
猕猴桃ACC氧化酶反义基因转化猕猴桃的研究
2012-01-08 11:06田宏现王曼玲夏新界谭晓风
中南林业科技大学学报 2012年11期
田宏现 ,苑 平 ,王曼玲 ,刘 艺 ,李 菁 ,夏新界 ,谭晓风
猕猴桃ACC氧化酶反义基因转化猕猴桃的研究
田宏现1,2,苑 平2,王曼玲3,刘 艺2,李 菁2,夏新界3,谭晓风1
(1.中南林业科技大学,湖南 长沙 410004;2.吉首大学,湖南 吉首 416000;3.中国科学院 亚热带农业生态研究所 , 湖南 长沙 410125)
为建立猕猴桃ACC氧化酶反义基因功能研究技术平台并通过生物技术改良猕猴桃,从海沃德猕猴桃(Actinidia chinensis Planch.var.deliciosa‘Hayward’)叶片中提取总RNA,反转录合成cDNA第一链,并以此为模板,扩增得到ACC氧化酶(ACO)基因片段约1 kb,经限制性内切酶图谱分析,组建亚克隆并进行测序,其开放阅读框长957 bp,编码319个氨基酸,与其他已公布的ACC氧化酶cDNA的核苷酸(AB003514.1)及氨基酸序列(BAA21541.1)同源率分别高达94.0 %和95.6 %,证明所克隆的cDNA序列的准确性。构建该基因的反义表达载体,并通过农杆菌介导的方法成功转化米良一号猕猴桃(Actinidia deliciosa cv.Miliang-1),得到再生苗。取14株经PCR检测,其中10株检测到ACC氧化酶基因目的条带,阳性植株占71.4 %。实时荧光定量PCR分析结果显示,再生苗ACO基因的表达量低于野生型58%~81%。
猕猴桃; ACC氧化酶;农杆菌介导;反义表达载体
米良1号是吉首大学在上世纪80年代选育出的美味猕猴桃优良品种,它树势强健,适应性强,抗旱抗病虫,果大丰产稳产,深受果农欢迎;其肉质细嫩、风味纯正、酸甜适度、清香宜人,加上维生素c、总糖、可溶性固形物含量均高于海沃德猕猴桃,颇受消费者青睐[1]。但采后常温下贮运月余开始软腐,失去其应有的经济价值,在贮运和鲜销上有很大的困难,急需对其进行遗传改良。猕猴桃的品种很多,有数十种,但都有贮存性能急需进行遗传改良的问题。例如新品种红阳A .chinensis Planch. var. chinensis,相对其他品种来说,口感最好,但贮藏矛盾就更突出。研究表明,猕猴桃果实属呼吸跃变型果实,呼吸跃变高峰与乙烯的产生相关[2-4],器官中乙烯合成量的增加将引起果实呼吸强度增加,导致果实中多聚半乳糖酸酶、淀粉酶、过氧化物酶、脂氧合酶、多酚氧化酶等代谢酶的活性升高,使细胞膜透性升高、细胞结构破坏、果实硬度下降,加速果实软化,贮藏时间变短。乙烯在植物体内的合成是从甲硫氨酸开始的,甲硫氨酸在S-腺苷转移酶作用下转变成S-腺苷甲硫氨酸,然后S-腺苷甲硫氨酸由1-氨基环丙烷-1-羧酸合成酶(ACC合成酶)催化形成1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid, ACC),最后,ACC在ACC氧化酶(ACC oxidase, ACO)催化下形成乙烯[5]。乙烯在植物体内的生物合成主要是由ACC合成酶和ACC氧化酶这两个限速酶来控制的,通过调节它们的表达能有效地调节乙烯的合成量[6]。ACC氧化酶催化ACC转化为乙烯,是乙烯生物合成途径中的最后一个酶,其活性高低直接决定植物体内乙烯生成量的多少[7]。目前,番茄、卡特兰、酸橙、富有柿果、香蕉、梨、苹果等中的ACC氧化酶基因均已经被克隆并测序[8]。由于核糖体不能翻译双链的RNA,所以ACC氧化酶的mRNA与和该mRNA有特异性碱基互补关系的反义RNA结合为双链后,即抑制了该mRNA的翻译。根据ACC氧化酶基因序列,人工合成反义RNA基因,并将其导入细胞内转录成反义RNA,即能抑制ACC氧化酶基因的表达,减少或阻断果实中乙烯的合成,大大延长呼吸跃变型果实采收后的贮藏保鲜时间。1990年,Hamilton等[9]将ACO基因的反义基因导入番茄后,实现了对番茄果实乙烯生成和成熟的控制。Picton等[10]将ACO基因的反义RNA导入番茄后,转基因番茄果实的乙烯含量比对照组下降了90%,成熟时间达到了3个星期才能变红,而对照组只有7 d。猕猴桃有人做过ACC氧化酶基因克隆和测序[11-12],但构建猕猴桃ACO基因的反义表达载体,通过农杆菌介导转化猕猴桃,并获得阳性再生苗的研究还未见报道。本文成功地构建了猕猴桃ACO基因的反义表达载体,并通过农杆菌介导的方法,转化米良一号猕猴桃,经过PCR检测和实时荧光定量PCR分析证实获得了转基因植株。
1 材料与方法
1.1 材 料
1.1.1 海沃德猕猴桃幼苗和米良一号猕猴桃枝条
海沃德猕猴桃A. chinensis Planch.var.deliciosa‘Hayward’幼苗由吉首大学石泽亮教授惠赠的海沃德猕猴桃果实收集种子,经低温砂藏后种植而得。受体植物米良一号猕猴桃(A.deliciosa cv.Miliang-1 Shi Ze-liang)枝条由石泽亮教授赠送。
1.1.2 试剂
胶回收试剂盒选购于AxyGen公司,cDNA合成试剂盒、TA克隆试剂盒、LA Tap酶、Trizol等购于Takara公司,其它试剂为国药集团产品。
1.1.3 菌种和质粒
根癌农杆菌菌株为EHA105,质粒为pCAMBIA1300-163,由中科院亚热带农业生态研究所提供。该质粒T-DNA区结构如图1,其中含有潮霉素磷酸转移酶基因(hpt)。

图1 植物表达载体pCAMBIA1300-163的T-DNA区Fig.1 Schematic T-DNA zone of plant representative plasmid pCAMBIA1300-163
1.1.4 培养基
诱导培养基:MS + 10 mg·L-16-BA + 0.2 mg·L-1NAA + 0.2 mg·L-1TDZ + 30 g·L-1蔗糖 + 7.5 g·L-1琼脂,pH 6.0;
继代培养基:MS + 0.25 mg·L-16-BA + 1 mg·L-1NAA + 30 g·L-1蔗糖 + 7.5 g·L-1琼脂,pH 6.0;
共培养基:MS + 0.25 mg·L-16-BA + 1 mg·L-1NAA + 30 g·L-1蔗糖 + 0.1 mmol·L-1乙酰丁香酮 +7.5 g·L-1琼脂,pH 5.6;
筛选培养基:MS + 0.25 mg·L-16-BA + 1 mg·L-1NAA + 500 mg·L-1头胞霉素 + 400 mg·L-1羧苄青霉素 + 20 mg·L-1潮霉素 + 30 g·L-1蔗糖 + 7.5 g·L-1琼脂,pH 6.0;
预分化培养基:MS + 3.0 mg·L-1ZT + 0.5 mg·L-16-BA + 0.05 mg·L-1NAA + 30 g·L-1蔗糖 + 400 mg·L-1羧苄青霉素 + 7.5 g·L-1琼脂,pH 6.0;
分化培养基:MS + 1.0 mg·L-1ZT + 30 g·L-1蔗糖+ 400 mg·L-1羧苄青霉素 + 7.5 g·L-1琼脂,pH 6.0;
生根培养基:1/2 MS + 0.7 mg·L-1IBA + 0.15 mg·L-16-BA + 15 g·L-1蔗糖 + 7.5 g·L-1琼脂,pH 6.0。
1.2 方 法
1.2.1 猕猴桃RNA的提取
RNA提取缓冲液于65℃水浴锅中温育30 min以上,然后在缓冲液中加入2%~5% β-ME,立即取新鲜嫩叶加缓冲液于研钵中快速研磨(0.1 g新鲜嫩叶/ 1 mL缓冲液)。把研磨好的,成粘稠状的匀浆液移入离心管中,65℃温育10 min,加入等体积氯仿∶异戊醇(24∶1),上下剧烈震荡15 s,然后于4℃,12 000 g,离心10 min。取上清,重复上述抽提步骤2~3次。取上清,加入1/4体积的10 mol LiCl溶液并混匀,-20 ℃条件下放置12 h以上,使RNA沉淀。4℃,12 000 g,离心20 min,弃上清,获粗提的总RNA沉淀。加入DEPC处理过的H2O重新溶解RNA。再加入1/10体积的3 mol NaAc,2.5体积的96 %乙醇。置于-20℃,1 h,使RNA沉淀。4℃,12 000 g,离心20 min,弃上清,用75 %乙醇清洗沉淀,空气中干燥后加入DEPC处理过的H2O溶解,保存于-70℃备用(也可直接用无水乙醇保存)。
1.2.2 cDNA第一链的合成
取1 μg mRNA作模板,于20 μL反应体系中加入 7 μL(约 1 μg)mRNA、1 μL Oligo(dT)18 primer、4 μL DEPC-treated Water,65℃水浴 5 min,冰上放置 3~ 4 min。接着加入 4 μL 5×reaction buffer、1 μL RiboLock ™ RNase Inhibitor (20 u/μL)、2 μL 10 mmol dNTP mix、1 μL RevertAid ™ M-MuLVReverse Transcriptase (200 U/μL),42℃水浴 60 min,70℃水浴5 min终止反应。cDNA可在-20℃暂时保存不超过一个星期,-70℃长期保存。
1.2.3 PCR扩增
引物设计参照海沃德的基因序列,使用Primer Expression 5.0软件设计PCR引物,由invitrogen公司合成。上游引物,5′-CCCGGGAGAGAGAT GGAGGCTTTCCCA-3′; 下 游 引 物,5′-GTCGAC TCACACAAACACAAGCGGCG-3′。 以 cDNA 为模板进行PCR,扩增反应条件:94℃ 5 min;94℃30 s,56℃ 45 s,72℃ 1.2 min,35 cycles;72℃ 10 min;22℃保温10 min。PCR反应使用LATaq酶。
1.2.4 PCR扩增产物的克隆与序列分析
PCR扩增产物经琼脂糖凝胶电泳分析后,回收特异片段,连接到载体pMD18-T Vector当中,转化大肠杆菌DH10B感受态细胞。在选择培养基上挑取单菌落摇菌,用碱法少量提取质粒并做酶切鉴定。将阳性克隆送华大基因测序。
1.2.5 ACC氧化酶反义表达载体的构建
从海沃德猕猴桃中克隆得到ACC氧化酶基因的cDNA片段,测序确定为所需片段后将其反向连入表达载体pCAMBIA1300 -163当中,得到猕猴桃ACC氧化酶基因的反义表达载体。
1.2.6 遗传转化
组织培养:米良一号猕猴桃愈伤组织和无菌苗的培养,参考田宏现等的方法[1]。
农杆菌与愈伤组织的共培养:取EHA105(含pCAMBIA1300-163 质粒 )划 LB 板 (加 Kan 50 mg·L-1和CHL 34 mg·L-1),28℃培养2 d后挑单菌落,在LB板划线,28℃过夜培养,用液体共培养基将菌洗下,调OD600=0.6;挑选表面干爽、结构致密的愈伤组织浸入上述菌液中,每隔5 min摇一次,浸泡30 min后用灭菌滤纸吸干愈伤组织表面多余菌液,将愈伤转移到固体共培养基上,25~26℃下暗培养3~4 d。
抗性愈伤组织的筛选:将共培养3 d后的愈伤移入灭菌三角瓶,无菌水冲洗5次至液体不浑浊,再用含有 500 mg·L-1头胞霉素和 400 mg·L-1羧变青霉素的无菌水浸泡30 min,其间每隔5 min摇一次。用灭菌滤纸吸干水分,转移到筛选培养基上,25~26℃暗培养20 d后,将抗性愈伤转移到新的筛选培养基上继续培养。
抗性愈伤组织的分化:两次筛选后,将抗性愈伤转移至预分化培养基上,置于25~26℃,光强1000~1500 lx,每天14 h光照培养,每20 d更换一次新培养基。当愈伤开始变成红褐色并开始分化出芽时将愈伤转移至分化培养基上。
抗性植株的生根:待苗长至3~5 cm高时,切下转移到生根培养基上,置光照培养箱中,条件同预分化。
抗性植株的炼苗:挑根多而粗壮的抗性植株,室内拧松瓶盖3 d后用自来水将幼苗根上的培养基冲洗干净,移栽到装有基质(土∶珍珠岩体积比=3∶1)的盆子,置于置物箱中室内炼苗2周后打开置物箱,室内炼苗3 d,移至室外7d即可移入实验田中。
1.2.7 抗性植株的PCR检测
经过转化获得29株潮霉素抗性植株,随机选取其中14株提取基因组DNA,进行PCR扩增。取抗性猕猴桃植株幼嫩叶片0.2~1.0 g,采用CTAB区室法提取基因组DNA。扩增产物为575 bp的基因片段。上游引物为Term F:5′-GGTGCTTTCCCAGTCCAAATCGTT-3′。 下 游引物为 Term R:5′-CCAGGTTTAGTCGTCTCGTG TCTGGT-3′。
PCR扩增体系(20 μL):Takara公司TaqDNA聚合酶 (5 mmol·μL-1) 0.2 μL,10×Buffer (KCl)缓冲 液 2.0 μL,MgCl2 1.6 μL,dNTP Mixture 2.0 μL,Term F(20 mmol·μL-1) 0.4 μL,Term R(20 mmol·μL-1) 0.4 μL,模板 1 μL,ddH2O 12.4 μL。
PCR反应程序为:95℃ 5 min;94℃ 35 s,59℃ 35 s,72℃ 45 s;35 cycles;72℃ 10 min。取PCR产物10 μL进行琼脂糖(1.0 %)凝胶电泳检测。
1.2.8 抗性植株的实时PCR检测
由于猕猴桃属于木质藤本,组培苗要3~5年才能结果,果实中靶基因表达结果的确定还有一段长时间的等待。故随机挑选5株经PCR鉴定为阳性的再生转基因猕猴桃植株和一株野生型米良一号植株,剪其叶片进行荧光定量实时PCR检测,检测靶基因的表达丰度。为猕猴桃ACO基因的反义基因导入后获得的再生植株的定性提供进一步的间接依据。提取猕猴桃总RNA[13],用Fermentas公司的DNase处理以去除混入的基因组DNA(按说明书操作)。采用invitrogen公司的PlatinumR SYBRR Green qPCR SuperMix-UDG(C11733-038)及 CFX96™Real-Time PCR Detection System荧光定量PCR仪进行PCR产物实时荧光检测。使用AlleleID 5软件设计引物,目的基因引物为ACO-F:5′-CGAGGACACGGAGGGTAGTTG-3′,ACO-R:5′-ACTGGACCTGCTATGTGAGAACG-3′,扩增片段长117 bp;内参基因为Action,其特异性引物为Action-F:5′-GCATCCCTCAGCACCTTTCAAC-3′,Action-R:5′-AAGCAGTCGCATAACTCAGAAGC-3′,扩增片段长105 bp。热循环设置:48℃30 min;95 ℃ 3 min;95 ℃ 10 s,60 ℃ 20 s,40 cycles。
2 结果与分析
2.1 猕猴桃总RNA的提取
提取的猕猴桃总RNA经1 %琼脂糖凝胶电泳检测,可见28 S、18 S两条清晰的rRNA条带,且两条带亮度比约为2∶1,表明总RNA未出现降解,完整性较好。紫外分光光度计测定的A 260 nm/A 280 nm比值在1.9~2.0之间,表明其纯度较高,无蛋白和DNA污染(图2)。
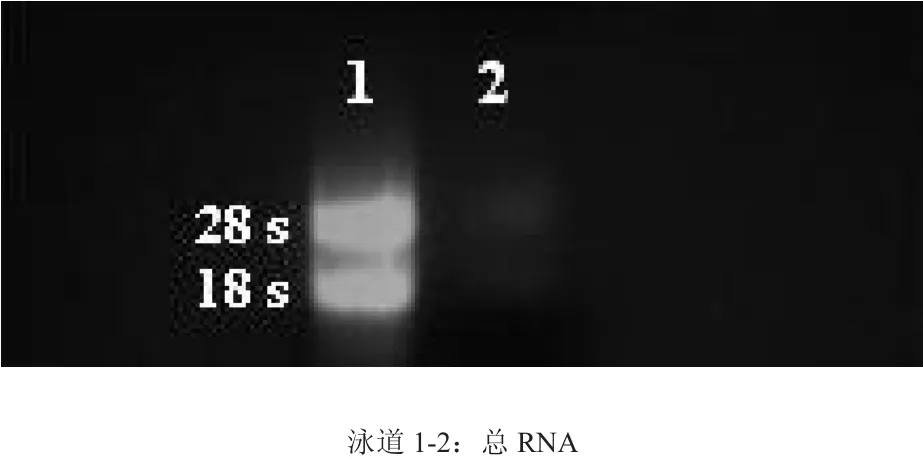
图2 猕猴桃总RNA电泳图Fig. 2 Electrophoregram of total RNA of kiwifruit
2.2 ACC氧化酶cDNA的PCR扩增与全序列测定
用高保真的Taq酶成功的克隆得到1 kb左右的ACC氧化酶cDNA目的条带(图3)。
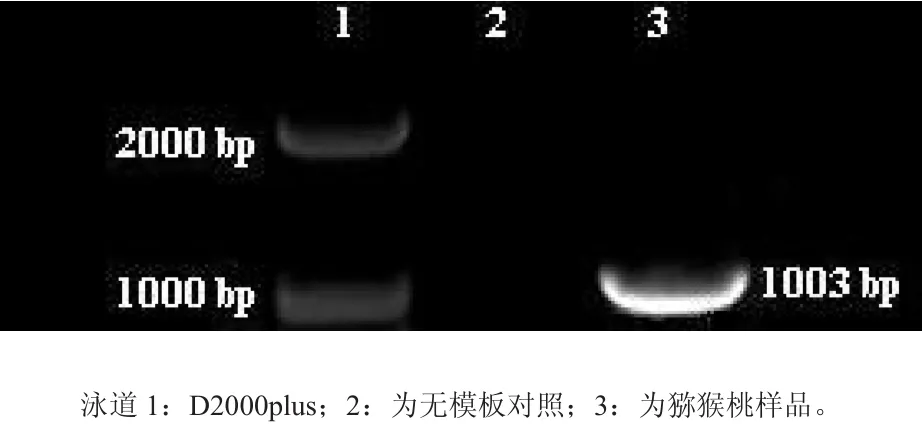
图3 PCR产物琼脂糖凝胶电泳分析Fig. 3 Electrophoretic analysis of agarose gel of PCR products
测序分析显示此cDNA全长为1003 bp。通过Vector Nti Explorer分析该基因的开放阅读框(ORF)为957 bp,编码一条319个氨基酸残基的多肽,经Blastn分析表明与设计引物时所用的国外报导的海沃德核苷酸序列(AB003514.1)有11个碱基不同,相似性为94 %,氨基酸序列中有2个氨基酸不同(见图4)。从而证实克隆所得确为猕猴桃ACC氧化酶基因的cDNA,已提交到GenBank当中,登陆号为JQ062390。
2.3 ACO基因反义表达载体的构建
用SmaⅠ、SalⅠ做双酶切鉴定构建完成的反义表达载体,目的片段为1015 bp(如图5)。由酶切结果得出,我们成功的将ACO基因反向的连入中间载体pJIT163,并成功的从pJIT163上将启动子、目的基因和终于止一起切下,连入表达载体pCAMBJA1300。
2.4 遗传转化
茎段愈伤诱导以继代培养基为好,愈伤结构致密,颜色较深。叶片诱导以诱导培养基为好,接种7 d开始出愈伤,叶片愈伤诱导率为100%(图6-A)。
继代:培养基选用继代培养基,20 d左右继代一次。愈伤组织结构致密,颜色翠绿,基本上不会出现褐化,但当继代时间超过30 d时,愈伤结构开始变疏松,颜色变白,后期分化效率变低(图6-B)。
筛选抗性愈伤:以继代培养基为基础,在其中添加20 mg·L-1潮霉素用于筛选抗性愈伤较好,愈伤的存活率为43.3%。抗性愈伤刚开始时为白色,添加头孢霉素和羧变霉素能有效抑制农杆菌的生长(图6-C)。

图4 猕猴桃 ACO cDNA 序列及推导出的氨基酸序列Fig. 4 Sequences of ACO cDNA and its deduced protein

图5 表达载体的Sma I和Sal I双酶切鉴定Fig.5 Estion result of SmaI and SalI of expression vector
分化:将抗性愈伤接到ZT的浓度加大到3.0 mg·L-1的预分化培养基上继代培养可加速芽的诱导。在预分化的过程中逐渐变成绿色,当愈伤开始变成红褐色时,再将愈伤接到ZT浓度降至1.0 mg·L-1分化培养基上,开始分化出芽(图6-D)。
生根:抗性愈伤分化芽接到1/2 MS + 0.7 mg·L-1IBA + 0.15 mg·L-16-BA的生根培养基上,20 d左右诱导出根,根比较粗状,平均每个芽10条根左右(图6-E)。
炼苗:经炼苗后将其释放到环境中去,存活率可达到90%以上(图6-F)。
2.5 抗性植株的PCR检测
抗性植株ACC氧化酶基因的PCR检测结果如图7。结果显示14株中有10株扩出目的条带,为阳性植株,阳性植株率为71%。目的条带大小为575 bp,抗性植株中10株的扩增片段大小与阳性对照的条带一致,而阴性对照(水和非转基因米良一号植株)没有目的条带,证明目的基因已整合到转化米良一号的基因组中。
2.6 实时荧光定量PCR分析
所得数据按相对定量法进行定量计算,目的基因相对表达量计算公式为Rel.Exp=2ΔΔCt,其中Ct值为阈值循环数(Threshold cycle),ΔΔCt=(未知样品ΔCt)-(参比样ΔCt),未知样品ΔCt=(内参基因Ct)-(目的基因Ct),参比样ΔCt=(参比样内参基因Ct)-(参比样目的基因Ct)[14](如图8)。
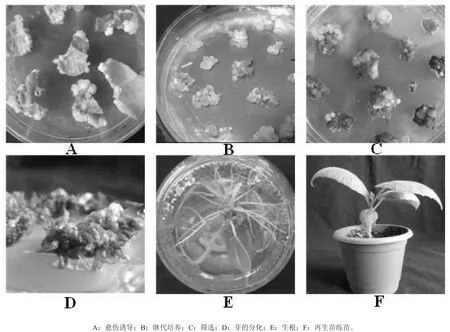
图6 米良一号农杆菌介导的遗传转化体系Fig.6 Genetic transformation of Miliang-1 by Agrobacterium-mediated method
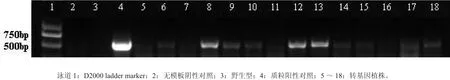
图7 转化植株的PCR分析Fig.7 PCR analyses of transformed plants
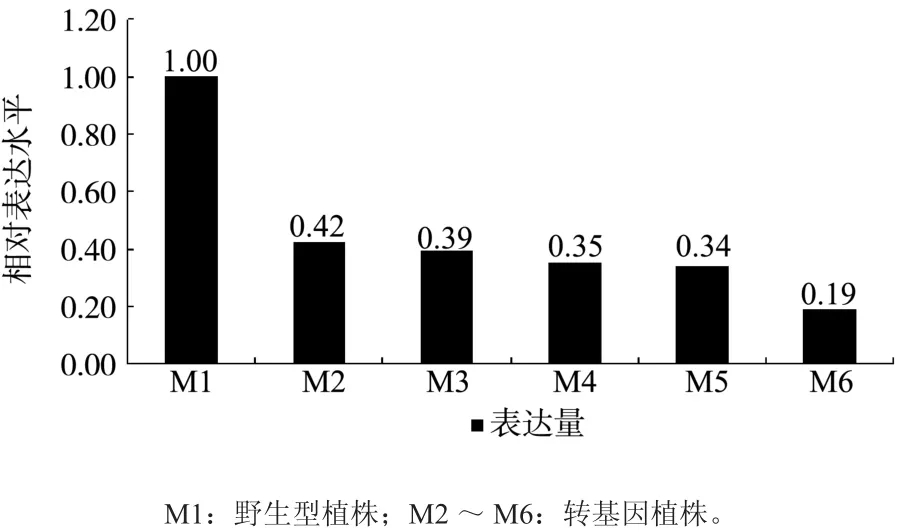
图8 转化植株的real time PCR分析Fig. 8 Real time PCR analyses of transformed plants
由图可看出将ACO基因在M1(野生型对照)植株中的相对表达量定为1,M2~M6的相对表达量比野生型要低58%~81%。
3 结论与讨论
本次研究中以海沃德猕猴桃的幼苗为材料,从中提取总RNA,然后反转录成cDNA,并克隆得到猕猴桃ACO基因。测序后得出此基因全长1003 bp,开放阅读框(ORF)为957 bp,编码一条319个氨基酸残基的多肽,经Blastn分析表明与设计引物时所用的国外报导的海沃德核苷酸序列(AB003514.1)有11个碱基不同,相似性为94 %,氨基酸序列中有2个氨基酸不同,进一步证实了我们所克隆得到的为猕猴桃ACO基因。并成功提交到GenBank当中登陆号为JQ062390。
成功克隆猕猴桃的ACO基因后,拟通过反义RNA技术来抑制猕猴桃乙烯的生物合成,达到延迟猕猴桃成熟的效果,成功构建了ACO基因的反义表达载体。并通过农杆菌介导的方法成功转化猕猴桃,所得抗性植株经PCR检测,证明本研究得到了猕猴桃ACC氧化酶转基因再生苗。挑选5株经PCR鉴定为阳性的红阳猕猴桃植株进行实时荧光PCR检测,结果表明ACO基因在转基因植株中的相对表达量比野生型降低58%~81%。成功的降低了ACO基因在猕猴桃中的表达活性,进一步验证了该实验结果的可靠性。ACO基因在转基因植株内表达的差异可能与插入的拷贝数和插入位点有关。
农杆菌介导转化法是一个细菌和愈伤组织共同作用的复杂过程。凡是涉及到细菌活性或愈伤组织状态的因素都可能影响转化效果。对米良一号而言,一方面是愈伤分化较难,另一面是潮霉素筛选时对愈伤组织造成伤害,潮霉素浓度过高时愈伤完全不能生长。筛选时培养基中合适的潮霉浓度是筛选成功的关键所在。目前仍未见农杆菌介导ACC氧化酶反义基因转化猕猴桃获得再生苗的报道。通过本实验的研究,找到合适的诱导、继代、分化培养基并优化了生根条件,为农杆菌介导转基因猕猴桃提供合适的组培体系。另一方面,本实验通过对潮霉素浓度的摸索,找到了一个适合用于米良一号筛选的潮霉素浓度,即在继代培养基中添加20 mg/L的潮霉素用于抗性愈伤的筛选。并对侵染时农杆菌的浓度,愈伤的处理以及共培养的条件等进行了优化且取得了较为理想的效果。
米良一号猕猴桃属于多糖多酚类植物,多糖物质主要存在于叶中,叶片越老多糖含量越高,在DNA提取过程中,这些物质容易与DNA不可逆地粘附在一起而污染DNA,从而抑制DNA限制性内切酶、T4 DNA连接酶、TaqDNA聚合酶等多种分了生物学酶类的活性,传统的CTAB法、SDS法等都很难达到要求,本实验中我们采用CTAB区室法提取米良一号猕猴桃叶片中的基因组DNA,并对其进行一定改进使所得到的基因组DNA能够满足PCR鉴定和Southern印迹杂交要求。
[1] 田宏现, 曾艳玲, 谭晓风, 等. 米良一号猕猴桃的组织培养研究[J] . 经济林研究, 2005, 23(1): 7-9.
[2] 姚泉洪, 黄晓敏, 刘宗镇. 植物乙烯生物合成与抗衰老基因工程[J].上海农业学报, 1994, 10(2): 89-96.
[3] 杨晓颖, 胡 伟, 徐碧玉, 等. 乙烯与果实成熟关系的研究进展[J].热带农业科学, 2008, 28(2): 70-75.
[4] 李日太, 张艳民. 中华猕猴桃果实成熟与其乙烯代谢的关系[J].淄博学院学报(自然科学与工程版), 2001, 7(1): 66-68.
[5] Lieberman M, Kunishi A, Mapson L, et al. Stimulation of Ethylene Production in Apple Tissue Slices by Methionine[J].Plant Phjsicl., 1966, 43: 376-382.
[6] 徐小彪, 张秋明. 基因工程与猕猴桃种质改良[J].果树学报,2003, 20(4): 300-304.
[7] 秦毓茜. 乙烯的生物合成及信号转导研究进展[J].安徽农业科学, 2006, 34(4): 622-624,640.
[8] 陈银华, 黄 伟, 王 海. ACC氧化酶基因研究进展[J].海南大学学报自然科学版, 2006, 24(2): 194-200.
[9] Hamilton A J, Lycett G W, Grierson D. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants[J].Nature, 1990, 346: 284-287.
[10] Picton S, Barton S L, Bouzayen M, et al. Altered fruit ripening and leaf senescence in tomato expressing an antisense ethyleneforming enzyme transgene[J]. Plant J., 1993, 3: 469-481.
[11] 任小林, 金志强, 彭世清,等. 猕猴桃ACC氧化酶cDNA克隆及全序列测定[J].园艺学报, 1997, 24(4): 333-337.
[12] MacDiarmid C W B, Gardner R C. A cDNA sequence from kiwifruit homologous to 1-aminocyclopropane- 1-carboxylic acid oxidase[J].Plant Physiol., 1993, 101: 691-692.
[13] 王曼玲, 朱虹琳, 周明全, 等. 莲藕组织总RNA的快速提取方法[J].武汉植物学研究, 2005, 23(5): 475-477.
[14] 谭晓风,胡芳名,谢禄山,等.油茶近成熟种子表达的发育相关基因及其分析[J].中南林业科技大学学报,2005,25(4):17-23
[15] Wall S J, Edwards D R. Quantitative reverse transcriptionpolymerase chain reaction (RT-PCR): A comparison of primerdropping, competitive, and real-time RT-PCRs[J].Analytical Biochemistry, 2002, 300(2): 269-273.
Genetic transformation of kiwifruit with its ACC oxidase antisense gene
TIAN Hong-xian1,2, YUAN Ping2, WANG Man-ling3, LIU Yi2, LI Jing2, XIA Xin-jie3, TAN Xiao-feng1
(1. Central South University of Forestry And Technology, Changsha 410004, Hunan, China; 2. Jishou University, Jishou 416000, Hunan,China; 3. Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, Hunan, China)
In order to establish the technology platform for the kiwifruit's with Its ACC oxidase antisense gene functions and improve kiwifruit quality through bio-technology, the kiwifruit leaves ( from Actinidia chinensis Planch.var.deliciosa‘Hayward’ ) were used to clone a cDNA of ACO gene, and its sequence was analysed.. The sequence was 1003 bp in length, with an open reading frame of 957 bp, encording 319 amino acid residues. The DNA sequence and its deduced amino acid sequence showed 94.0 % and 95.6 % homology and similarity with the sequences of Hayward plants in GenBank, respectively. Ten of the analyzed plants were shown to be positive for the ACO gene insert (71.4 %). The result of Quantitative Real-Time Fluorescence PCR Technology shows that the ACC oxidase gene expression of re-growth leaves was 58%~81% of the wild type.
Actinidia deliciosa (A. Chev.): ACC oxidase;agrobacterium-mediated;antisense expression vector
S759.3
A
1673-923X(2012)11-0115-07
2012-10-10
湖南省省级科技计划(专项计划)项目“耐贮保鲜高抗性转基因猕猴桃的研究和开发”(编号:2008JT3008);湖南省高校产学研合作示范基地开放项目(2011jsjk003)
田宏现(1953-),男,湖南保靖人,教授,硕士生导师,博士研究生,主要从事微生物学和林业生物技术方面的研究;E-mail:jstianhx@163.com;电话:13974310662
谭晓风(1956-),男,湖南茶陵人,教授,博士生导师,博士,主要从事经济林和林业生物技术研究;E-mail:tanxiaofengcn@126.com.cn;夏新界(1959-),男,博士,研究员,博士生导师,主要研究方向为作物耐逆分子生物学;E-mail:jxxia@isa.ac.cn
[本文编校:邱德勇]
猜你喜欢
小学生学习指导·低年级(2021年6期)2021-09-10
考试与评价·七年级版(2021年4期)2021-08-14
昆明医科大学学报(2020年12期)2021-01-26
水泵技术(2021年4期)2021-01-22
食品与生物技术学报(2021年4期)2021-01-17
石油化工技术与经济(2021年4期)2021-01-13
小学阅读指南·低年级版(2018年5期)2018-11-02
郑州大学学报(医学版)(2015年1期)2015-02-27
儿童故事画报·发现号趣味百科(2014年1期)2014-03-31
食品工业科技(2014年13期)2014-03-11
