
鞘氨醇激酶通过调控血管发生促进肝癌转移
2012-02-03 07:30赛岩代学强白纪民刘勇常菁范礼斌崔春萍
中国医药生物技术 2012年3期
赛岩,代学强,白纪民,刘勇,常菁,范礼斌,崔春萍
鞘氨醇激酶通过调控血管发生促进肝癌转移
赛岩,代学强,白纪民,刘勇,常菁,范礼斌,崔春萍
目的确定鞘氨醇激酶(SPK)在肝癌转移中的作用。
方法用 Ad-SPK1 腺病毒感染肝癌细胞 LM-3 使其 SPK过表达,Western blot 检测 SPK1 表达及激活情况;Transwell、划痕实验检测肝癌细胞迁移情况。体外管状结构形成实验检测 SPK 对脐静脉内皮细胞(HUVEC)管状化形成的影响。建立 SPK 过表达裸鼠模型,体内检测肝癌迁移情况。
结果在体外 SPK 对肝癌细胞的迁移无显著影响;Ad-SPK 能够促进 HUVEC 管状结构形成;裸鼠皮下移植瘤结果显示 Ad-SPK 促进肿瘤生长和转移。
结论鞘氨醇激酶通过调控血管发生促进肝癌转移。
疾病模型,动物; 肝肿瘤; 肿瘤移植; 移植,异种; 鞘氨醇激酶
1 材料和方法
1.1 材料
1.1.1 实验动物和细胞 BALB/c 裸鼠,雌性,4 周龄,体重 16 ~ 18 g,购自军事医学科学院实验动物中心,饲养于解放军总医院 SPF 级动物房。人肝癌细胞 LM-3 购自中国科学院上海细胞库;人脐静脉内皮细胞(HUVEC)购自中国协和细胞库;人胚肾细胞(HEK293)购自中国科学院上海细胞库。
1.1.2 主要试剂与仪器 BCA-200 蛋白检测试剂盒购自美国 Pierce 公司;蛋白质分子量标准购于北京全式金生物技术公司;DMEM 培养基、小牛血清购自美国 Gibco 公司;ECM 培养基、内皮细胞生长添加物(ECGS)均购自美国 ScienCell 公司;胰蛋白酶购自美国 Sigma 公司;基质胶(Matrigel 胶)购自美国 BD 公司;SPK1 抗体购自美国 Abcam 公司;GAPDH 抗体购自中杉金桥公司;其他试剂均为国产分析纯试剂。细胞培养器材购自美国 Costar 公司;电泳仪、转膜仪为美国Bio-Rad 公司产品。
1.2 方法
1.2.1 Ad-SPK 的扩增、纯化及滴度的测定 将HEK293 细胞接种于 150 mm 平皿中,待细胞生长至 90% 汇合状态时,加入 Ad-SPK1 病毒,感染复数(multiplicity of infection,MOI)约为 10 个蚀斑形成单位(plaque forming unit,pfu)/细胞[7],36 ~ 48 h 后,当细胞出现完全病变(CPE)时,收集细胞,冻存于 -70 ℃,待累计到 50 ~ 60 个平皿时,统一纯化病毒。
将出现 CPE 的 HEK293 细胞于 -70 ℃ 和37 ℃ 之间反复冻融 3 次,2500 r/min 离心 10 min,除去细胞碎片;配制 CsCl 密度梯度液,将冻融后的待纯化病毒液加于各管的 CsCl 梯度液之上;10 ℃,136 000 ×g离心 1.5 h,于 1.35 g/ml CsCl溶液和 1.25 g/ml CsCl 溶液之间出现白色雾状病毒带;收集病毒带,并与 1.35 g/ml CsCl 溶液混合,10 ℃,136 000 ×g离心 5.5 h,进一步纯化病毒,以 PBS 液为透析液于 4 ℃ 透析病毒,每 2 小时更换一次透析液,共换 5 次;取出纯化的病毒液,加无菌甘油至终浓度为 10%,分装,-70 ℃ 保存。用 TCID50 法测定病毒滴度(IU/ml)[8]。
1.2.2 细胞的感染效率确定 为评价 Ad-SPK 的感染效率,以 Ad5/F11p-GFP(刘杨博士提供,感染滴度 1.0 × 1010pfu/ml)为对照,分别用 0、50、100、150、200 MOI 感染肝癌细胞 LM-3和脐静脉内皮细胞 HUVEC,2 h 后换液,继续培养 48 h 后荧光显微镜观察绿色荧光蛋白的表达情况,收集细胞进行流式细胞仪检测[9]。
1.2.3 SPK 在肝癌细胞中的表达及激酶活性鉴定 用腺病毒 Ad-SPK 感染 LM-3 细胞。48 h 和72 h 后收集细胞,弃上清,加入冰预冷的 1 ml PBS重悬细胞,500 ×g离心 10 min,吸净上清,加入预冷的细胞裂解液;4 ℃ 放置 30 min 以上使细胞彻底裂解,随后 16 000 ×g、4 ℃ 离心 30 min,收集上清。蛋白定量后,进行 SDS-PAGE 凝胶电泳,将蛋白转移至 PVDF 膜上,置于 5% 脱脂牛奶中封闭,用 SPK 多克隆抗体检测 LM-3 细胞中SPK 的表达及 Ad-SPK 对 LM-3 细胞中 SPK 的激活情况[10]。
1.2.4 Transwell 和划痕实验检测肝癌细胞迁移情况 Transwell:24 孔板下室加入 600 µl 含 5%FBS 的 DMEM 培养基,将小室嵌入。Ad-SPK 感染 LM-3 细胞 36 h 后,胰酶消化细胞,使浓度为1 × 106个/ml,悬于 2% FBS 的 DMEM 培养基中,每孔加入 200 μl;37 ℃ 常规培养 12 ~ 48 h;将 Transwells 转移至另一加有 4% 多聚甲醛的孔中,固定细胞 15 min;结晶紫染色 10 min;用棉签轻轻擦去膜上未迁移细胞,光镜下观察结果[11]。
划痕实验:用马克笔在 6 孔板背面画横线。Ad-SPK 感染 LM-3 细胞 36 h 后且细胞汇合率100%,划痕,PBS 洗 2 遍;无血清 DMEM 培养液继续培养 24 h 后显微镜下观察划痕距离[12]。
1.2.5 HUVEC 体外管状结构形成实验 用添加ECGS 的 ECM 培养基常规培养 HUVEC,待细胞长至密度 70% ~ 80%,PBS 轻洗 4 ~ 5 次后加入无血清无生长因子的 ECM 培养基 10 ml。收集培养 24 h 后的条件培养基,500 ×g,4 ℃ 离心10 min,除去死细胞。将上清转移 EP 管中 -80 ℃备用。
将 Matrigel 胶放于冰水中,置于 4 ℃ 冰箱过夜,使胶完全融化[13]。并预冷所需的吸管、枪头和培养板;将培养板放在冰上,将胶混匀,用遇冷的无血清无生长因子的 ECM 培养基将 Matrigel 胶稀释为 30% 的浓度。按 50 μl/cm2比例将胶液加入培养板中;将培养板置于 37 ℃ 细胞培养箱,放置 1 h,使胶凝固。将无血清无生长因子的 ECM 培养基与收集的条件培养基等比例混合,加入胎牛血清,使其浓度达到 10% ~ 20%,将该混合物加到凝固的 Matrigel 胶上。按每孔 1.5 × 105个/100 μl 加入细胞,6 h 后显微镜下观察结果[14]。
1.2.6 SPK 过表达裸鼠模型的建立及组织学检测 皮下种植:选取 4 周龄雌性 BALB/c 裸鼠12 只,分为 Ad-SPK-LM-3 组与 Ad-null-LM-3组,每组 6 只。取对数生长期的肝癌细胞 LM-3,分别感染 Ad-SPK和 Ad-null 36 h 后消化计数,PBS 洗涤 2 次,重悬于 PBS 中制备成浓度为 1 ×107个/100 μl 的细胞悬液。以 100 μl/只的细胞悬液接种于裸鼠左前肢腋下[15],3 d 后观察皮下成瘤情况。接种 7 d 后,瘤体成型,此时瘤内注射腺病毒[16-17]。每周注射 2 次至实验终点。7 周后颈椎脱臼法处死小鼠,将肿瘤组织剥离,取出肝脏和肺脏固定于 10% 缓冲福尔马林液中 48 h,将肺脏每隔 2 mm 水平剖开查找转移灶,进行脱水、包埋、切片、HE 染色。
尾静脉注射:选取 4 周龄雌性 BALB/c 裸鼠12 只,分为 Ad-SPK-LM-3组与 Ad-null-LM-3组,每组 6 只。取对数生长期的肝癌细胞 LM-3,分别感染 Ad-SPK 和 Ad-null 36 h 后消化计数,PBS 洗涤 2 次,重悬于 PBS 中制备成浓度为 1 ×106个/100 μl 的细胞悬液。以 100 μl/只的细胞悬液注射于裸鼠尾静脉[18-19]。7 d 后,尾静脉注射腺病毒。每周注射 2 次至实验终点。4 周后颈椎脱臼法处死小鼠,取出肝脏和肺脏固定于 10% 缓冲福尔马林液中 48 h,将肺脏每隔 2 mm 水平剖开查找转移灶,进行脱水、包埋、切片、HE 染色。
1.3 统计学处理
应用 SAS 8.2 统计学软件进行数据处理。实验组与对照组检测结果的比较采用 Student’st检验,均以P< 0.05 为差异有统计学意义。
2 结果
2.1 重组腺病毒的感染滴度
重组腺病毒在 HEK293 细胞中大量扩增(图1),收集细胞,释放出病毒。经 CsCl 密度梯度和连续密度超速离心,获得高度浓缩的腺病毒带。感染滴度为 1.0 × 1011pfu/ml。
2.2 Ad5/F11P-GFP 对 LM-3 细胞和 HUVEC 细胞的感染效率
结果如图 2所示:随着 MOI 的增加 Ad5/F11P-GFP 对 LM-3 细胞和 HUVEC 细胞的感染效率增加,在 50 MOI 时均高达 90% 以上,说明Ad5/F11P 腺病毒载体对细胞的感染效率满足后续实验要求。

图 1 HEK293 细胞出现病变效应(A:正常细胞;B:病变区域)Figure 1 HEK293 cells with CPE (A: HEK293 cells; B:CPU)
2.3 SPK 在肝癌细胞中的表达
肝癌细胞 LM-3 中的 SPK 活性高于正常细胞,利用这株细胞为模型,检测 Ad-SPK 对 SPK的激活情况。结果如图 3 所示:Ad-SPK 能有效激活 SPK酶活性。
2.4 Ad-SPK 对 LM-3 细胞的迁移的影响
结果如图 4、图 5 所示:Ad-SPK 感染 LM-3细胞 36 h 后对细胞迁移并没有显著影响。Transwell实验结果显示,穿过小室的细胞并无明显增加;划痕实验结果显示未促进肝癌细胞迁移。

图 2 Ad5/F11P-GFP 对细胞的感染效率Figure 2 Cells were transfected with Ad5/F11P-GFP

图 3 Ad-SPK 对 LM-3 细胞 SPK 酶活性的激活Figure 3 Activation of SPK by Ad-SPK

图 4 SPK 诱导 LM-3 细胞的迁移[A:光学显微镜下观察的细胞迁移结果(100 ×);B:镜下细胞计数结果]Figure 4 The effect of SPK to migration ability of LM-3 cells (A: The result of LM-3 cell migration; B: The result of LM-3 cell count)
2.5 Ad-SPK 促进 HUVEC 细胞管状化结构形成
用腺病毒 Ad-SPK 感染 HUVEC 24 h 后,收集细胞进行管状化实验分析,结果如图 6 所示,50 MOI Ad-SPK 感染能够显著增加内皮细胞的成管。

图 5 SPK 诱导 LM-3 细胞的迁移光学显微镜下观察的细胞迁移结果(100 ×)2.0 mmFigure 5 The effect of SPK to migration ability of LM-3 cell
2.6 Ad-SPK 在体内促进肿瘤生长
实验终点时脱臼法处死小鼠,剥取肿瘤,去除表面脂类和结缔组织,如图 7A。可见瘤体表面光滑,表面被一层薄膜包裹,形状不规则,质软。统计肿瘤体积和重量如图 7B。结果显示 Ad-SPK 在体内明显促进肿瘤生长,随瘤内注射病毒刺激7 周后移植瘤体积明显高于正常组。其称重结果显示,正常组较病毒组轻。
2.7 Ad-SPK 在体内促进肿瘤生长
实验终点时脱臼法处死小鼠,剥取肿瘤,去除表面脂类和结缔组织,如图 7A。可见瘤体表面光滑,表面被一层薄膜包裹,形状不规则,质软。统计肿瘤体积和重量如图 7B。结果显示 Ad-SPK 可在体内促进肿瘤生长,瘤内注射病毒刺激 7 周后移植瘤体积明显高于对照给药组。称重结果显示,对照给药组较病毒组轻 40% 左右。HE 染色结果显示肝肺部均未见转移。
2.8 Ad-SPK 在体内促进肿瘤转移
实验终点时脱臼处死小鼠,将肺脏每隔 2 mm水平剖开查找转移灶,Ad-null-LM-3 组 6 只小鼠肺部切面正常;Ad-SPK-LM-3 组 6 只小鼠中 4 只可见灰白色粟粒状结节。取全部小鼠肝脏和肺脏做HE 切片染色如图 8,肝脏中均无明显转移灶;尾静脉注射病毒组在肺部发现多处转移灶,对照给药组未见转移灶。
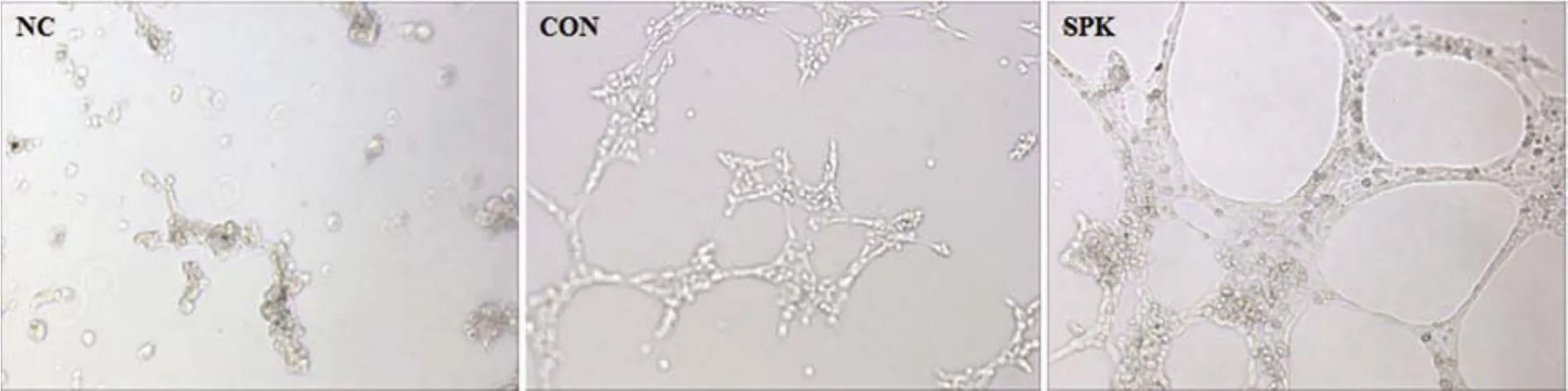
图 6 SPK 对 HUVEC 细胞管状化结构形成的影响(100 ×)(NC:DMEM 培养 HUVEC 组;CON:正常 HUVEC 组;SPK:Ad-SPK 感染 HUVEC 组)Figure 6 Effect of SPK on the tubular morphogenesis of HUVEC cells (100 ×).The cells were stimulated with Ad-SPHK (MOI 50)and plated on the Matrigel-coated 24-well plate; NC: DMEM culture; CON: Control group; SPK: SPK treat group
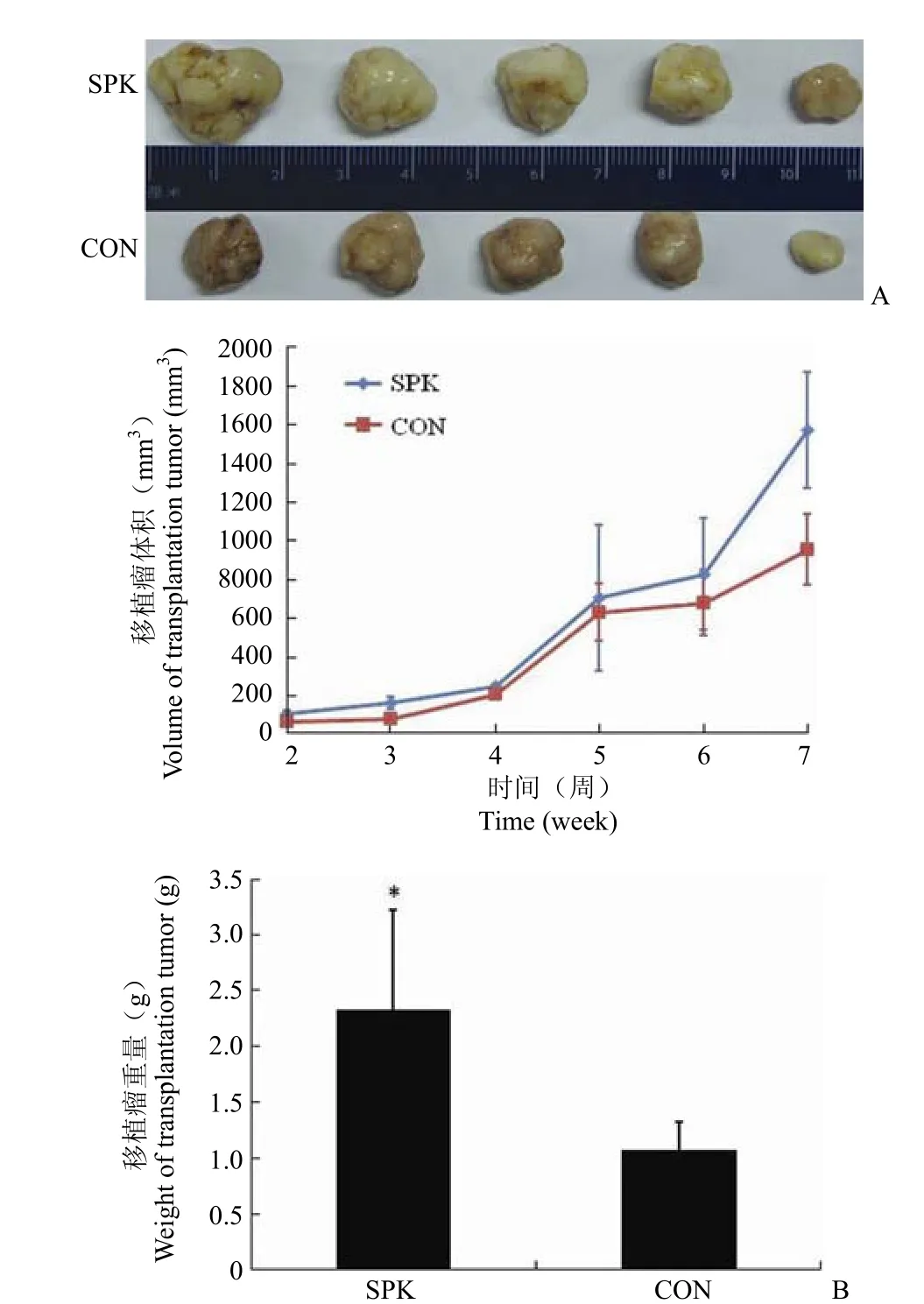
图 7 移植瘤的体积和重量统计[A:皮下移植瘤的大体观察;B:移植瘤的体积和重量(SPK:Ad-SPK 给药组;CON:Ad-null 对照给药组)]Figure 7 Volume and weight of the transplantation tumors of SPK and control groups [A: Observation of tumor; B: Volume and weight of the transplantation tumors (SPK: Ad-SPK treat group; CON: Ad-null treat group)]

图 8 裸鼠肺脏 HE 染色(红色箭头:转移灶)Figure 8 Histology of lung sections (HE stain, original magnification ×400; Red-arrow: observed in lung sections of tumor)
3 讨论
磷脂代谢产生多种磷脂代谢产物,如神经酰胺、SPK、S1P 等。SPK 是细胞内合成 S1P 的关键酶,它可以使 SPK 磷酸化生成 S1P,是细胞增殖及存活的重要信号分子。S1P 在人胃腺癌细胞的增殖中起着极其关键的调控作用[20]。在氧化偶氮甲烷(azoxymethane,AOM)诱发的小鼠肠腺癌中,SPK 在 mRNA 水平的表达增高,说明 SPK/S1P信号途径与结肠癌的发生是密切相关的[21]。
SPK 与肝癌相关报道仍不多见,肝脏是磷脂类代谢的重要器官,SPK/S1P 作为细胞微环境的重要信号途径之一,其调节肝癌发生发展的作用和机制并不清楚。目前许多研究只停留在体外试验,还有一些问题需要进一步研究,例如 SPK 与肿瘤的关系,本文就 SPK/S1P 信号通路和肝癌转移的关系作为研究重点,在体内外研究 SPK 与肝癌转移是否存在联系。
在体外实验中,我们用 Ad-SPK 感染肝癌细胞系 LM-3,使其高表达 SPK,用 Transwell、划痕实验检测肝癌细胞迁移情况。结果显示,SPK 在肝癌细胞中高表达并不能明显促进其迁移。但在体内实验中我们却发现 Ad-SPK 可以明显促进肝癌细胞转移。我们推断 SPK 可能是通过调控血管发生促进肝癌转移。动物实验中首先我们用 Ad-SPK 感染 LM-3 细胞 36 h,收集细胞接种于裸鼠皮下。因为腺病毒 Ad-SPK 不能长期稳定使 SPK 高表达,所以待成瘤后我们于瘤体内多点注射病毒以使其持续 SPK 高表达。实验终点处死小鼠发现肿瘤周围血管丰富,肿瘤体积及重量都有明显区别,但体内转移并不明显。之后我们同样处理的细胞给裸鼠尾静脉注射。据文献[22-23]报道,肝癌细胞尾静脉注射约 5 ~ 6 周发生转移,鉴于肝癌本身具备高转移性,本研究中尾静脉注射小鼠 4 周处死,拟选择这个时间点是为了使 Ad-null-LM-3 组与Ad-SPK- LM-3 组产生明显区别。处死小鼠后,将肺部每隔 2 mm 水平剖开查找转移灶,Ad-null-LM-3 组 6 只小鼠肺部切面正常;Ad-SPK-LM-3组 6 只小鼠中 4 只可见灰白色粟粒状结节。HE染色后 Ad-null-LM-3 组切片中未明显转移灶,但尾静脉补注 Ad-SPK 组肺部见多处转移灶。
大量研究表明,VEGF 在肝癌组织中表达较肝硬化和正常肝组织显著增高[24]。VEGF 高表达有利于原发性肝癌的复发、侵袭和转移,并且与肝癌的临床病理特征密切相关。文献[25]报道,在内皮细胞及膀胱癌细胞中,VEGF 可以通过蛋白激酶 C(protein kinase C,PKC)激活 SPK 信号途径,表明 SPK/S1P 参与 VEGF 的信号传导,但 SPK/S1P是否参与调节 VEGF 的表达和其在肝癌细胞信息传导通路间的相互作用仍不清楚。人肝癌细胞HepG2 能够合成和分泌 VEGF,而且经不同浓度的 DMS 作用 12 h 后,其 VEGF 的表达较对照组明显减少。因此,在肝癌细胞中,SPK/S1P 信号途径是调节 VEGF 信号传导的重要途径之一[26-27]。所以我们用 Ad-SPK 感染 HUVEC,发现高表达SPK 可以明显促进脐静脉内皮细胞管状化形成,较未处理的 HUVEC 管状化密度更大。该结果与体内实验结果吻合。
在体内研究中,我们通过注射 Ad-SPK 促进其VEGF 表达从而促进肿瘤的生长。SPK/S1P 是维持细胞存活的重要分子,并且参与了肝癌细胞的增殖及肿瘤血管生成的调节过程。
[1]Tanaka M, Katayama F, Kato H, et al.Hepatitis B and C virus infection and hepatocellular carcinoma in China: a review of epidemiology and control measures.J Epidemiol, 2011, 21(6):401-416.
[2]El-Serag HB.Hepatocellular carcinoma: an epidemiologic view.J Clin Gastroenterol, 2002, 35(5 Suppl 2):S72-S78.
[3]Breuhahn K, Longerich T, Schirmacher P.Dysregulation of growth factor signaling in human hepatocellular carcinoma.Oncogene, 2006,25(27):3787-3800.
[4]LaBrecque DR, Pesch LA.Preparation and partial characterization of hepatic regeneration stimulator substance from rat liver.J Physiol,1975, 248(2):273-284.
[5]LaBrecque DR, Steek G, Fogertys, et al.Purification and physical-chemical characterization of hepatic stimulator substance.Hepatology, 1987, 7(1):100-106.
[6]Cuvillier O, Levade T.Sphingosine 1-phosphate antagonizes apoptosis of human leukemia cells by inhibiting release of cytochrome c and Smac/DIABLO from mitochondria.Blood, 2001, 98(9):2828-2836.
[7]Moehler TM, Ho AD, Goldschmidt H, et al.Angiogenesis in hematologic malignancies.Crit Rev Oncol Hematol, 2003, 45(3):227-244.
[8]Bonhoure E, Pchejetski D, Aouali N, et al.Overcoming MDR-associated chemoresistance in HL-60 acute myeloid leukemia cells by targeting sphingosine kinase-1.Leukemia, 2006, 20(1):95-102.
[9]Yabu T, Tomimoto H, Taguchi Y, et al.Thalidomide-induced antiangiogenic action is mediated by ceramide through depletion of VEGF receptors, and is antagonized by sphingosine-1-phosphate.Blood, 2005, 106(1):125-134.
[10]Krump-Konvalinkova V, Yasuda S, Rubic T, et al.Stable knock-down of the sphingosine 1-phosphate receptor S1P1 influences multiple functions of human endothelial cells.Arterioscler Thromb Vasc Biol,2005, 25(3):546-552.
[11]Tarui T, Majumdar M, Miles LA, et al.Plasmin-induced migration of endothelial cells.A potential target for the anti-angiogenic action of angiostatin.J Biol Chem, 2002, 277(37):33564-33570.
[12]Hla T.Signaling and biological actions of sphingosine 1-phosphate.Pharmacol Res, 2003, 47(5):401-407.
[13]Szpaderska AM, DiPietro LA.In vitro matrigel angiogenesis model.Methods Mol Med, 2003, 78:311-315.
[14]Duan HF, Wu CT, Lu Y, et a1.Sphingosine kinase active-tion regulates hepatocyte growth factor induced migration of endothelial cells.Exp Cel Res, 2004, 298(2):593-601.
[15]Shu X, Wu W, Mosteller RD, et al.Sphingosine kinase mediates vascular endothelial growth factor-induced activation of ras and mitogen-activated protein kinases.Mol Cell Biol, 2002, 22(22):7758-7768.
[16]Xin X, Yang S, Ingle G, et al.Hepatocyte growth factor enhances vascular endothelial growth factor-induced angiogenesis in vitro and in vivo.Am J Pathol, 2001, 158(3):1111-1120.
[17]Endo A, Nagashima K, Kurose H, et al.Sphingosine 1-phosphate induces membrane ruffling and increases motility of human umbilical vein endothelial cells via vascular endothelial growth factor receptor and CrkII.J Biol Chem, 2002, 277(26):23747-23754.
[18]Kluk MJ, Colmont C, Wu MT, et al.Platelet-derived growth factor(PDGF)-induced chemotaxis does not require the G protein-coupled receptor S1P1 in murine embryonic fibroblasts and vascular smooth muscle cells.FEBS Lett, 2003, 533(1-3):25-28.
[19]Li QF, Huang WR, Duan HF, et al.Sphingosine kinase-1 mediates BCR/ABL-induced upregulation of Mcl-1 in chronic myeloid leukemia cells.Oncogene, 2007, 26(57):7904-7908.
[20]Li QF, Wu CT, Duan HF, et al.Activation of sphingosine kinase mediates suppressive effect of interleukin-6 on human multiple myeloma cell apoptosis.Br J Haematol, 2007, 138(5):632-639.
[21]Li QF, Wu CT, Guo Q, et al.Sphingosine 1-phosphate induces Mcl-1 upregulation and protects multiple myeloma cells against apoptosis.Biochem Biophys Res Commun, 2008, 371(1):159-162.
[22]Ogawa K, Nakanishi H, Takeshita F, et al.Establishment of rat hepatocellular carcinoma cell lines with differing metastatic potential in nude mice.Int J Cancer, 2001, 91(6):797-802.
[23]Ogawa K, Pitchakarn P, Suzuki S, et al.Silencing of connexin 43 suppresses invasion, migration and lung metastasis of rat hepatocellular carcinoma cells.Cancer Sci, 2012, 103(5):860-867.
[24]Guo Q, Li QF, Liu HJ, et al.Sphingosine kinase 1 gene transfer reduces postoperative peritoneal adhesion in an experimental model.Br J Surg, 2008, 95(2):252-258.
[25]Purdie KJ, Whitley GS, Johnstone AP, et al.Hepatocyte growth factor-induced endothelial cell motility is mediated by the upregulation of inducible nitric oxide synthase expression.Cardiovasc Res, 2002, 54(3):659-668.
[26]Younes MN, Park YW, Yazici YD, et a1.Concomitant inhibition of epidermal growth factor and vascular endothelial growth factor reptor tyrosine kinases reduces growth and metastasis of human salivary adenoid cystic carcinoma in an orthotopic nude mouse model.Mol Cancer Ther, 2006, 5(11):2696-2705.
[27]Hirohashi T, Yamamoto T, Uenishi T, et a1.CD44 and VEGF expression in extrahepatic metastasis of human hepatic carcinoma.Hepatogastroentemlogy, 2004, 51(58):1121-1123.
www.cmbp.net.cn Chin Med Biotechnol, 2012, 7(3):171-177
Sphingosine kinase promote HCC metastasis via regulation of angiogenesis
SAI Yan, DAI Xue-qiang, BAI Ji-min, LIU Yong, CHANG Jing, FAN Li-bin, CUI Chun-ping
ObjectiveTo determine the role of sphingosine kinase (SPK) in hepatocyte carcinoma (HCC) metastasis.
MethodsThe expression and activation of SPK in Ad-SPK infected LM-3 HCC cell line was detected by Western blot; and HCC cells migration was evaluated by Transwell and scar-healing experiments.The impact of SPK on HUVEC tube morphologenesis was examined byin vitrotube formation assay.The SPK-overexpressing nude mice model was established to access HCC metastasisin vivo.
ResultsSPK did not influence significantly HCC cells migrationin vitro; HUVEC tube formation was stimulate by Ad-SPK.Ad-SPK accelerated the tumor growth and metastasisin vivo.
Conclusion Sphingosine kinase promotes the HCC metastasis via regulating angiogenesis.
Disease models, animal; Liver neoplasms; Neoplasm transplantation; Transplantation, heterologous;Sphingosine kinase
s: FAN Li-bin, Email: lfan@ahmu.edu.cn; CUI Chun-ping, Email: cui_chunping2000@yahoo.com.cn
www.cmbp.net.cn 中国医药生物技术, 2012, 7(3):171-177
肝细胞癌(hepatocellular carcinoma,HCC)流行性广、病程短、病死率高,是严重危害人类健康的重大疾病。2010年统计数据显示我国每年约有34.4 万人死于肝细胞癌,占全球肝癌死亡人数的55%[1]。因此,深入研究肝癌发病机制并开发新的治疗策略是关系人类健康的重大科学问题。
肝癌浸润和转移特性是影响肿瘤患者生存和预后的关键因素,基质胶原纤维的降解、肿瘤细胞的迁移、肿瘤血管形成等过程参与了肿瘤的转移[2]。在许多肿瘤的发生发展过程中,鞘磷脂的代谢产物神经酰胺(ceramide,Cer)、神经鞘氨醇(sphingosine,Sph)和 1-磷酸鞘氨醇(sphingosine-1-phosphate,S1P)发挥着极为重要的作用,它们调节着细胞的增殖、存活和凋亡[3]。在细胞内,这些代谢产物可以相互转化,鞘氨醇激酶(sphingosine kinase,SPK)则是维系 Cer、Sph 和 S1P 三者间代谢平衡的关键酶,具有调节 Cer 和 S1P 的双重功能。SPK可诱导血管内皮细胞和血管平滑肌细胞等多种与血管生成有关的细胞的生物学反应,通过其表面受体EDGs 依赖的细胞骨架重组维持内皮细胞屏障的
10.3969/cmba.j.issn.1673-713X.2012.03.003
国家自然科学基金(30930041、30800558、81070330);“重大新药创制”科技重大专项(2012ZX09102301-012)
230032 合肥,安徽医科大学生命科学院生物学教研室(赛岩、范礼斌);100850 北京,军事医学科学院放射与辐射医学研究所(赛岩、代学强、白纪民、刘勇、常菁、崔春萍)
范礼斌,Email:lfan@ahmu.edu.cn;崔春萍,Email:cui_chunping2000@yahoo.com.cn
2012-02-20完整性[4],促进内皮细胞向血管样结构的形态转变等,从而完成血管内皮细胞的增殖和迁移[5]。SPK在多种实体肿瘤中均有表达,如结肠癌、乳腺癌、胃癌、肺癌,其基因具有癌基因的特征,并且通过SPK/S1P 信号途径对肿瘤发挥着促进细胞生长的作用[6]。本文主要分析鞘氨醇激酶在肝癌转移中的作用。
Author Affiliations: Biology Laboratory of Life Science Academy, Anhui Medical University, Hefei 230032, China (SAI Yan, FAN Li-bin); Institute of Radiation Medicine, Academy of Military Medical Sciences, Beijing 100850, China (SAI Yan, DAI Xue-qiang,BAI Ji-min, LIU Yong, CHANG Jing, CUI Chun-ping)
·信息站点·
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
世界科学技术-中医药现代化(2022年3期)2022-08-22
传染病信息(2022年3期)2022-07-15
中国动物保健(2022年2期)2022-05-05
现代临床医学(2021年5期)2021-11-02
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
湖南师范大学学报(医学版)(2020年6期)2020-02-23
中国眼镜科技杂志(2019年9期)2019-11-11
灾害医学与救援(电子版)(2018年1期)2018-06-05
