
几种激发子处理对苹果梨采后青霉病的控制
2011-10-24 08:24:52杨志敏李永才陈松江范存斐王云飞张祖斌
食品工业科技 2011年7期
杨志敏,毕 阳,李永才,陈松江,尹 燕,范存斐,王云飞,张祖斌
(甘肃农业大学食品科学与工程学院,甘肃兰州 730070)
几种激发子处理对苹果梨采后青霉病的控制
杨志敏,毕 阳,李永才*,陈松江,尹 燕,范存斐,王云飞,张祖斌
(甘肃农业大学食品科学与工程学院,甘肃兰州 730070)
研究了β-氨基丁酸(BABA)、水杨酸(SA)、Na2SiO3和NO供体硝普钠(SNP)处理对苹果梨抗青霉病诱导的时效性及其作用机理。结果表明,各激发子处理后不同时间损伤接种均能不同程度地抑制扩展青霉(Penicillium expansum)的侵染和扩展,处理后18h接种各处理的病斑直径明显低于对照,其中BABA处理效果最为明显,病斑直径比对照降低了14.59%,SNP次之。进一步研究表明,各激发子处理对果实组织抗性相关酶过氧化物酶(POD)、多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)均有一定的影响,其中SNP能明显提高POD活性,PAL活性能显著地被SNP和BABA诱导,而对于PPO活性,SNP和Na2SiO3处理的诱导作用高于BABA和SA。
苹果梨,激发子,扩展青霉,抗性相关酶
苹果梨(Pyrus bretchneideri cv.Pingguoli)是甘肃省名优特产,颇具市场声誉。其虽具良好的耐贮性,但贮藏后期腐烂仍很严重,其中扩展青霉(Penicillium expansum)引起的青霉病是其主要的采后病害[1]。采用化学杀菌剂可有效控制青霉病,但由于杀菌剂残留、环境污染及产生抗药性等问题使其应用受到了限制。因此,寻找新的、更安全有效的果蔬防腐措施已成为当前生产中亟待解决的问题[2]。近年来,用各种诱导剂处理来提高产品抗病性已成为果蔬采后防腐的研究热点。β-氨基丁酸(BABA)作为一种对环境安全且具高效诱抗作用的非蛋白氨基酸类诱抗剂,能诱导番茄等果蔬对卵菌或真菌病害的系统获得抗病性[3]。水杨酸(SA)被认为是诱发植物系统获得抗病性(SAR)的信号物质,能诱导黄瓜[4]、柑 桔 和 马 铃 薯[5]、香 蕉[6]、猕 猴 桃[7]和 苹 果梨[8]等果蔬对采后病害产生抗性。硅作为地壳中存在的第二大类元素,研究表明可用于植物病害的控制[9],其中作为一般认为安全物质(GRAS)的Na2SiO3采后处理可明显抑制哈密瓜黑斑病、白霉病和粉霉病[10]、苹果梨黑斑病[11]以及马铃薯块茎干腐病[12]、苹果梨青霉病[13]等。NO 是一类脂溶性可扩散小分子物质,作为信息传递的信使分子,不仅参与植物生长和发育等生理代谢过程[14],而且对植物的抗逆性具有一定的作用[15],它是植物和病原物互作过程中的一种信号分子[16,18]。本研究拟采用SNP、Na2SiO3、BABA和 SA等几种激发子,研究其对苹果梨果实抗青霉病的诱导以及对抗病相关酶活性的影响,以期为采后苹果梨青霉病的控制筛选低毒、安全的防腐剂。
1 材料与方法
1.1 实验材料
供试苹果梨(Pyrus bretchneideri cv.Pingguoli)采自甘肃景泰县条山农场,单果包纸,纸箱包装后运抵实验室冷藏待用;供试扩展青霉(Penicillium expansum) 参照方中达[19]的方法分离自自然发病苹果梨果实,纯化鉴定后PDA上保存待用;硝普钠(SNP)、水杨酸(SA) 天津市光复精细化工研究所;硅酸钠(Na2SiO3) 天津市科密欧化学试剂开发中心;β-氨基丁酸(BABA) sigma公司。
1.2 实验方法
1.2.1 激发子处理对苹果梨损伤接种病斑直径的影响 参考刘志等[20]的方法并改进,选择外观整齐、无病虫害的苹果梨洗净后晾干,75%酒精表面消毒后,用灭菌打孔器在苹果梨表面等距离打孔4个,深度为 3mm。1h后将浓度 为 100μmol/L的 SNP、100mmol/L的 Na2SiO3、500μmol/L的 SA 和50mmol/L的BABA各20μL注入孔内,取另一组作为对照,加入等量的无菌水。分别于6、12、18、24h后用微量加样器接入1×106个/m L的孢子悬浮液20μL,晾干后用聚乙烯袋包裹,然后在室温下贮藏并观察发病情况,采用十字交叉法测量病斑直径并记录结果。每处理用果实10个,重复三次。
1.2.2 酶液提取及酶活性测定
1.2.2.1 粗酶液的制备 取经不同激发子处理后0、6、12、18、24、48h 的果实,在果皮面下 1mm 处取果肉组织样品3g,用锡箔纸分包,包好后立即用液氮冷冻,并保存在-85℃超低温冰箱中备用,每个处理用果10个。参照曹建康等[21]的方法并做修改:取保存待测的果肉组织3g于预冷的研钵中,加入4m L提取缓冲液,在冰浴条件下充分研磨成匀浆,在4℃,12000×g条件下离心15m in,上清液为粗酶液。
1.2.2.2 酶活的测定 POD和PPO活性测定均参照曹建康等[21]的方法,分别以每分钟反应体系在波长470nm和420nm处吸光度值读数变化增加1所需的酶量为一个酶活单位,单位是(ΔOD470/m in·mg蛋白质和ΔOD420/m in·mg蛋白质)。PAL活性测定参照曹建康等[21]的方法并修改,以每小时反应体系在波长290nm处吸光度值读数变化增加1所需的酶量为一个酶活单位,单位是(ΔOD290/h·mg蛋白质)。
1.2.2.3 蛋白含量及标准曲线的测定 蛋白质含量测定参照曹建康等[21]的方法,同时标准曲线以牛血清蛋白(BSA)为标准蛋白,计算蛋白含量。
1.3 结果统计
数据处理采用Microsoft Excel 2003软件进行分析,并计算标准误差(±SE)。采用 SPSS Statistics 16.0软件进行显著性差异分析。
2 结果与分析
2.1 激发子处理对苹果梨抗青霉病的诱导效果
各激发子处理后不同时间接种青霉病菌的果实,24h内均能不同程度地防止青霉病菌的侵染和扩展(图1)。处理后12h内损伤接种4种激发子的诱导效果均不明显,随着时间延长,病斑直径除BABA处理外,其它均呈先降低后趋于平缓的趋势,其中18h的诱导效果较好,而BABA处理24h接种效果最好,比对照降低了13.8%。

图1 激发子处理对苹果梨抗青霉病的时效性
从图2可知,处理后18h接种各激发子对青霉病菌的扩展均存在不同程度的抑制作用。SA、Na2SiO3、SNP和BABA处理的病斑直径分别比对照降低5.99%、6.15%、7.10%、14.59%,其中BABA处理的诱导效果显著高于对照和其它激发子处理。
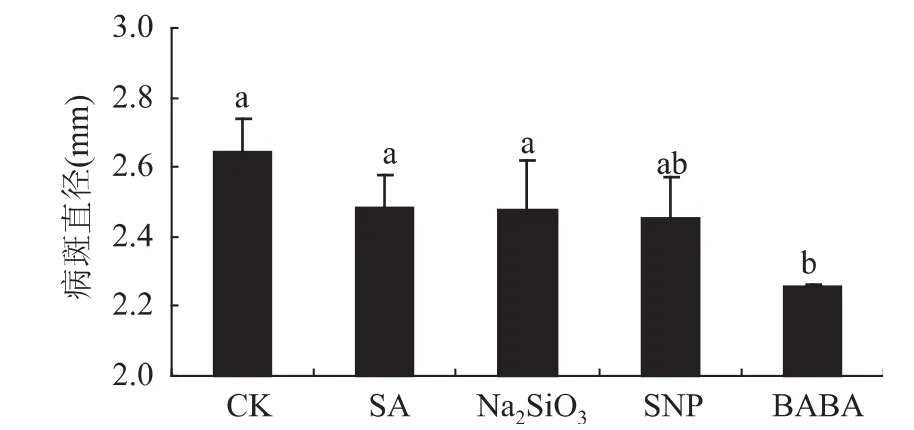
图2 激发子处理后18h接种对苹果梨青霉病的控制效果
2.2 激发子处理对苹果梨组织抗性相关酶活性的影响
2.2.1 激发子处理对苹果梨POD活性的影响 各激发子处理对苹果梨组织POD活性均有一定的影响(图3)。处理后18h之前除SNP处理的POD活性高于对照外,其余各处理均与对照无明显差异。在处理后18h各处理的POD酶活性均下降,达到最小值。随后各处理POD活性均呈先上升后下降的趋势。其中Na2SiO3和SNP处理明显增强了果实组织的POD活性,处理后24h高出对照21.95%和26.03%。都呈单峰曲线。在处理后12h到18h PPO酶活性急剧上升,18h时各处理酶活性均达到最大值,其中Na2SiO3处理果实的PPO活性在18h明显高于对照及其它处理,其酶活性比对照高20.47%。随后各处理的酶活性均呈现下降趋势,其中SNP处理的酶活性下降缓慢,在处理后24h和48h较其它处理均保持较高的酶活性,其酶活性值分别较对照高出29.42%和14.56%。

图3 不同激发子处理对苹果梨POD酶活性的影响
2.2.2 激发子处理对苹果梨PPO活性的影响 从图4可以看出,不同激发子处理的果实PPO活性基本

图4 不同激发子处理对苹果梨PPO酶活性的影响
2.2.3 激发子处理对苹果梨PAL活性的影响 SA、Na2SiO3、SNP和BABA 4种激发子处理虽对苹果梨果实组织PAL活性均有一定的影响,但各处理对酶活性影响的变化趋势各不相同(图5)。处理后48h内,除SA和BABA平稳波动上升外,Na2SiO3与对照趋势相似且低于对照,24h时SNP处理的峰值最高,高出对照44.86%,SNP和BABA在48h酶活均显著高于对照,约高出对照1.19倍。
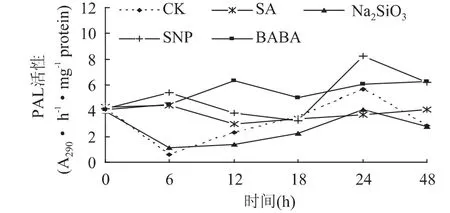
图5 不同激发子处理对苹果梨PAL酶活性的影响
3 讨论
随着诱导抗性在果实采后病害控制中应用的增多,诱导因子的筛选及其作用机理的研究越来越被广泛关注。许多研究表明,SA[22]和 NO[17-18]处理能够激活植物过敏性反应和获得性系统抗性的提高,是重要的信号分子。本实验中用SA和SNP处理苹果梨果实,降低了扩展青霉对果实的危害程度,说明SA和SNP能诱导苹果梨产生抗病性,激活组织自身的防御反应,这与刘志[20]、王芳芳[23]在富士苹果及 Tian等[24]在梨上的结果相似。BABA作为非蛋白类氨基酸,能诱导植株细胞壁结构发生变化,产生胼胝质,形成乳突或导致细胞木质化等,具有防止侵染和增强抗病性作用的结构[25]。本实验中BABA处理能有效控制苹果梨青霉病是对上述结论的有力证明。同时发现Na2SiO3处理能够抑制苹果梨的青霉病,该结果与前人在苹果梨[10]、马铃薯[11]和甜瓜[26]上的研究结果相一致,表明Na2SiO3处理能有效地控制果蔬采后病害的发生。
POD是寄主植物细胞内抵御活性氧伤害的主要保护酶,在阻止活性氧形成和清除活性氧中起重要作用,与植物的抗病反应密切相关。一般认为,POD活性的提高是果实抗病性被诱导的重要生化指标[12]。PPO是酚类物质氧化的主要酶,在植物体内催化酚类物质氧化产生醌类,并参与木质素合成,以杀死或抑制病原菌的繁殖而起到抗病作用[27]。本实验中Na2SiO3和SNP处理明显促进了果实体内POD和PPO活性增加,表明其诱导了苹果梨的抗病性,这与 Na2SiO3处理马铃薯[11]、哈密瓜[12]以及 SNP 处理富士苹果[20]结果相一致。PAL则是苯丙烷类代谢途径中的第一个关键酶,与植保素、木质素及酚类化合物的形成密切相关,在PAL的作用下,苯丙氨酸脱氨基可以生成酚类和其他具有抗菌活性的物质,它们对抑制病原菌具有明显的作用[28]。在实验中,苹果梨果实经SA、Na2SiO3、SNP和BABA处理后,均可不同程度地提高PPO、POD、PAL三种酶的活性水平,但各个处理对不同酶的活性影响的变化模式和程度有所不同。综上所述,四种激发子虽然能诱导苹果梨使其产生抗病反应,对苹果梨青霉病进行控制,但其诱导机理可能存在差异,尚有待进一步的研究。
[1]张维一,毕阳.果蔬采后病害与控制[M].北京:中国农业出版社,1996.
[2]田世平,范青.控制果蔬采后病害的生物技术[J].植物学通讯,2000(17):211-217.
[3]Cohen Y R.β-aminobutyric acid induced resistance against plant pathogens[J].Plant Disease,2002,86(5):448-457.
[4]李淑菊,马德华,庞金安,等.水杨酸对黄瓜几种酶活性及抗病性的诱导作用[J].华北农学报,2000,15(2):118-122.
[5]Gaur A,Chenulu W.Chemical control of postharvest diseases of Citrus reticulata and Solanum tuberosum[J].Indian Phytopathology,1982,35(4):628-632.
[6]Ram V,Vir D.Efficacy of fungicide XXXVI.Relative evaluation of various postharvest chemical treatments against spoilage of banana fruits caused by Culvularia lunata[J].India Phytopathology,1986,39(4):594-595.
[7]Poole PR,Mcleod L C.Development of resistance to picking wound entry Botrytis cinerea storage rots in kiwifruit[J].New Zealand Journal of Crop and Horticultural Science,1994,22(4):387-392.
[8]曹建康,毕阳,李永才,等.水杨酸处理对苹果梨采后黑斑病及贮藏品质的影响[J].甘肃农业大学学报,2001,36(4):438-442.
[9]Bi Y,LiY C,Ge Y H.Induced resistance in postharvest fruits and vegetables by chemicals and its mechanism[J].Stewart Postharvest Review,2007,3(6):1-7.
[10]郭玉蓉,葛永红,毕阳,等.采后硅酸钠处理对苹果梨黑斑病的影响[J].食品科学,2003,24(12):140-142.
[11]盛占武,毕阳,鄯晋晓,等.采后硅酸钠处理对马铃薯干腐病的抑制[J].食品工业科技,2007(9):190-191.
[12]Bi Y.Tian S P,Guo Y R,et al.Sodium silicate reduces postharvest decay on Hami melons induced resistance and fungistatic effects[J].Plant Disease,2006,90(3):279-283.
[13]李云华,毕阳,张怀予,等.采后硅酸钠处理对苹果梨黑斑病的抑制[J].甘肃农业大学学报,2008,43(6):150-153.
[14]Leshem Y Y,Haramaty E.The characterization and contrasting effects of the nitric oxide free radical in vegetative stress and senescence of Pisum sativum Linn.foliage[J].Plant Physiology,1996,148:258-263.
[15]Delledonne M,Xia Y J,Dixon R A,et al.Nitric oxide function as a secondary signal in plant disease resistance[J].Nature,1998,394:585-588.
[16]Romero-Puertas mC,Rodrigues-Serrano M,Corpas F J,et al.Cadmium-induced subcellular accumulation of·and H2O2in pea leaves[J].Plant Cell and Environment,2004,27(9):1122-1134.
[17]Wendehenne D,Durner J,Klessig D F.Nitric oxide:a new player in plant signaling and defence responses[J].Current Opinion in Plant Biology,2004,7(4):449-455.
[18]Delledonne M.NO news is good news for plants[J].Current Opinion in Plant Biology,2005,8(4):390-396.
[19]方中达.植病研究方法[M].第三版.北京:中国农业出版社,1998.
[20]刘志,张华磊,谢兴斌,等.激发子物质处理富士苹果果实后抗轮纹病病菌侵染的研究[J].果树学报,2009,26(5):666-670.
[21]曹建康,姜微波,赵玉梅.果蔬采后生理生化实验指导[M].北京:中国轻工业出版社,2007.
[22]Sticher L,Mauch-Mani B,Métraux JP.Systemic acquired resistance[J].Annual Review of Phytopathology,1997,35:235-270.
[23]王芳芳.外源水杨酸诱导苹果抗轮纹病效应的研究[D].河北农业大学,2008:14-24.
[24]Tian S P,Wan Y K,Qin gZ,et al.Induction of defense responses against Alternaria rot by different elicitors in harvested pear fruit[J].Applied Microbiology and Biotechnology,2006,70(6):729-734.
[25]Cohen Y,Reuveni M,Baider A.Local and systemic activity of BABA(DL-β-aminobutyric acid)against Plasmopara viticolain grapevines[J].European Journal of Plant Pathology,1999,105:351-361.
[26]郭玉蓉,陈德蓉,毕阳,等.硅化物处理对甜瓜白粉病的抑制效果[J].果树学报,2005,22(1):35-39.
[27]郭红莲,程根武,陈捷,等.玉米灰斑病抗性反应中酚类物质代谢作用的研究[J].植物病理学报,2003,33(4):342-346.
[28]Nicholson R L,Hammerschmidt R.Phenolic compounds and their role in disease resistance[J].Annual Review of Phytopathology,1992,30:369-389.
Inhibiting effect of several elicitors treatment on blue mould of Pingguolipear
YANG Zhi-min,BI Yang,LI Yong-cai*,CHEN Song-jiang,YIN Yan,FAN Cun-fei,WANG Yun-fei,ZHANG Zu-bin
(College of Food Science and Engineering,Gansu Agricultural University,Lanzhou 730070,China)
Induction disease resistance against bluemould in Pingguoli pear by different elicitors,such as β-aminobutyric acid(BABA),salicylic acid(SA),sodium silicate and nitric oxide(NO)donor sodium nitroprusside(SNP)treatment at different time intervals after treatment and their possible mechanism s were studied.The results showed that there was a different reduction level in lesion diameter of Pingguolipear treated with four elicitors at different time intervals after treatment.BABA showed the best result in reducing lesion diameter by 14.59%com pared with the control,and then the inhibiting effect of SNP was better than others at 18h after treatments.Further studies showed that peroxidase(POD),polyphenol oxidase(PPO)and phenylalanine ammonialyase(PAL)of the fruit tissue were affected by elicitors.SNP could increase the activity of POD enzyme,PAL activity could be significantly induced by SNP and BABA,SNP and Na2SiO3were better than BABA and SA treatments in induction of PPO activity increase.
Pingguolipear;elicitor;Penicillium expansum;resistant related enzyme
TS255.3
A
1002-0306(2011)07-0379-04
2010-07-06 *通讯联系人
杨志敏(1985-),女,硕士研究生,主要从事采后生物学方面的研究。
国家自然科学基金(30960243)。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:28
内蒙古林业(2021年3期)2021-03-23 06:22:18
农产品市场周刊(2019年12期)2019-08-30 08:22:30
农产品市场周刊(2019年9期)2019-07-02 16:42:14
少年文艺·我爱写作文(2016年10期)2016-09-10 07:22:44
中国当代医药(2015年26期)2015-03-01 02:06:51
现代检验医学杂志(2015年4期)2015-02-06 02:02:12
现代检验医学杂志(2015年2期)2015-02-06 02:00:51
食品工业科技(2014年9期)2014-03-11 18:15:31
食品科学(2013年22期)2013-03-11 18:29:40
