
烟草与TMV非亲和性互作中根系活力及CAT活性变化
2012-01-17 05:36:26马学萍刘开全刘永琼
中国烟草科学 2012年1期
马学萍,刘开全,和 云,刘永琼
(曲靖师范学院生物资源与环境科学学院,云南 曲靖 655011)
作为一种生物胁迫因子,病原物的侵染会使寄主植物在与其互作中产生明显的生理变化[1-3]。植物病害的发生与保护酶系活性变化有着密切关系[4]。陈学平等[5]研究烟草与 TMV不同互作体系中烟草叶片抗氧化酶系活性变化发现在互作早期 CAT活性较高,互作后期 CAT活性快速下降使得抗病系统保持较高水平的活性氧,诱导植物防卫基因的表达而产生抗病反应。李晔等[6]研究认为烟草幼苗叶片中CAT等酶的活性变化规律与烟株缓解TMV对烟草造成的伤害有一定的相关性。在育苗过程中TMV可通过根部侵染烟苗[7-8]。病原物侵染可能引起植物根系的生理变化,进而影响植株的抗病性。研究烟草接种病毒后根系的活力及CAT活性变化,其结果对探讨抗病品种的抗病机制具有一定的参考价值。
1 材料与方法
1.1 材料
以枯斑三生烟(Nicotiana tabacumcv ‘Samsun NN’)为供试烟草,烟草花叶病毒(TMV)普通株系为供试病毒。
1.2 方法
漂盘培育烟苗,在烟苗长到6片真叶时,在育苗池中加入病毒溶液使其接种 TMV的烟苗为处理,以不接种TMV的烟苗为对照。接种病毒后3、7、11、15、19、23、27、31 d分别取处理和对照烟株根系用高锰酸钾滴定法测定其CAT活性[9]用甲烯蓝法[10]测定其活力,重复 3次。采用 SPSS10.0软件进行数据处理,并用Excel软件绘图。
2 结 果
2.1 接种病毒后枯斑三生烟根系CAT活性变化
枯斑三生烟接种病毒后不同时间取样测得其根系CAT活力值(表1)。
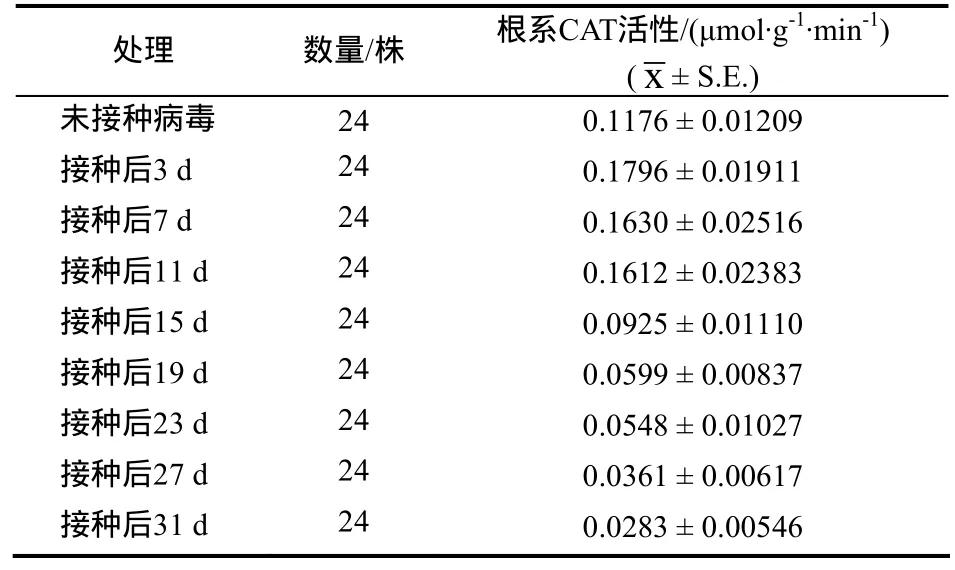
表1 不同取样时间测得的烟草根系CAT活力Table 1 The CAT activity of tobacco root in different sampling time
枯斑三生烟接种病毒后3、7、11 d的根系CAT活性高于对照组,接种病毒后15、19、23、27 d和31 d的根系CAT活性低于对照组。相对于对照,枯斑三生烟接种病毒后其根系 CAT活性呈先升高后下降趋势(图1)。即接种病毒后,枯斑三生烟根系CAT活性在迅速升高后下降并显著低于对照组。
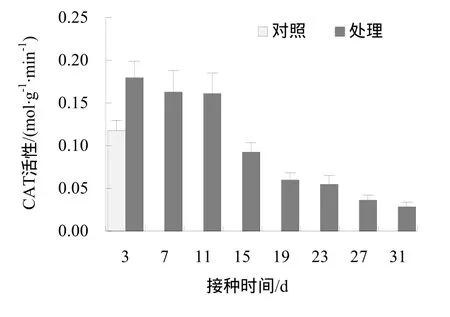
图1 不同处理下烟草根系CAT活性变化Fig.1 The CAT activity of tobacco root in different sampling time
单因素方差分析表明,接种后各时间点取样的根系CAT活性与对照间有极显著差异(p< 0.01),多重比较(Duncan法)显示各处理间的差异显著(表2)。将接种病毒后取样时间与对应的CAT活性值进行相关性分析结果表明,CAT活性值与取样时间有较强的负相关性(r= -0.9,p< 0.01)。
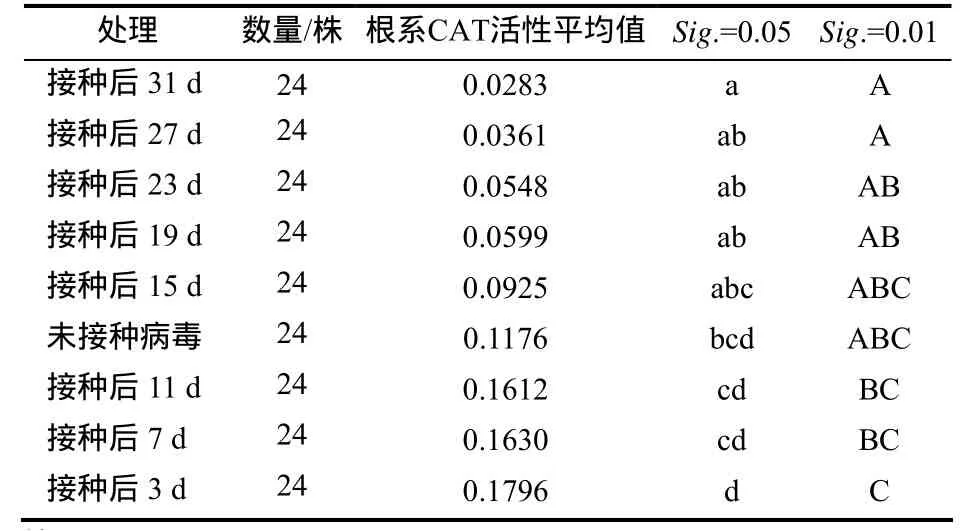
表2 各处理CAT活性的多重比较Table 2 Multiple comparisons of the CAT activity value in different sampling time
2.2 接种病毒后枯斑三生烟根系活力变化
枯斑三生烟接种病毒后3、7、11、15、19、23、27、31 d取样测得其根系活力(表3)。
相比对照,枯斑三生烟接种病毒后,其根系活力迅速升高之后降低,呈平稳趋势(图 2)。方差分析表明,处理间有显著差异(p=0.02<0.05);多重比较(LSD法)表明,接种病毒后 3、7、11 d后测得的烟草根系活力与未接种病毒的根系活力存在差异显著。
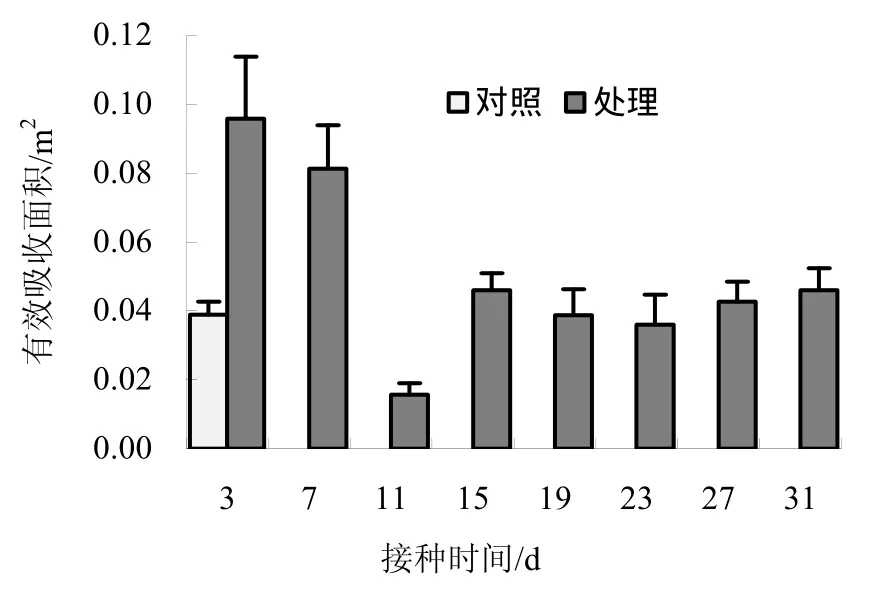
图2 不同取样时间测得的根系活力Fig.2 The root vigor of tobacco in different sampling time
3 讨 论
(1)作为植物细胞内保护酶系统的一个重要酶,CAT是植物细胞内重要的活性氧清除剂。病原物胁迫诱发植株体内活性氧的积累,在植物与病原物互作中,CAT活性发生变化诱导植物的抗病反应。一般认为,植物感染病原菌后CAT活性降低,或与抗病品种的非亲和性互作中其活性降低[11]。本研究结果表明,枯斑三生烟接种病毒后3、7、11 d的根系CAT活性高于对照组,接种病毒后15、19、23、27、31 d的根系CAT活性低于对照组。枯斑三生烟接种病毒后初期植株体内 CAT活性上升可能是烟草响应 TMV侵染伤害而导致的;之后其CAT活性反而快速降低,可能与病毒侵染诱导植株获得系统抗性,缓解了病毒对植株的生理胁迫。
(2)孙浩等研究[12]的 Cu2+胁迫对烟草幼苗根系活力的影响发现,Cu2+胁迫明显抑制植物的根系活力,且随着Cu2+的胁迫加强,烟草幼苗的根系活力呈明显下降趋势。本试验研究结果表明,枯斑三生烟接种病毒后,其根系活力迅速升高,即烟草在接种TMV后的3、7 d的根系活力明显高于未处理组,这可能是TMV侵染初期烟草通过提高根系活力来应对病原物对其胁迫。之后根系活力降低而后呈平稳趋势,并与对照无显著差异。这也可能与植株获得系统抗性有关。
[1]李兰真,赵会杰,杨会武,等.小麦锈病与叶片中活性氧代谢的关系[J].植物生理学通讯,1999,35(2):115-117.
[2]宋凤鸣,郑重,葛秀春,等.两种植物中膜脂质体氧化物与活性氧的作用[J].植物生理学通讯,1996,32(5):377-385.
[3]Kepler L D, Baker C J.Initiated lipid peroxidation in a bacteria induced hypersensitive reaction in tobacco cell suspensions[J].Phytopathology, 1989, 79: 555-559.
[4]Barbara Baker, Patricia Zambryski, Brian Staskawicz.Signaling in plant microbe interactions[J].Science, 1997,276(23): 726-733.
[5]陈学平,姚忠达,郭家明,等.不同烟草品种感染TMV病程过程中CAT、PAL活力变化研究[J].安徽农业大学学报,2002,29(2):l03-107.
[6]李晔,昊元华,赵秀,等.不同供 Fe水平下受 TMV感染的烟草中几种防御酶活性变化[J].安徽农业科学,2006,34(17):4237-4238.
[7]李凡,王钰丽,吴德喜,等.烤烟漂浮育苗中普通花叶病的主要传播途径[J].烟草科技,2006(10):53-55.
[8]曾嵘,滕永忠,张庆刚,等.烤烟漂浮育苗中TMV的发生及预防研究[J].云南农业大学学报,2005,20(1):136-139.
[9]萧浪涛,王三根.植物生理学实验技术[M].北京:中国农业出版社,2005:105-107.
[10]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:58-59.
[11]吴元华,文才艺,李浩戈,等.烟草感染马铃薯 Y病毒坏死株系后六种酶活性变化的研究[J].中国烟草学报,1999,5(2):30-34.
[12]孙浩,张艳英,宋金敏,等.Cu2+胁迫对烟草幼苗生长发育的影响[J].贵州农业科学,2008,36(5):34-36.
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06 01:48:20
当代水产(2022年1期)2022-04-26 14:35:30
浙江中西医结合杂志(2017年2期)2017-01-12 18:23:59
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
当代化工研究(2016年9期)2016-03-20 16:22:08
山东医药(2015年38期)2015-12-07 09:12:28
食品工业科技(2014年13期)2014-03-11 18:17:06
声屏世界(2014年6期)2014-02-28 15:18:09
食品科学(2013年24期)2013-03-11 18:30:53
