
半滑舌鳎三倍体诱导研究*
2012-01-10 09:35:04刘志鹏王旭波翟介明张全启
中国海洋大学学报(自然科学版) 2012年6期
刘志鹏,王旭波,翟介明,张全启**
(1.中国海洋大学海洋生命学院,山东青岛266003;2.莱州明波水产有限公司,山东烟台261400)
半滑舌鳎(Cynoglossus semilaevis)隶属于鲽形目(Pleuronectiformes)、舌鳎科(Cynoglossidae)、舌鳎属(Cynoglossus),俗称为鳎米、龙利等,为近海温水性大型底栖鱼类,主要在黄海、渤海区域。半滑舌鳎是一种名贵的海水鱼类,由于其肉鲜味美、营养丰富,深受广大消费者欢迎,拥有很高的经济价值和广阔的养殖前景[1]。随着近些年来我国北方兴起的比目鱼养殖热潮,半滑舌鳎已经成为我国海水鱼类养殖的主要品种之一。优良品种是养殖产业的基础,但目前对半滑舌鳎的养殖品种缺乏有效地选育,在养殖中的情况并不令人满意,因此改良现有种质资源,提高品种的繁殖、发育水平,进而培育出产量高、抗病强的新品种就成为了推动半滑舌鳎养殖产业发展的关键。
鱼类多倍体育种是鱼类遗传育种工作的重点研究领域之一,三倍体鱼类一般具有不育或低育性[2],可以在成鱼期将二倍体消耗在生殖腺发育的能量用于生长,克服性成熟带来的不利影响,减少由于过早成熟造成的产量和质量下降,因而能提高肉类质量和饵料转换效率[3-4],可被应用于大规格商品鱼培育,例如在加拿大,大规模的三倍体苗种养殖几乎遍布全国,他们用三倍体苗种抑制鱼类性成熟造成的死亡以及肉质下降[5],因此三倍体育种受到国内外水产科学工作者的广泛重视。本研究对半滑舌鳎三倍体的培育进行了尝试,使用冷休克法诱导得到了半滑舌鳎的三倍体,并筛选了最佳的诱导条件,为进一步开展半滑舌鳎细胞工程育种提供了基础数据。
1 材料与方法
1.1 实验用鱼
实验所用鱼取自莱州明波水产有限公司人工养殖的半滑舌鳎适龄亲鱼,选取性腺已经开始发育的健康亲鱼,雌鱼体重1.5~2.5 kg,雄鱼体重150~300 g,在循环水养殖车间养殖,控光控温进行催熟,性腺成熟后用GNRH组合催产剂进行催产。
1.2 人工授精
选取性成熟的雌雄鱼,通过挤压腹部的方法来采集卵子和精液。收集完毕后将精卵混合均匀,加入海水搅拌授精,并记录受精时间。
1.3 冷休克处理温度的确定
根据半滑舌鳎雌核发育[6]以及预实验得到的初步数据,设置2、3、4、5、6、7℃6个温度梯度,在受精后4~5 min,将盛有受精卵的筛网放入各梯度海水中进行冷休克处理,持续时间为20~25 min。每组处理完毕后各取1 000粒受精卵放入单独的筛网中在23℃的海水里进行孵化,另取1 000粒未处理的受精卵作为对照组。及时统计各组的死卵,统计孵化率并在仔鱼孵出后统计三倍体率。实验重复3次。
1.4 倍性检测
参考杨景峰的方法并做稍许改进,在仔鱼孵化2 d后,取30尾分别加一步法试剂,粉碎鱼体解离细胞,用200目的筛绢过滤细胞后,用流式细胞仪进行分析,二倍体对照组初孵仔鱼的DNA相对含量设为100[7]。
1.5 最佳起始时间和持续时间的筛选
在确定最佳冷休克处理温度后,将起始时间分为受精后3、4、5、6、7 min 5个梯度,在最佳水温下每个梯度持续处理15、20、25、30、35 min。统计孵化率并在仔鱼孵出后统计三倍体率。
1.6 染色体鉴定
在鱼苗发育至体长10~15 cm左右的时候,参考周丽清的方法进行染色体制片并做稍许改进[8]。从鱼鳃盖下沿的腹腔部注射PHA溶液(用0.7%的NaCl配制,终浓度为5 mg/m L)处理3 h,然后在同一位置注射秋水仙素溶液(用0.7%的NaCl配制,终浓度为1 mg/m L)处理1 h,将鱼放血取出头肾,在生理盐水中进行解离并过滤到离心管中,1 000 r/min离心8 min并洗涤细胞,接着在浓度为0.075 mol/L的KCl中低渗处理30 min后再离心8 min收集细胞,最后用新配制的预冷卡诺固定液(无水乙醇∶冰醋酸=3∶1)固定3次,每次不少于30 min。滴片时采用冷滴片法,用10%的Giemsa染色30 min,显微镜下观察中期分裂相并拍照,观察50尾个体。
2 结果
2.1 最佳冷休克温度
在受精后4~5 min,处理20~25 min的相同条件下,6个不同的冷休克处理温度与半滑舌鳎平均三倍体率的关系如图1所示,在3~6℃的温度下都有三倍体仔鱼孵化,其中4~5℃时三倍体率最高,在此将4℃确定为最佳处理温度,此时三倍体率最高为27.8%,孵化率为5.3%。
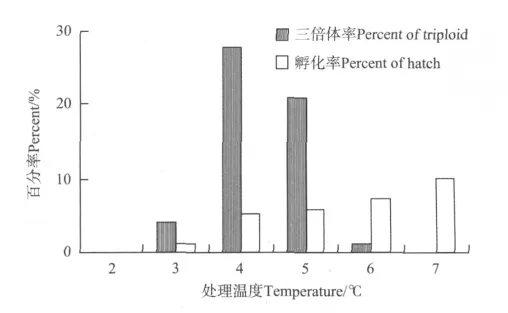
图1 不同冷休克温度与半滑舌鳎三倍体诱导率的关系Fig.1 The relationship between the yield of triploid and the temperature of cold shocks in C.semilaevis
2.2 最佳起始时间和处理时间
在根据上述实验得到的最佳处理温度4℃下,起始时间和持续时间与平均三倍体率(3n)、平均孵化率(H)的关系如表1所示。各处理组的孵化率均不高,在起始时间相同的情况下,处理时间越长,孵化率越低,受精后3和35 min处理时间的实验组孵化率为0,而在相同的处理时间下,孵化率与起始时间没有特别明显的关系。各处理组的三倍体率也是非常低,其中以受精后5 min为处理起始时间,20 min为持续处理时间获得的三倍体率最高,虽然在此条件下受精卵的孵化率不是最高的,但考虑到本研究中孵化率都比较低,因此最佳起始时间为受精后5 min,最佳处理时间为20 min。
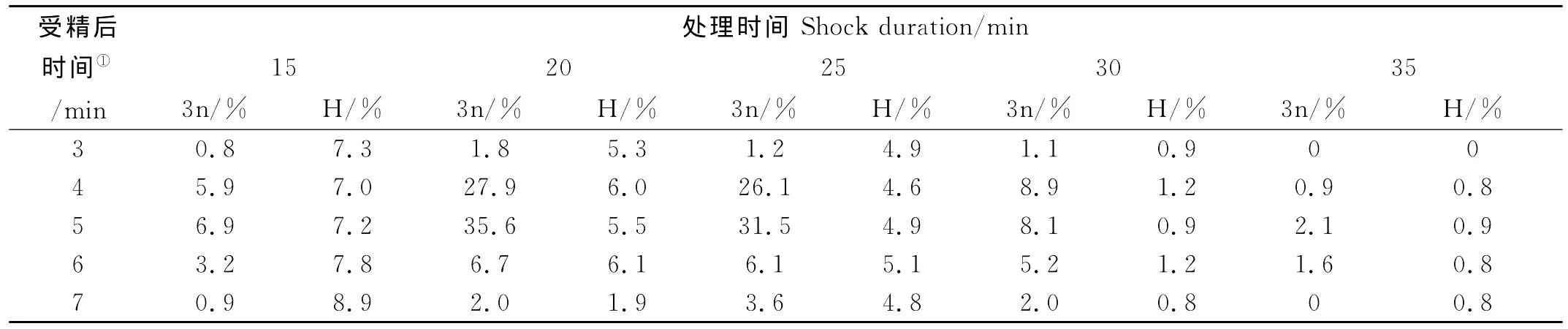
表1 不同处理起始时间和持续时间对半滑舌鳎三倍体诱导效果Table 1 The triploidy induction result of different starting time and shock duration
2.3 染色体鉴定
染色体观察结果(见图2,图中箭头所示为W染色体,标尺为5μm)表明,未经处理的正常二倍体染色体核型为2n=42,为ZZ(见图2A)和ZW(见图2B),经过冷休克处理的三倍体染色体核型为3n=63,其性染色体有2种类型,分别为ZZZ(见图2C)和ZWW(见图2D),证明了它们确实是半滑舌鳎的三倍体。
3 讨论
3.1 诱导方法
人工诱导鱼类三倍体常用的有生物、物理、和化学3种方法,冷休克是一种物理方法,因为其操作简便,处理量大目前被广泛应用。用冷休克诱导三倍体需要确定3个重要因素:处理开始时间、处理水温以及持续时间,实验也表明半滑舌鳎三倍体诱导明显受到这3个因素的影响。
鱼类三倍体诱导的原理是抑制受精卵第二极体出,在受精卵中多保留一套母本染色体而成为三倍体,所以冷休克起始时间必须在第二极体排出之前。有的研究把卵从第二次成熟分裂中期到第二极体排出的这段时间划分为启动期、敏感期和不应期3个阶段[9],在敏感期内适当的处理就能将第二极体保留在受精卵内,当从敏感期进入不应期后外界处理对抑制第二极体排出失去作用,无法产生三倍体。在本研究中,半滑舌鳎受精卵在受精后5 min诱导的效果最好,说明受精卵正处于敏感期,是抑制第二极体排出的最佳时期,而起始时间在受精7 min以后的三倍体率非常低,受精卵已经进入了不应期。大部分鱼类的三倍体诱导起始时间为3~10 min[10],与本研究结果相符。第二极体的排出时间也与受精水温、卵子成熟程度等有很大关系,因此在确定处理起始时间时还应考这些方面的影响。
冷休克处理温度不应过低或过高,最合适的温度应该是其亚致死温度[11],过低的温度会超过受精卵承受的极限,使受精卵大量甚至全部死亡,而过高的温度又起不到诱导的效果。在本研究中,当处理温度低于4℃时,胚胎大量死亡,三倍体率也明显下降,而5和6℃进行冷休克的三倍体率也比使用4℃进行处理的三倍体率低,说明在亚致死温度以内,温度越低对第二极体释放的抑制作用越强。
在起始时间和处理温度都确定的情况下,处理时间会直接影响受精卵的诱导率和孵化率,本研究中处理20 min时诱导率最高,但随着处理时间的延长,诱导率和孵化率都开始下降,直到受精卵全部死亡,过长的处理时间会对受精卵造成严重损伤。需要注意的是,同一种鱼不同的人用相同的方法进行多倍体诱导也会出现不同的结果,这可能与试验用鱼的地理种群、养殖工艺、性腺成熟度、倍性鉴定方法等因素有关。
3.2 诱导率和孵化率
在本实验中,三倍体率与孵化率之间没有明显的相关性,例如在处理温度较高的6和7℃中或是10 min等处理强度较低的实验组中,低强度冷处理对受精卵的发育和分化影响较小,受精卵的孵化率比较高,但在这些实验组中的三倍体率却很低甚至为0,而在最佳处理条件下进行处理时其诱导率虽然是最高的,但其孵化率却不是最高的。
本研究验证了冷休克对半滑舌鳎三倍体的诱导是有效的,在初孵仔鱼期的最高三倍体率达到35%,但与在黑鲷(50%)[12]、大菱鲆(70%)[13]等冷休克诱导的三倍体率相比,半滑舌鳎的诱导率还是比较低。未经处理的对照组受精卵孵化率在50%左右,而在经过筛选的最佳处理条件下(受精后5 min,4℃下处理20 min)冷休克后的孵化率只有5.5%,经观察,发现在冷休克处理后,胚胎受到了较大程度的损伤,到原肠期阶段就无法继续发育,导致孵化率非常低。与兴国红鲤(50%~70%)[14]、红鳍东方鲀(80%)[15]等进行冷休克诱导三倍体的孵化率相比,半滑舌鳎的孵化率明显偏低,这可能是因为半滑舌鳎的受精卵对低温损伤更为敏感,所以造成了其孵化率大大低于别的鱼类。
3.3 三倍体半滑舌鳎的性染色体类型
半滑舌鳎的雌性个体中被发现含有1条巨大的异形染色体,经推测为W性染色体,进而推测半滑舌鳎的性别决定类型是ZZ/ZW型,雌性为配子异型ZW,雄性为配子同型ZZ[8,16]。由此可以推测出通过抑制受精卵第二极体释放而形成的三倍体,其染色体类型应该为ZZZ和ZWW。本研究中进行的半滑舌鳎三倍体个体的染色体制片,经过镜检后确实发现了这2种类型的染色体,且其比例为1∶1,未发现ZZW型的三倍体个体。这一结果说明在半滑舌鳎雌鱼减数分裂中,Z染色体和W染色体之间的交叉互换被严重抑制,半滑舌鳎雌雄个体之间巨大的形态差异,除了与W染色体“多余”的遗传信息相关外,可能与ZW染色体之间的遗传信息交流受阻有关,而这种交换的机制也恰好是性染色体起源与进化的必然途径。
[1] 李思忠,王惠民.中国动物志,硬骨鱼纲、鲽形目[M].北京:科学出版社,1995:94-98.
[2] 桂建芳,肖武汉.人工三倍体水晶彩卿的性腺发育[J].动物学报,1991,37(30):297-304.
[3] 尤锋,沙学绅.黑鲷三倍体人工诱导的初步研究[J].海洋与湖沼,1991,22(5):489-491.
[4] Benfey T J,Sutterlin A M.Triploidy induced by heat shock and hydrostatic pressure in landlocked Atlantic salmon(salmon salar L)[J].Aquaculture,1984,36:359-367.
[5] Benfey T J.Use of all-female and triploid salmonids for aquaculture in Canada[J].Bulletin of the Aquaculture Association of Canada,1996,96(2):6-8.
[6] Chen Song Lin,Tian Yong Sheng,Yang Jing Feng,et al.Artificial Gynogenesis and Sex Determination in Half Smooth Tongue Sole(Cynoglossus semilaevis)[J].Marine Biotechnology,2008,11(2):243-251.
[7] 杨景峰,陈松林,徐亘博,等.异源精子诱导犬齿牙鲆的雌核发育[J].水产学报,2009,33(4):533-540.
[8] 周丽青,杨爱国,柳学周,等.鳎染色体核型分析[J].水产学报,2005,29(3):417-419.
[9] 桂建芳,肖武汉,梁绍昌,等.静水压休克诱导水晶彩鲫三倍体和四倍体的细胞学机理初探[J].水生生物学报,1995,28(2):49-55.
[10] 楼允东.鱼类育种学[M].北京:中国农业出版社,1999:122-126.
[11] 尤锋.海产鱼类多倍体育种研究[J].海洋科学,1997(1):33-36.
[12] 尤锋.黑点三倍体的人工诱导研究[J].海洋与湖沼,1993,24(3):248-255.
[13] Francesc Piferrer,Rosa M-Cal,Blanca A lvarez-Blazquez,et al.Induction of triploidy in the turbot(Scophthalmus maximus)Ⅰ.Ploidy determination and the effects of cold shocks[J].Aquaculture,2000,188:79-90.
[14] 洪一江,胡成钰.人工诱导兴国红鲤三倍体最佳诱导条件的研究[J].动物学,2000,35(4):2-4.
[15] 王茂林,姜志强,李荣.红鳍东方鲀三倍体诱导的初步研究[J].水产科学,2006,25(7):349-351.
[16] Zhuang Z M,Wu D,Zhang S C,et al.G-banding patterns of the chromosomes of tonguefish Cynoglossus semilaevis Günther 1873[J].Journal of Applied Ichthyology,2006,22:437-440.
猜你喜欢
中国水产(2024年3期)2024-04-11 07:36:00
天津农学院学报(2024年1期)2024-04-07 11:20:22
水产学报(2024年3期)2024-03-25 07:43:22
中国蚕业(2022年1期)2022-03-09 02:59:20
科学养鱼(2021年11期)2022-01-11 11:21:10
今日农业(2021年6期)2021-11-27 08:05:59
蚕桑通报(2020年3期)2021-01-07 03:05:20
食品与生活(2019年12期)2019-05-15 15:01:41
生命科学研究(2018年1期)2018-05-29 01:12:56
动物营养学报(2015年10期)2015-12-01 02:26:34
