
LE540对视网膜色素上皮bFGF及TGF-β2的影响
2011-10-20 06:14:34苗英彬张未娟邓志宏曾丽明杨瑞波赵少贞
天津医科大学学报 2011年2期
苗英彬,张未娟,邓志宏,曾丽明,杨瑞波,张 琛,赵少贞
(天津医科大学眼科中心屈光角膜科,天津300384)
近年来,视网膜色素上皮(retinalpigmentepithelium,RPE)细胞在近视发生中的作用已成为眼科研究中的热点。体外研究表明视黄酸可以调节RPE细胞分泌碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)、 转 化 生 长 因 子 β2(transfergrowth factor-β2,TGF-β2)等生长因子,而这些细胞因子都和近视的发生发展有着密切联系[1]。但RPE细胞内通过何种信号转导途径调节细胞因子的分泌尚未明确。本研究通过在培养的豚鼠RPE细胞加入全反式视黄酸和视黄酸β受体阻断剂LE540来观察其对细胞分泌bFGF和TGF-β2的影响,寻找控制人类近视发展的药物靶点。
1 材料与方法
1.1 主要仪器及试剂 CO2培养箱 (美国Forma公司);倒置生物显微镜(日本Olympus公司);培养板、培养瓶(美国Corning公司);DMEM/F12培养基、胎牛血清(fetal bovine serum,FBS)、胰蛋白酶(美国Gibco公司);小鼠抗大鼠广谱细胞角蛋白(pan cytokeratin,PCK)单克隆抗体(1∶200)(武汉博士德公司);LE540(日本 Wako);bFGF、TGF-β2ELISA 试剂盒(美国ADL公司)。
1.2 豚鼠RPE细胞的培养及鉴定 2~3周龄健康普通级三色豚鼠4只,雌雄不限(购自北京维通利华实验动物技术有限公司),腹腔注射过量水合氯醛处死,摘除眼球,去除眼周组织,PBS冲洗,于含0.2 g/L庆大霉素的PBS中4℃浸泡20~30min后取出眼球用PBS冲洗2~3次,于装满PBS的培养皿中沿锯齿缘剪开眼球去除眼前节部分,去除玻璃体和神经视网膜层,置于干净平皿中撕下色素膜浸于0.25%的胰蛋白酶中37℃消化10~15min。加入培养液吹打,1 000 r/min离心5min,弃上清,加入培养液吹打,200目细胞筛过滤,收集于培养瓶中培养。原代培养用含20%FBS的DMEM/F12培养基,传代后用含10%FBS的培养基培养。2~3 d后第1次换液,洗掉未贴壁的细胞。5~7 d后细胞长满传代。先加入少量0.25%的胰蛋白酶消化30 s,用PBS洗涤,弃去洗液以去除贴壁不牢的其他细胞。再加入胰蛋白酶于37℃孵箱中消化1~2min,震荡后可见细胞脱落时加入含10%FBS的培养液终止消化,吹打后移入离心管,1 000 r/min离心5min,按1∶3传代。培养30~60min,稍震荡后转移细胞悬液至另一培养瓶以去除贴壁较快的成纤维细胞。取第2代细胞,进行上皮细胞特异的角蛋白免疫组织化学染色,采用小鼠抗大鼠PCK单克隆抗体一抗(1∶200)鉴定后用于实验。
1.3 ELISA检测bFGF、TGF-β2的分泌 第2代细胞接种于24孔板,待细胞长满后换无血清培养液,24 h后分别换含10×10-6mol/LATRA和5×10-6mol/L LE540 的无血清培养液,分别于换液后 2、4、6、8、16 h收集培养液,-20℃保存。按试剂盒说明步骤严格操作检测 bFGF、TGF-β2的分泌量:(1)取出酶标板分别加入50μL标准品或样品于空白孔。(2)在样品孔中加入10μL生物素标记液。(3)在标准品和样品孔中加入100μL酶标记液。(4)37℃孵育60min。(5)清洗5次。(6)每孔加入底物A液、B液各50μL,37℃避光孵育15min。(7)每孔加入终止液50μL终止反应。(8)测量波长450 nm处各孔的OD值,并根据标准曲线计算出样品浓度。
1.4 统计学方法 采用SPSS 16.0统计学软件进行统计处理。实验检测指标的数据资料以用表示,ATRA处理组、LE540处理组与对照组在不同时间点bFGF、TGF-β2分泌量的比较采用重复测量设计资料的方差分析,同时间点组间比较及不同时间点间的多重比较采用Dunnett t检验。P<0.05为差异有统计学意义。
2 结果
2.1 RPE细胞的原代培养及鉴定 原代豚鼠RPE细胞含有丰富的色素,体积大而扁平,呈多角形或圆形向周围伸展,可见细胞核分裂相(图1)。传代后色素减少,细胞近融合时形态近六角形(图2)。免疫组织化学角蛋白染色RPE细胞显示为棕黄色阳性反应(图 3)。

2.2 视黄酸和LE540对RPE细胞分泌的TGF-β2影响 加入ATRA和LE540后,实验组和对照组三者的比较结果有统计学意义,三者在 2、4、6、8、16 h TGF-β2的分泌量存在差别,F=297.776,P<0.001。ATRA与对照组比较,P<0.001;LE540与对照组比较,P<0.001。差别均有统计学意义。处理因素与时间有交互作用,P<0.001。加入10×10-6mol/L的ATRA后,2、4、6、8、16 h 时,TGF-β2的分泌量 ATRA 组较对照组均明显增加(t=28.178,P=0.000;t=37.133,P=0.000;t=25.839,P=0.000;t=7.283,P=0.005;t=10.271,P=0.002)。加入 5×10-6mol/L 的 LE540后,2、4、6、8、16 h 时,TGF-β2的分泌量 LE540 组较对照组均明显减少(t=-9.689,P=0.002;t=-21.714,P=0.000;t=-14.461,P=0.000;t=-33.906,P=0.000;t=-3.204,P=0.049)(表1)。
表1 ATRA和LE540处理组各时间点RPE细胞的TGF-β2分泌量比较 (pg,n=4)Tab 1 Comparison of TGF-β2 secreted by RPE cell in different time pointof thegroup treatedwith ATRA and LE540(pg,n=4)

表1 ATRA和LE540处理组各时间点RPE细胞的TGF-β2分泌量比较 (pg,n=4)Tab 1 Comparison of TGF-β2 secreted by RPE cell in different time pointof thegroup treatedwith ATRA and LE540(pg,n=4)
不同时间点上RPE分泌TGF-β2的水平组别 6 h 119.5±4.3 168.8±1.7 145.9±1.3 4 h 119.6±1.3 168.0±1.6 138.2±0.6 2 h 100.1±0.7 195.9±1.8 123.5±4.2 8 h 124.6±1.9 176.8±1.8 159.7±3.7 16 h 120.6±1.9 131.6±0.7 125.3±1.8 LE540组ATRA组对照组
2.3 视黄酸和LE540对RPE细胞分泌的bFGF影响 ATRA组、LE540组及对照组3者总体比较差别有统计学意义,F=914.432,P<0.001。LE540组、ATRA分别与对照组比较采取Dunnett t检验:LE540与对照组比较,P=0.178,差别没有统计学意义;ATRA与对照组比较,P<0.001,差别有统计学意义。处理因素和时间之间存在交互作用,P<0.001。加入10×10-6mol/L 的 ATRA 后,2、4、6、8、16 h 时,bFGF分泌量ATRA组较对照组均明显减少(t=-5.328,P=0.033;t=-7.348,P=0.018;t=-8.448,P=0.014;t=-25.062,P=0.002;t=-29.375,P=0.001)(表2)。
表2 ATRA处理组各时间点RPE细胞的bFGF分泌量比较pg,n=4)Tab 2 Com parison of bFGF secreted by RPE cell in different time pointof thegroup treated w ith ATRA(pg,n=4)
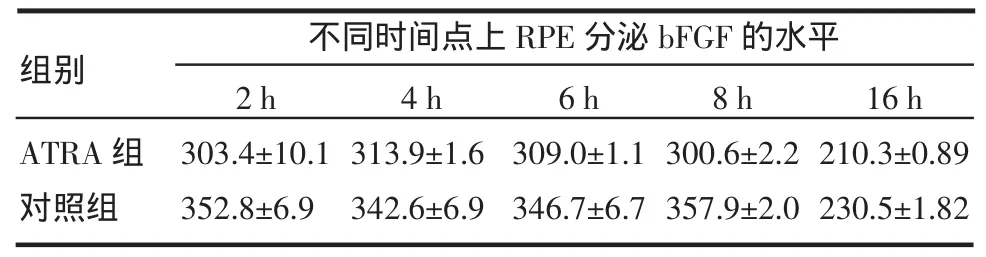
表2 ATRA处理组各时间点RPE细胞的bFGF分泌量比较pg,n=4)Tab 2 Com parison of bFGF secreted by RPE cell in different time pointof thegroup treated w ith ATRA(pg,n=4)
不同时间点上RPE分泌bFGF的水平组别 6 h 309.0±1.1 346.7±6.7 4 h 313.9±1.6 342.6±6.9 2 h 303.4±10.1 352.8±6.9 8 h 300.6±2.2 357.9±2.0 16 h 210.3±0.89 230.5±1.82 ATRA组对照组
3 讨论
近视发病机制的研究已成为近年来的热点。Heber和Mosest[2]较早阐述了“局部视网膜调控”机制,这种机制认为:视网膜感知外界环境变化后产生一级近视信号因子(如视黄酸、多巴胺、血管活性肠肽、胰岛素样生长因子等),并作用于视网膜色素上皮-脉络膜,使之产生二级信号因子(转化生长因子TGFβ、肝细胞生长HGF、碱性成纤维细胞生长因子bFGF等),从而启动了视网膜-视网膜色素上皮层-脉络膜信号转导系统,把局部视网膜信号转化为调控巩膜ECM重塑的信号,眼轴增长而近视形成[3]。
bFGF和TGF-β2已被证实与近视的发生发展密切相关,它们是RPE细胞分泌的细胞因子,此外RPE也分泌其他细胞因子[4]如色素上皮源性因子、白介素-1、肝细胞生长因子HGF等,这些因子又可以反作用于RPE细胞本身,影响其功能。对于近视形成的过程中,巩膜[5-6]、视网膜、脉络膜中[7]TGF-β2水平均降低,它的变化要早于近视巩膜的变化,但是还没有直接的证据说明TGF-β2的变化引起了近视的发生[8]。也有研究显示近视眼的视网膜、RPE-脉络膜复合体及巩膜的TGF-β2浓度升高[5],并且通过研究发现TGF-β2对巩膜发生重塑起到了重要的作用也影响到巩膜细胞的表型变化[6]。但都肯定了TGF-β2与近视发生发展密切相关,具体如何变化还需要大家探讨,我们的实验研究发现视黄酸引起TGF-β2分泌的增加,支持在近视发生过程中TGF-β2是近视促进因素的观点。在形觉剥夺的近视眼的后极部bFGF的表达明显减少[9],大量研究表明bFGF是实验性近视的终止信号,能够阻止视网膜神经上皮的凋亡,bFGF有可能成为治疗人类近视的突破点[10]。
视黄酸(retinoic acid,RA)是眼及视网膜感光细胞分化发育中不可缺少的物质,近年来研究发现RA与实验性近视的发生发展关系密切,RA可能作为一种信使通过某种机制来调控巩膜重塑而诱发形觉剥夺性近视[11],大量的研究表明RA确实促进了实验性近视的进展。RA受体属于细胞核受体,包括视黄酸受体(retinoic acid receptor,RAR)和视黄素X受体(retinoic X receptor,RXR),它们各有α、β和γ 3种亚型。而在视黄酸受体中,RARβ是由视黄酸诱导产生并受视黄酸自身调节,在中枢神经系统和眼球的发育中起着非常重要的作用,它通过与RA结合,调节关键酶基因的表达,抑制细胞DNA的合成,从而抑制细胞增殖,促进细胞分化。研究表明,RARβ可能参与鸡的形觉剥夺性近视的形成,形觉剥夺2周后雏鸡眼视网膜RARβmRNA的表达增高[12],体外实验同样证明了这点。对哺乳动物的研究发现形觉剥夺的眼的后极部视网膜、脉络膜巩膜的RARβmRNA的表达增高,对豚鼠巩膜细胞进行培养,发现RA抑制成纤维细胞的增生[13]。这些结果提示我们RARβ参与豚鼠FDM的形成,其变化可能受RA变化的诱导。并且有证据提示RA的变化要早于眼球形态学的改变[14]。但是最近通过对高度近视的群体研究发现:RARβ在高度近视的人群的基因序列并没有变化[15]。我们团队研究发现豚鼠后极部巩膜中RARβ蛋白含量改变与RA水平的变化趋势和方向相同。因此RA促进近视发展引起bFGF和TGF-β变化是否通过RARβ成为本实验的研究范围,为视网膜近视的信号通路的研究提供更为有力的证据。
本实验通过分别在培养的豚鼠RPE细胞加入ATRA和视黄酸β受体阻断剂LE540来观察其对细胞分泌bFGF和TGF-β2的影响,结果显示视黄酸能明显促进RPE细胞分泌TGF-β2,同时又明显抑制bFGF的分泌;培养的RPE细胞加入LE540后,TGF-β2的分泌量比对照组明显减弱,而bFGF的分泌没有明显变化。TGF-β2的分泌与视黄酸引起的作用正好相反,提示我们视黄酸引起的TGF-β2分泌变化是通过视黄酸β受体而实现的。bFGF的分泌受到视黄酸的影响,但是视黄酸β受体阻断对bFGF的分泌没有明显影响,说明视黄酸引起bFGF分泌的变化是通过其他受体起作用的,而非视黄酸β受体。
本研究认为全反式视黄酸能够调控RPE细胞分泌bFGF和TGF-β2,不过只有TGF-β2的分泌是通过视黄酸β受体而进行调节的,bFGF的分泌与此受体没有直接关系。我们推测,在近视发生过程中首先是视黄酸的分泌增加,然后RA进一步作用于RPE细胞,与视黄酸β受体结合来促进TGF-β2的分泌,TGF-β2或者直接起到促进近视发生的作用,或者通过拮抗bFGF而起作用。总之视黄酸β受体是视黄酸信号转导通路的重要分子,在近视的发生中起到重要作用。而RA是通过何种途径而引起bFGF分泌变化需要进一步探讨。本研究为探索近视形成的机制及药物治疗提供线索。但体内环境较体外复杂多变,LE540在体内是否会有相同作用尚有待进一步研究。
[1]胡诞宁,McCormick SA.视网膜色素上皮-脉络膜在近视发病中的作用[J].眼视光学杂志,2000,2(4):197
[2]Heber LU,Mosest K.Mechanisms of drosophila retinal morphogenesis:The Virtuesof Being Progressive[J].Cell,1995,81(7):987
[3]Stone RA,Pendrak K,Sugimoto R,et al.Local patterns of image degradation differentially affect refraction and eye shape in chick[J].Curr EyeRes,2006,31(1):91
[4]李臻,王震,荣翱.视网膜色素上皮细胞相关细胞因子[J].眼科新进展,2004,24(3):234
[5]Jobling AI,Nguyen M,Gentle A,etal.Isoform-specific changes in scleral transforming growth factor-βexpression and the regulation ofcollagensynthesisduringmyopiaprogression[J].BiolChem,2004,279(18):18121
[6]Jobling AI,Gentle A,Metlapally R,etal.Regulation ofscleral cell contraction by transforming growth factor-beta and stress:competing roles inmyopic eyegrowth[J].JBiolChem,2009,284(4):2072
[7]谢芳,陈跃国.双硫仑对鸡形觉剥夺性近视眼中转化生长因子-β2表达的影响[J].北京大学学报,2008,40(6):610
[8]毛俊峰,刘双珍,秦文娟,等.豚鼠近视眼视网膜Müller细胞中TGFβ2、VIP、DA 的表达 [J].眼科研究,2008,26(11):801
[9]Seko Y,Shimokawa H,Tokoro T.Expression ofbFGFand TGF-β2inexperientalmyopia in chicks[J].InvestOphthalmolVisSci,1995,36(6):1183
[10]Mao J,Liu S,Wen D,et al.Basic fibroblast growth factor suppresses retinal neuronal apoptosis in form-deprivationmyopia in chicks[J].Curr Eye Res,2006 ,31(11):983
[11]Mertz JR,Wallman J.Choroidal retinoic acid synthesis:a possible mediator between refractive error and compensatory eye growth[J].Exp Eye Res,2000,70(4):519
[12]Seko Y,Shimokawa H,Tokoro T.In vivo and in vitro association of retinoic acid with form deprivationmyopia in the chick[J].Exp Eye Res,1996,63(4):443
[13]吕勇,张金嵩,谢坤鹏.视黄酸对体外培养豚鼠巩膜成纤维细胞的影响[J].眼外伤职业眼病杂志,2005,27(3)∶164
[14]吕勇,张金嵩,闫磐石,等.RARβ在豚鼠形觉剥夺性近视眼中的表达[J].眼科研究,2006,24(4):411
[15]Yang D,Xiao Y,Dong S,etal.Association analysisof retinoic acid receptor beta(RARβ)genewithhighmyopia in Chinese subjects[J].Molecular Vision,2010,13(16):855
猜你喜欢
东方少年·布老虎画刊(2024年5期)2024-06-01 12:56:25
婚育与健康(2024年2期)2024-02-25 02:11:01
中国中医眼科杂志(2023年8期)2023-09-12 11:24:19
医学前沿(2021年14期)2021-10-28 09:16:47
阅读(低年级)(2019年6期)2019-08-27 04:29:53
中医眼耳鼻喉杂志(2019年2期)2019-04-13 05:23:50
中国眼镜科技杂志(2018年1期)2018-02-08 02:15:30
中华老年多器官疾病杂志(2016年9期)2016-04-28 08:52:37
中国老年学杂志(2015年19期)2015-12-29 03:11:59
军事文摘(2015年6期)2015-06-16 08:47:58
