophoretic analysis of the pharmacologic mechanisms responsible for trigeminal motoneuronal discharge during masticatory-like activity in the guinea pig.J Neurophysiol,1990,63:354-369
[16]Forrest DM,Yuzaki HD,Soares L,et al.Targeted disruption of NMDA receptor 1gene abolishes NMDA response and results in neonatal death.Neuron,1994,13:325-338
图 版 说 明
图 三叉神经运动核光学显微镜像示大鼠生后不同发育阶段Ⅰ型囊泡膜谷氨酸转运体免疫组织染色。新生大鼠(图a),生后3d(图b),生后7d(图c),生后10d(图d),生后14d(图e)和成年大鼠(图f)。Vmo,三叉神经运动核。标尺=150μm (图a-f)。
EXPLANATION OF FIGURES
Fig.Microphotographs through the trigeminal motor nucleus showing vesicular glutamate transporter 1 immunoreactivty in newborn(Fig.a),3-day-old(Fig.b),7-day-old(Fig.c),10-day-old(Fig.d),14-day-old(Fig.e)and adult rat(Fig.f).Vmo,trigeminal motor nucleus.Bar=150μm (Fig.a-f).
猜你喜欢
海南医学院学报(2022年18期)2022-10-13
广东药科大学学报(2022年4期)2022-07-22
青岛大学学报(医学版)(2021年2期)2021-07-08
郑州大学学报(医学版)(2021年1期)2021-03-03
中国药剂学杂志(网络版)(2021年1期)2021-02-24
中国实用神经疾病杂志(2020年21期)2020-12-04
中国人兽共患病学报(2018年10期)2018-12-08
中国当代医药(2018年21期)2018-11-10
中成药(2018年8期)2018-08-29
星星·散文诗(2016年23期)2016-12-29
