
拟南芥MPK17基因在玉米中的电子克隆分析
2012-01-12 03:59王俊红
长治学院学报 2012年2期
王 琦 ,王俊红 ,王 荣
(1.长治学院 生物科学与技术系,山西 长治 046011;2.陕西师范大学 生命科学学院,陕西 西安 710062;3.长治卫生学校,山西 长治 046000)
拟南芥MPK17基因在玉米中的电子克隆分析
王 琦1,王俊红2,王 荣3
(1.长治学院 生物科学与技术系,山西 长治 046011;2.陕西师范大学 生命科学学院,陕西 西安 710062;3.长治卫生学校,山西 长治 046000)
文章主要运用生物信息学的方法,利用拟南芥MPK17基因在玉米中进行了电子克隆,并得到了目的基因,进一步对其DNA碱基序列与蛋白质氨基酸序列进行了分析,旨在为更好地研究MPK17基因提供一定的理论资料。
拟南芥;玉米;MPK17;电子克隆
拟南芥(Arabidopsis thaliana)形态结构简单,茎直立,花小,株高15-30cm,生殖周期短,繁殖系数高[1-3]。其基因组全序列测序工作已于2000年12月完成[4],这样使得有关拟南芥基因克隆、结构与功能的研究发展迅速。且在已知基因组的高等植物中,拟南芥核基因组最小[5]。基因组中所含高度重复,中度重复及低度重复的比例也很低,绝大多数为单拷贝序列[6-8]。
MAP激酶(Mitogen activated protein kinases MPKs),中文全称为有丝分裂原活化蛋白激酶[9],它与其他一系列蛋白激酶构成级联放大信号通路[10],控制细胞生物学反应和参与植物对多种外界刺激的信号转导过程[11]。对MPK最早研究始于哺乳动物和酵母(Yeast),近年来,在拟南芥、苜蓿(Medicago sativa Linn)、烟草(Nicotiana tabacum)、水稻(O-ryza sativa)等高等植物中也取得了很大的进展[12-14]。文章主要通过生物信息学方法,分析了单子叶植物玉米(Zea mays)中存在的MPK17基因,旨在增强人类对于MPK17基因的认识,提高人类对玉米生长发育的控制能力,增加玉米抗旱、抗胁迫的能力,从而最终达到增加产量的目的。
1 材料与方法
1.1 材料
拟南芥MPK17基因编码的氨基酸序列、克隆得到的玉米新蛋白激酶的cDNA序列及其编码的氨基酸序列等。
1.2 方法
1.2.1 玉米中MPK17基因的电子克隆分析
从GenBank的核酸数据库中检索拟南芥MPK17基因,获得MPK17基因cDNA序列,以该序列为模板对玉米EST数据库进行BLAST检索,获得与之部分同源的EST群,从中选取一条EST作为种子序列BLAST检索玉米的EST数据库,将检出与种子序列同源性较高或有部分重叠的EST序列拼接组装为重叠群,再以此重叠群序列重复以上BLAST检索过程,反复进行EST重叠群序列的拼接和比对,直至检出所有的重叠EST或重叠群不能继续延伸,最终获得玉米MPK17基因的cDNA全序列。
1.2.2 玉米MPK17基因及其编码蛋白的生物信息学分析
(1)ORF分析
将之前电子克隆得到的玉米MPK17基因的cDNA在ORFFinder中进行开放读码框(ORF)分析。
(2)玉米MPK17基因编码蛋白的一级结构分析
首先利用在线分析工具ExPASy tool对玉米MPK17基因编码蛋白进行分子量分析,然后利用BIOEDIT软件对玉米MPK17基因编码蛋白的氨基酸序列分布进行分析。
(3)玉米MPK17基因编码蛋白的二级结构分析
利用蛋白质二级结构在线预测工具ExPASy Secondery Structure Prediction GOR对玉米MPK17基因编码蛋白进行二级结构分析。
(4)玉米MPK17基因编码蛋白的三级结构分析
利用网络同源建模服务器SWISS-MODEL对玉米MPK17基因编码蛋白进行三级结构预测[15-17]。
(5)蛋白质等电点分析
利用生物软件ANTHEPROT对玉米MPK17基因编码蛋白进行等电点分析。
(6)蛋白质信号肽分析
利用在线生物软件ExPASy Signal P对玉米MPK17基因编码蛋白进行信号肽分析。
(7)蛋白质疏水性与跨膜区分析
利用生物软件TopPred和TMHMM Server v.2.0分别对玉米MPK17基因编码蛋白的疏水性与跨膜区进行分析。
(8)蛋白质电荷分布分析
利用在线生物软件SAPS对玉米MPK17基因编码蛋白进行电荷分布分析。
(9)蛋白质保守域分析
利用在线蛋白质保守域分析工具NCBICDSearch对玉米MPK17基因编码蛋白进行结构域分析。
2 结果与分析
2.1 玉米MPK17基因的电子克隆
按照上述方法,使用拟南芥MPK17基因所编码的氨基酸序列(NCBI登录号:Q84M93.1)在玉米EST数据库中利用tBLASTn程序进行同源检索,最后将所有检索到的EST进行拼接获得一个长度为2344bp序列。然后把此序列在玉米EST数据库里进行BLASTn检索,结果表明,该序列受玉米EST数据库完全支持(如图1)。

图1 玉米MPK17基因编码蛋白序列检索玉米EST数据库Fig.1 EST database results of protein coded by corn MPK17 gene
2.2 玉米MPK17基因的ORF分析
开放阅读框(ORF)的识别是证明一个新的DNA序列为特定的蛋白质编码基因的部分或全部的先决条件。ORF是基因序列的一部分,包含一段可以编码蛋白的碱基序列,不能被终止子打断。由于蛋白质由三联体密码子所编码,所以一个双链DNA序列就有6个潜在的阅读框。其中一条链的3个读框称为正向读框,互补链上的三个读框称为反向读框。本文将电子克隆得到的玉米MPK17基因的cDNA在ORF Finder中进行ORF分析,结果如图2所示,不难发现在259-1998 bp之间有一个最长的完整ORF,长为1740 bp,编码579个氨基酸。

图2 玉米MPK17基因的ORFFinder结果Fig.2 ORFF inderr esult of corn MPK17 gene
2.3 玉米MPK17基因编码蛋白一级结构分析
利用在线生物软件ExPASy预测玉米MPK17基因编码蛋白的分子量约为66.04 kDa。利用BIOEDIT软件预测其氨基酸序列组成情况如图3所示。

图3 玉米MPK17基因编码蛋白的氨基酸组成Fig.3 The amino acid composition of protein coded by corn MPK17 gene
2.4 玉米MPK17基因编码蛋白二级结构分析
运用蛋白质二级结构在线分析工具GOR4对玉米MPK17基因编码蛋白进行分析,结果如图4所示,其中h代表α螺旋,e代表β折叠,c代表无规则卷曲。且α螺旋占35.06%;β折叠占15.20%;无规则卷曲占49.74%。由此可知,该玉米MPK17基因编码蛋白是由大量α螺旋、无规则卷曲以及少量的β折叠所构成的。

图4 GOR4分析玉米MPK17基因编码蛋白的二级结构Fig.4 Secondary structures predicted by GOR4 of coded protein by corn MPK17 gene
2.5 玉米MPK17基因编码蛋白三级结构分析
利用基于同源建模的分析工具Swiss-Model对玉米MPK17基因编码蛋白进行三级结构分析,软件自动选择PDB数据库中匹配度最高的的3py3A(磷酸化的p38-alpha MAPK激酶的晶体结构)作为模板,对目的蛋白进行三级结构的预测。由图5可知,目的蛋白的确是由大量α螺旋、无规卷曲以及少量的β折叠共同构成的复杂结构,与上述二级结构的分析结果一致。

图5 蛋白质的三级结构Fig.5 Tertiary structure of protein
2.6 蛋白质等电点分析
利用生物软件ANTHEPROT对玉米MPK17基因编码蛋白进行分析,结果如图6所示,玉米MPK17基因编码蛋白的理论等电点为6.43。
2.7 蛋白质信号肽分析
对蛋白质进行信号肽分析,有助于蛋白质功能域的区分及蛋白质细胞定位。用Signal P对玉米MPK17基因编码蛋白进行信号肽分析,结果如图7所示,通过直观分析氨基酸分值,可知该玉米MPK17基因编码蛋白的分值曲线不典型,因而得知玉米MPK17基因编码蛋白没有信号肽,属于非分泌性蛋白。
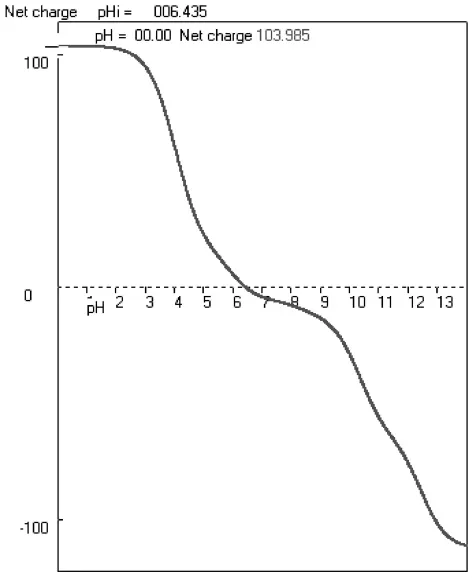
图6 等电点分析Fig.6 The anal ysis of isoel ectric point

图7 玉米MPK17基因编码蛋白信号肽分析Fig.7 Singal peptide prediction in protein coded by corn MPK17 gene
2.8 蛋白质疏水性和跨膜区分析
利用TopPred软件对玉米MPK17基因编码蛋白进行分析,结果如图8所示,横坐标表示蛋白质中氨基酸残基的序号,纵坐标表示残基的亲水/疏水特性,正值为疏水,负值为亲水。因此,可以判断得出,该蛋白质为亲水性蛋白。
利用TMHMM Server v.2.0对玉米MPK17基因编码蛋白跨膜区进行分析,结果如图9所示,其中横坐标代表蛋白质中氨基酸残基的序号,纵坐标代表可能性的大小,结果表明此蛋白质不存在跨膜区,说明不是跨膜蛋白。

图8 玉米MPK17基因编码蛋白亲/疏水性分析Fig.8 Hydrophilic/hydrophobic analysis of protein coded by corn MPK17 gene
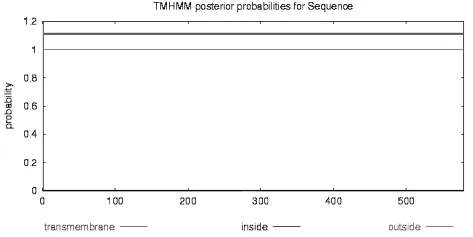
图9 玉米MPK17基因编码蛋白跨膜区Fig.9 Striding filmsection of protein coded by corn MPK17 gene
2.9 蛋白质电荷分布分析
利用在线分析工具SAPS对玉米MPK17基因编码蛋白氨基酸序列进行蛋白质电荷分布分析,结果如图10所示,该蛋白质含有88个带负电荷的氨基酸(Glu和Asp)和82个带正电荷的氨基酸(Arg和Lys),因此该玉米MPK17基因编码蛋白总计带6个负电荷。
2.10 蛋白质保守域分析
运用保守域(CD)分析玉米MPK17基因编码蛋白的结构域,结果表明,玉米MPK17基因编码蛋白中的保守结构域包括ATP结合位点,底物结合位点,活性位点以及KIM(即激酶相互作用模体)停泊位点,它们所在的具体位置则如图11所示。
3 讨论
电子克隆技术是伴随着基因定位、人类基因组测序及生物信息学技术的发展而出现的,它是一种克隆基因的新方法。文章利用电子克隆技术从玉米中成功得到MPK17基因,并对该基因及其编码蛋白进行了一系列生物信息学分析。经过初步分析,表明该目的基因长为2344 bp,编码579个氨基酸。所编码的蛋白质的分子量约为66.04 KDa,等电点为6.43,不存在跨膜区。该蛋白净带有六个负电荷,保守结构包括一些例如:ATP结合位点,底物结合位点,活性位点以及KIM停泊位点等。二级结构中,β折叠占15.20%;无规则卷曲占49.74%;α螺旋占35.06%。最后通过利用软件SWISS-MODEL对其进行三级结构的预测,找到与其匹配度最高的模板3py3A,从而预测出它的三级结构。需要注意的是,在实验过程中,笔者会采用不同的软件对目的蛋白进行分析,难免有时的结果不一定全部一致。这是因为不同的数据库,存放的数据有些差异,算法也有些差异,是属于正常的,基本上大部分都是可靠的。总之,以上种种分析都仅仅只是一个推测,还需要在实验室中进行进一步的研究论证。

图10 玉米MPK17基因编码蛋白电荷分布Fig.10 Change dist ribution of protein coded by corn MPK17 gene

图11 玉米MPK17基因编码蛋白质的保守域Fig.11 Conserved domain in protein coded by corn MPK17 gene
[1]袁月星,李巧丽.植物界中的“果蝇”——拟南芥[J].生物学教学,2003,28(1):48-49.
[2]陈璋.拟南芥:植物分子生物学研究的模式物种[J].植物学通报,1994,11(1):6-11.
[3]安贤惠.拟南芥及其研究发展[J].西北农业学报,1998,7(1):92-94.
[4]徐平丽,张传坤,孙万钢,等.模式植物拟南芥基因组研究进展[J].山东农业科学,2006,(6):100-102.
[5]Cai-Ping Feng,John Mundy.Gene discovery and functional analysis in the model plant Arabidopsis[J].Journal of integrative plant biology,2006,48(1):5-14.
[6]梁成真,张锐,郭三堆.染色体步移技术研究进展[J].生物技术通报,2009,(10):75-87.
[7]Shinozaki K G,Tena J,Sheen Y.Mitogen-activated protein kinase cascades in plants:a new nomenclature[J].Trends in Plant Science,2002,7(7):301-308.
[8]Mizoguch T,Ichimura K,Shinozaki K.Environmental stress response in plants:the role of mitogen-activated protein kinases[J].Trends Biotech,1997,(15):15-19.
[9]刘强,薛成.SHINOZAKI K.植物促分裂原活化蛋白(MAP)激酶[J].植物学报,2000,42(7):661-667.
[10] 吴涛,宗晓娟,谷令坤等.植物中的MAPK及其在信号传导中的作用[J].生物技术通报,2006,(5):1-7.
[11] 陈娅斐,冯斌,赵小明等.MAPK级联途径在植物信号转导中的研究进展[J].植物学通报,2005,(3):357-365.
[12]肖文娟,宾金华,武波.植物体中的 MAPK[J].植物学通报,2004,21(2):205-215.
[13]张成岗,贺福初.生物信息学方法与实践[M].北京:科学出版社,2002.
[14]张淼,赵茂林.利用表达序列标签电子克隆cDNA全序列的策略[J]. 生物技术通报,2010,(1):45-67.
[15]Arnold K,Bordoli L,Kopp J, et al.The SWISS-MODEL Workspace:A web-based environment for protein structure homology modelling.Bioinformatics,2006,(22):195-201.
[16]Schwede T, Kopp J, Guex N, et al.SWISS-MODEL:an automated protein homology-modeling server.Nucleic Acids Research,2003,(31):3381-3385.
[17] Guex N and Peitsch M C.SWISS-MODEL and the Swiss-PdbViewer:An environment for comparative protein modelling.Electrophoresis,1997,(18):2714-2723.
In Silico Cloning Analysis of Arabidopsis Thaliana MPK17 in Zea Mays
Wang Qi1,Wang Jun-hong2,Wang Rong3
(1.Department of Life Science and Technology Changzhi University,Changzhi Shanxi 046011;2.College of Life Sciences Shaanxi Normal University,Xi'an Shaanxi 210037;3.Changzhi Municipal Health School,Changzhi Shanxi 046000)
MPK is a Ser/Thr-type protein kinase which exists widely in each eukaryote.Nowadays,a lot of MPKs have been isolated in the plant.However,in the plant the MPK research mainly concentrates on the double seed leaf pattern plant (e.g.Arabidopsis thaliana),is very few about the single seed leaf pattern plant Zea mays'MPK research.We used Arabidopsis thaliana's MPK17 gene and then in silico cloned Zea mays'MPK17 gene using bioinformatics in the corn and moreover,carried on the preliminary analyses.It has provided the rich rationale for better research MPK17.
a rabidopsis thaliana;zea mays;mitogen activated protein kinase 17 (MPK17);in silico cloning
Q785
A
1673-2014(2012)02-0009-05
2011—10—18
长治学院校级课题基金资助项目(2010111)。
王 琦(1980—),男,山西长治人,硕士,讲师,主要从事生物化学、分子生物学、生物信息学研究。
(责任编辑 王璟琳)
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
国际放射医学核医学杂志(2020年4期)2020-07-27
中国洗涤用品工业(2019年4期)2019-05-11
生命科学研究(2018年1期)2018-05-29
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
上海农业学报(2017年3期)2017-04-10
动物医学进展(2015年10期)2015-12-07
小资CHIC!ELEGANCE(2015年15期)2015-09-01
