
离子与通道相互作用对NaK通道通透特性影响的研究*
2011-10-25 07:09:04张素花安海龙刘玉芝张振东耿金鹏
物理学报 2011年4期
张素花 安海龙 刘玉芝 张振东 耿金鹏 展 永
1)(河北工业大学理学院,天津 300130)2)(天津市第三中学,天津 300131)(2010年3月24日收到;2010年8月5日收到修改稿)
离子与通道相互作用对NaK通道通透特性影响的研究*
张素花1)安海龙1)刘玉芝1)张振东2)耿金鹏1)展 永1)
1)(河北工业大学理学院,天津 300130)2)(天津市第三中学,天津 300131)(2010年3月24日收到;2010年8月5日收到修改稿)
以NaK通道的三维精细结构为基础,从理论上阐述了NaK通道对钠离子、钾离子、铷离子以及钙离子的通透机理,钡离子可以作为NaK通道阻断剂的微观机理,我们的研究结果表明,通道与离子的相互作用是决定通道对不同离子选择性的基础,反映通道与离子相互作用的位能曲线是通道对不同离子通透性的外在表现.
NaK通道,密度泛函,布朗动力学,通透性
PACS:87.10.- e,87.15.-v
1.引 言
离子通道是生物体中广泛存在的跨膜蛋白质分子.生命的很多过程如发育、生长、分泌、兴奋、运动,甚至于学习和记忆都与离子通道的正常发挥有直接关系[1—3].目前,有关离子通道的研究已经成为分子生物学、分子药理学、细胞生物学、生物化学和生物物理学研究热点.离子通道能够选择性的介导离子的跨膜转运,通道如何区分不同离子?通道对不同离子选择性的决定因素是什么?传统观点认为,钾离子通道的选择性过滤器的保守氨基酸序列是决定通道对不同离子产生选择性的原因,即通道结构决定通道对不同离子的选择性[4,5].文献[5]以及基于该文献我们开展的研究[4]都表明,决定通道离子选择性的不仅仅是选择性过滤器的氨基酸序列.对这些问题的研究,有助于人们在分子水平上理解通道蛋白的一些机理,帮助人们搞清膜的物质、信息输运机理,深入认识生命活动的本质,而且对单个离子通道工作机理的阐述可以帮助人们认识并最终治愈大量神经性和肌肉失调病症.
2006年Shi等人[6]报道了一种源自腊样芽孢杆菌的新型离子通道结构——NaK通道结构,它是第一个对K+没有高度选择性的阳离子通道结构.NaK通道和绝大部分的环核苷酸门控通道在选择过滤器结构、中心孔结构以及离子通透等方面有高度的相似性[7—9],但与钾离子通道相比,两者的结构高度相似,序列高度同源,值得注意的是NaK通道选择性过滤器氨基酸序列为TVGDG,而钾离子通道选择性过滤器氨基酸序列多是 TVGYG,KcsA钾离子通道选择过滤器中有4个等效的钾离子绑定位点,而NaK通道只保留了与钾离子通道等效的3,4位点,因此研究NaK通道的离子通透机理不仅可以帮助我们理解环核苷酸门控通道的离子通透机理,而且可以帮助我们进一步理解通道的离子选择性机理,以及通道的结构和功能的关系.
本文基于NaK通道的三维结构,从两个角度研究其通透性质:1)在充分考虑电子相互作用的基础上,采用密度泛函方法(DFT)在分子水平构造描述NaK通道与离子相互作用的位能曲线;2)建立描述离子跨膜输运的布朗动力学模型,得到了不同离子在通透过程中的位移时间关系曲线,从理论上解释离子通透的动力学特性.我们的研究结果表明,通道与离子的相互作用是决定通道对不同离子选择性的基础,而反映通道与离子相互作用的位能曲线可以直观的体现通道对不同离子的通透性.
2.位能曲线的计算
离子通道一般都是同源四聚体,4个亚基对称的围成一个传导离子的中央孔道[10,11].考虑通道结构的对称性,本文首先由文献[6]所给出的两个相邻亚基的三维精细结构,计算了完整通道四个亚基的过滤器结构,如图1所示.
通道对不同离子的选择作用主要发生在选择性过滤器部分,基因变异学研究发现,钾离子通道的选择性与位于选择性过滤器部分的氨基酸序列密切相关[11].因此本文以 NaK通道的选择性过滤器部分为研究对象,该系统包括20个氨基酸和1个阳离子,共1000余个电子,选择性过滤器细胞膜外侧、内侧原子坐标分别为 x=7×10-10m,x=28 ×10-10m.根据文献[12—14]给出的选择基组方案,结合我们所研究的系统,本文选择B3LYP算法和6-31G基组,各收敛参数均采用Gaussian 03程序包的缺省值.
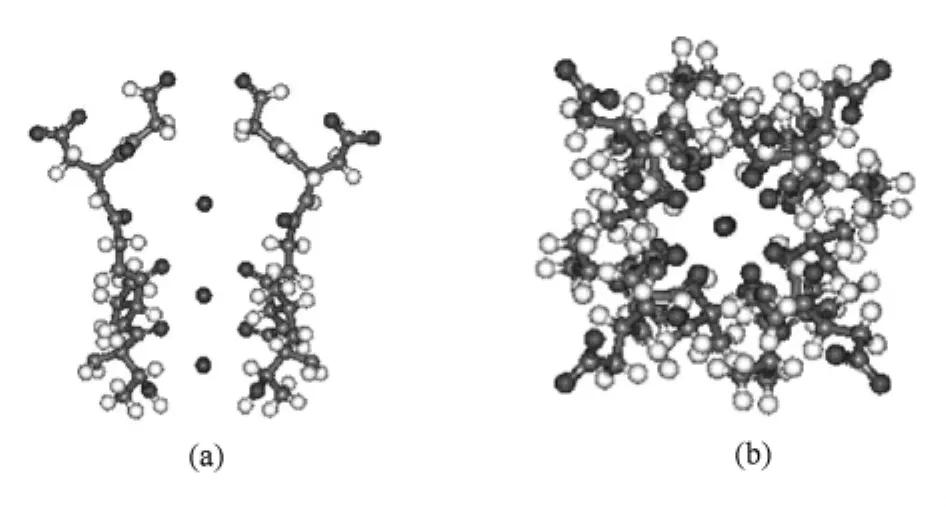
图1 NaK通道选择性过滤器的结构 (a)俯视图:为了便于观察只显示了两个亚基,图(a)中下方两个小球代表绑定位点3,4;(b)俯视图
从图2可以看出,钠、钾、铷、钙离子位能曲线在(18—22)×10-10m范围内都存在极小值,这与实验中测得的结合位点3,4对应;并且钠、钾、铷、钙离子位能曲线具有非对称性,细胞膜内侧高于细胞膜外侧,根据统计热力学的观点,非对称的位能曲线能够驱动钠、钾、铷、钙离子通过 NaK通道产生由膜内侧到膜外侧定向移动,完成离子的跨膜输运过程.
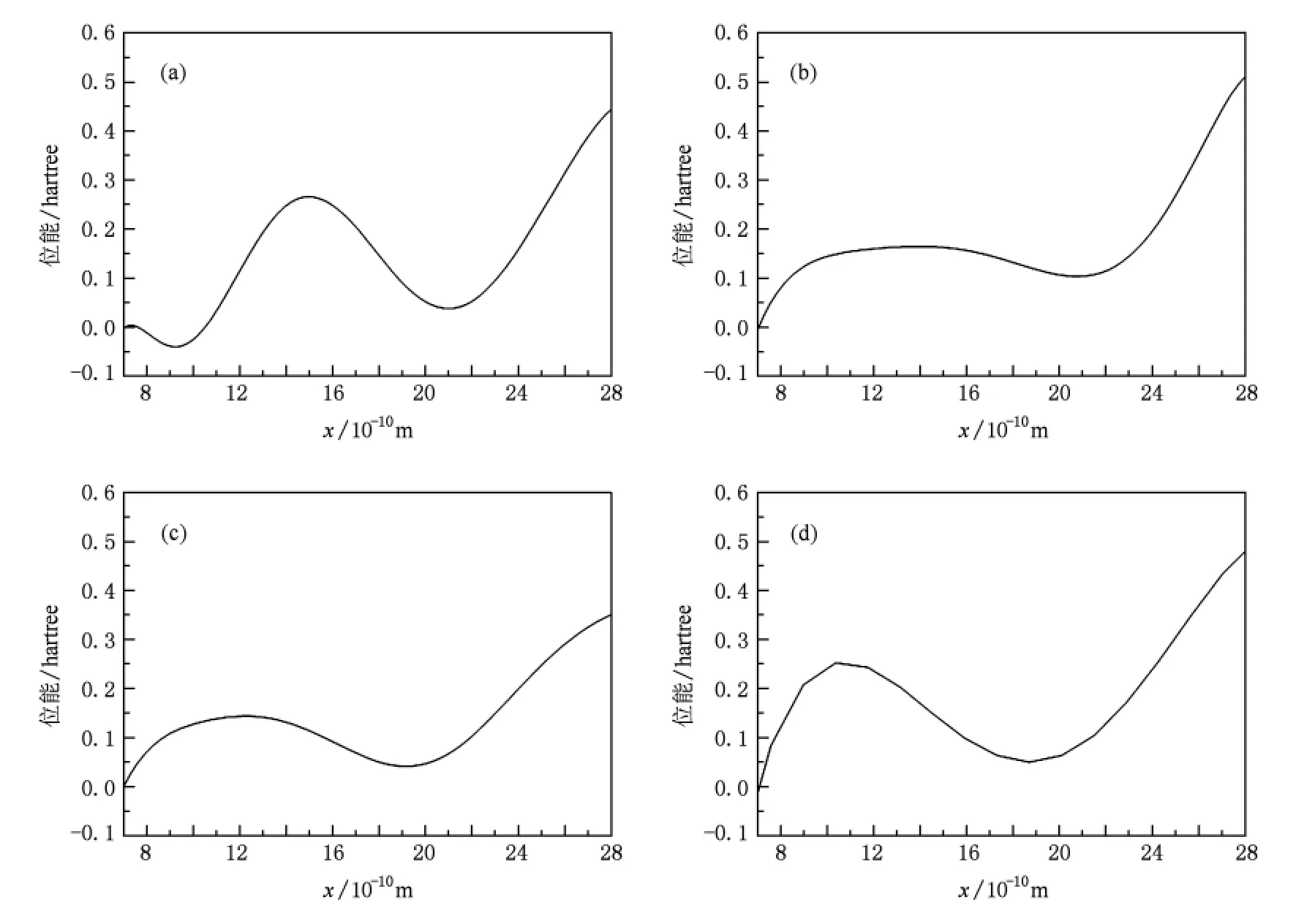
图2 钠、钾、铷、钙离子在NaK通道中的位能曲线(右边对应细胞膜内侧),(a),(b),(c),(d)分别对应钠、钾、铷、钙离子与通道相互作用的位能曲线(1 hartree=110.5×10-21J)

图3 钡离子在NaK通道中的位能曲线(右边对应细胞膜内侧)
图3表明,钡离子位能曲线具有对称性,在(18—22)×10-10m范围内存在极小值,说明钡离子可以绑定在该位置,这与电子密度图实验结构一致.由非平衡统计物理学观点:位于对称势阱内的粒子不能产生定向移动.由于钡离子可以绑定在通道内,占据结合位点,导致其他离子不能进入通道产生定向移动.
以上计算、分析的结果与实验结果[6,8]相符合,说明通道和离子间的相互作用决定了通道对离子的选择通透性.
3.布朗动力学方法
布朗动力学是统计力学中用来研究非平衡系统动力学行为的常用方法.离子通道的布朗动力学模型把离子在通道内的行为看作是随机动力学行为,由通道与离子的相互作用及膜电位提供驱动力,把通道内的离子看作布朗粒子,考虑离子的运动是一维运动,Langevin方程可以简化为
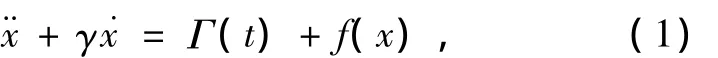
其中γ是单位质量的阻尼系数.Γ(t)是一种涨落力,来源于细胞内水分子及其他离子对离子的作用力.其统计平均值为0,关联函数具有δ函数的形式
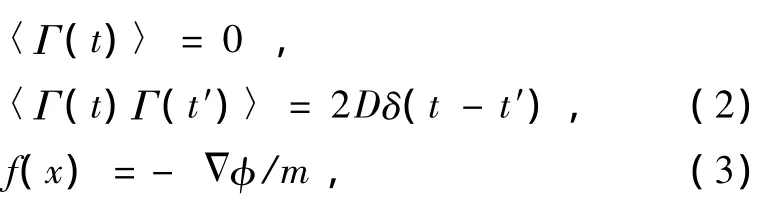
为单位质量的布朗粒子受到的作用力,该力起源于通道对离子的作用,其大小等于位能曲线负梯度.(3)式中 为离子在通道中的位能.
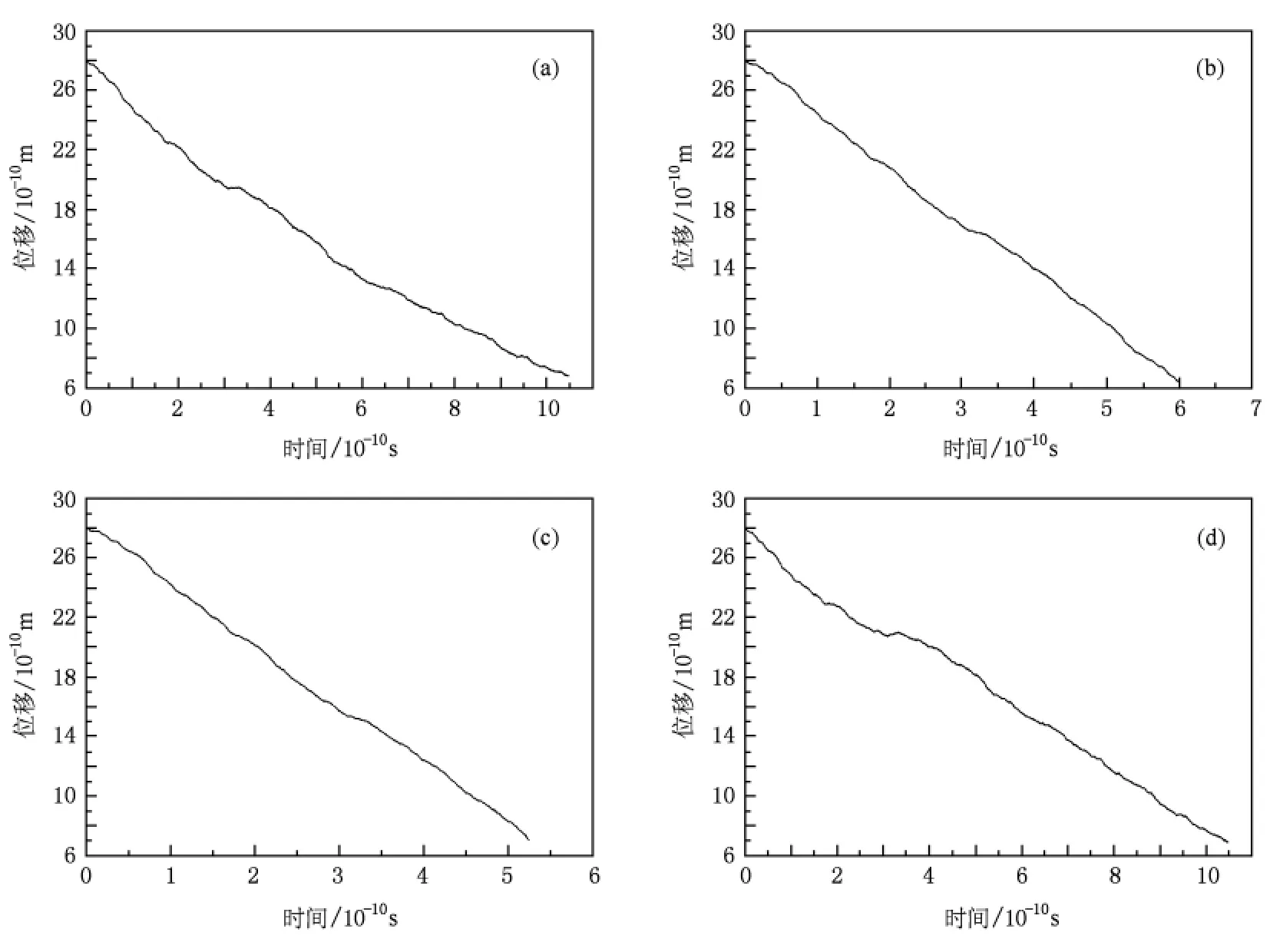
图4 钠、钾、铷、钙离子平均位移随时间变化曲线,(a),(b),(c),(d)分别对应钠、钾、铷、钙离子与通道相互作用的位能曲线
给定初始位移 x0、初始速度 v0,采用随机龙格-库塔算法[15,16]求解郎之万方程.由于随机力的作用,在相同的初始条件下,离子的轨道具有随机性.为了使模拟结果更精确,模拟过程中对同一离子模拟10000条轨道,通过对这些轨道的平均得到离子的平均位移随时间的变化曲线,如图4所示.模拟的轨道数越多,模拟的结果就越精确.由位移时间关系曲线,可以得到平均速度、平均通透时间以及单位时间内的通透离子个数——平均通透频率等动力学参数.
图4表明:随着时间的推移,钠、钾、铷、钙离子从中心腔内经过选择性过滤器到达膜外侧,但由于通道对不同离子的作用能不同,以致钠、钾、铷、钙离子通过通道的时间不同,铷离子的通透时间最短,而钙离子的通透时间最长(如表1所示).
结合图4和表1,我们可以得出以下结论:1)钠离子、钾离子、铷离子及钙离子都可以通过NaK通道到达膜外侧,即NaK通道对阳离子没有选择性.2)不同离子在通过通道的过程中,所遇到的位垒高度不尽相同,由此导致通透动力学过程的多样性,钾、铷离子具有相对较小的平均通透时间和较大的通透频率,即NaK通道对钾、铷离子更具通透性.3)计算得到的通透频率在数量级上与文献[17]提到的单通道每秒钟可以通过约106—108个离子一致,可见NaK通道的布朗动力学模拟的可靠性.
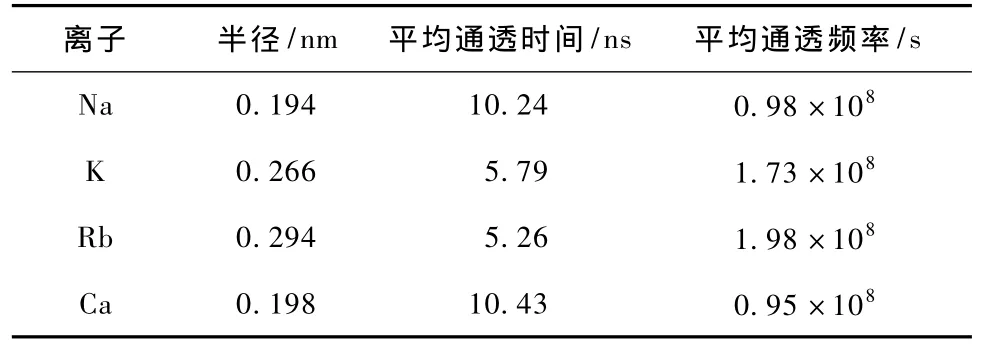
表1 钠、钾、铷、钙离子直径及其动力学参数
4.结 论
本文研究了NaK通道对钠、钾、铷、钙离子以及钡离子的通透特性,得到了一些有意义的结论,对进一步理解通道结构和功能的关系做了有益的探索:NaK通道与钠离子、钾离子、铷离子和钙离子相互作用位能曲线具有非对称性,钠离子、钾离子、铷离子和钙离子可以在通道内通透,NaK通道对这些离子没有选择性;钡离子与NaK通道的位能曲线具有对称性,钡离子被绑定在通道内部,导致其他离子不能通透,这是钡离子可以作为该通道阻断剂的微观机理;布朗动力学的计算结果与文献结果一致.
附 录 密度泛函方法
密度泛函理论是基于Hohenberg2Kohn第一定理和第二定理,通过确定体系的基态电子密度,同时确定体系的电子数,从而决定了体系的哈密顿算符.体系的能量是电子密度的泛函,根据泛函的变分原理,即体系基态总能量(表示成粒子密度的泛函形式)在体系基态单粒子密度处取极小值,即为体系的基态真实总能量.离子处于不同位置体系的电子分布也就不同,这样就可以通过计算离子在不同位置的电子密度,得到体系的位能面.在密度泛函中,钾离子通道的单点能具有如下形式:

式中各项均是电子密度的函数,其中ET是N个电子所具有的动能,EV是电子势能项,EJ是第i个电子与N-1个电子的排斥能,EXC是交换相关项(包括交换能和相关能).N为体系电子数目.
与其他理论方法比较,DFT具有如下优势:
1)DFT采用泛函方法求解薛定谔方程,既可以考虑原子与原子间的相互作用,又可以考虑电子相关,这有别于在经典物理基础上发展起来的分子理论方法,后者只能考虑化学键和原子间的相互作用,而忽略了电子的作用.
2)DFT采用泛函方法求解薛定谔方程,不同于量子化学计算中的半经验方法,后者需要输入一些实验或者经验参数,帮助求解薛定谔方程.
3)密度泛函方法的计算量仅约正比于N3(N为体系的电子数),而不是从头算的 N4和 CI与 MP的 N6—N8.应用密度泛函理论方法的计算量对于体系电子数变化不是特别敏感,该方法比较适用于研究计算生物大分子的性质.
[1] Tang Y Q 1998 Chemistry of Life(Hunan Science& Technology Press)P96(in Chinese)[唐有祺1998生命的化学(湖南科学技术出版社)第96页]
[2] Wang H Q,Yu LC,Chen Y 2009 Acta Phys.Sin.58 5070(in Chinese)[王慧巧、俞连春、陈 勇2009物理学报58 5070]
[3] Qiao X Y,Li G,Dong Y E,He B J 2008 Acta Phys.Sin.57 1259(in Chinese)[乔晓艳、李 刚、董有尔、贺秉军2008物理学报 57 1259]
[4] Heginbotham L, LuZ, AbramsonT, MacKinnonR 1994 Biophys.J.66 1061
[5] So I,Ashmole I,Davies N W,Sutcliffe M J,Stanfield P R 2001 J.Physiol.53 37
[6] Shi N,Ye S,Alam A,Chen L P,Jiang Y X 2006 Nature 440 570
[7] Zagotta W N 2006 Nature 440 427
[8] Alam A,Shi N,Jiang Y X 2007 Proc.Natl.Acad.Sci.U.S.A.104 15334
[9] Kaupp U B,Seifert R 2002 Physiol.Rev.82 769
[10] Jiang Y,Pico A,Cadene M,Chait B T,MacKinnon R 2001 Neuron 29 593
[11] Mashl R J,Tang Y,Schnitzer J,Jakobsson E 2001 Biophys J.81 2473
[12] Zhang R Q,Bu Y X,Li Sh T 2000 Chinese Science B 30 419(in Chinese)[张瑞勤、步宇翔、李述汤 2000中国科学 B 30 419]
[13] Chen L,Xu C,Zhang X F 2009 Acta Phys.Sin.58 1603(in Chinese)[陈 亮、徐 灿、张小芳 2009物理学报 58 1603]
[14] Fang F,Jiang G,Wang H Y 2006 Acta Phys.Sin.55 2241(in Chinese)[方 芳、蒋 刚、王红艳 2006物理学报 55 2241]
[15] Rebecca L 1992 Phy.Rev.A 15 600
[16] Zhan Y,Zhao T J,Yu H,Song Y L 2002 Chin.Phys.11 624
[17] Hille B 2001 Ion Channels of Excitable Membranes(Sinauer Associates,INC.Publishers 3rd edition)p72
Effect of ion-channel interaction permeability of NaK channel*
Zhang Su-Hua1)An Hai-Long1)Liu Yu-Zhi1)Zhang Zhen-Dong2)Geng Jin-Peng1)Zhan Yong1)
1)(Hebei University of Technology,Tianjin 300130,China)2)(The Third Middle School of Tianjin,Tianjin 300131,China)(Received 24 March 2010;revised manuscript received 5 August 2010)
Based on the three-dimensional structure of NaK channel,the permeabilities of the NaK channel for Na+,K+,Rb+,Ca2+ions and the Ba2+ion blockage mechanisms are investigated.The results show that the interaction is fundamental for determining the selectivities of the channels and the potential curve reflects the external behaviors for the permeabilities of different ions.
The NaK channel,density functional theory,Brownian dynamic,permeability
.E-mail:zhany@hebut.edu.cn
*国家自然科学基金(批准号:10775038,10975045),教育部博士学科点专项基金(批准号:20060080001),河北省自然科学基金(批准号:C2007000026,C2009000029,C2009001104)资助的课题.
E-mail:zhany@hebut.edu.cn
*Project supported by the National Natural Science Foundation of China(Grant Nos.10775038,10975045),the Specialized Research Fund for the Doctoral Program of Higher Education of China(Grant No.20060080001),The Natural Science Foundation of Hebei Province,China(Grant Nos.C2007000026,C2009000029,C2009001104).
PACS:87.10.- e,87.15.-v
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25 13:17:14
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:31:36
第一财经(2019年8期)2019-08-26 17:53:46
电子测试(2018年9期)2018-06-26 06:45:56
趣味(语文)(2018年2期)2018-05-26 09:17:55
哈尔滨医药(2015年2期)2015-12-01 03:57:13
学习月刊(2015年14期)2015-07-09 03:37:48
物理化学学报(2015年5期)2015-02-28 17:34:47
自动化博览(2014年6期)2014-02-28 22:32:20
中国氯碱(2014年11期)2014-02-28 01:05:07
