
雷达在昆虫学研究中的应用
2011-08-27 03:59:42封洪强
植物保护 2011年5期
封洪强
(河南省农业科学院植物保护研究所,河南省农作物病虫害防治重点实验室,郑州 450002)
雷达在昆虫学研究中的应用
封洪强*
(河南省农业科学院植物保护研究所,河南省农作物病虫害防治重点实验室,郑州 450002)
昆虫雷达因能无干扰地远距离探测到自然状态下昆虫自由飞行行为,已被广泛用于昆虫高空远距离迁飞和近地飞行行为研究,成为昆虫学研究中一个卓越的研究工具。同时天气雷达也越来越多地用于鸟和昆虫的监测,探地雷达也在探测堤坝蚁穴中起到了重要作用。本文分析了各种不同类型雷达的性能和近年来在昆虫学研究中的应用,以昆虫学家的视角审视了雷达能为昆虫学研究做什么以及其中的挑战。
雷达; 昆虫; 迁飞; 目标识别; 应用
雷达(radar)是无线电检测和测距(radio detection and ranging)的缩写,是利用电磁波进行探测的电子设备,它发射电磁波对目标进行照射并接收其回波,从而测得目标的距离、距离变化率(径向速度)、方位、高度及目标形状等信息[1-3]。自从1949年被证明可以检测到单个昆虫以来,雷达不断地在昆虫学研究中得到应用,并逐渐演变出一系列专门设计的昆虫雷达,国际上形成了雷达昆虫学学科,还建立了雷达昆虫学网站(http:∥www.pems.adfa.edu.au/~s9104004/trews/ww_re_hp.htm)。
关于昆虫雷达的原理、类型、发展历史和对昆虫行为研究的经典贡献已有较为详细的总结[1,4-8],在此不再赘述。本文主要分析最近十年来用于昆虫学研究的雷达的性能、对昆虫学研究的贡献以及面临的挑战,并提出对我国雷达昆虫学发展的建议,激发更多昆虫学家的兴趣。
1 天气雷达在昆虫学研究中的应用
1.1 天气雷达鉴定昆虫目标的意义
敏感的微波多普勒天气雷达经常观测到边界层的晴空回波(clear-air echoes),晴空回波的产生原因自1940年代雷达应用之时开始就一直存在争论[9]。晴空回波主要来自于两类散射体或其混合体:(1)产生布拉格散射的折射率的梯度变化(折射率波动)或(2)产生粒子散射的空中粒子,包括空中生物(如昆虫、鸟和蝙蝠等)、灰尘与其他粒子[9-12]。
布拉格散射是大气中折射率波动产生的散射,主要来自于湿度在雷达的半波长尺度上的变化或更小程度上温度的变化[9],这是频率在3GHz以下的雷达的主要目标。而鸟和昆虫等粒子散射体是更高频率的雷达的主要目标[11]。这两种散射对正在运行的天气雷达都普遍存在,如美国的3GHz的多普勒天气预警雷达(weather-surveillance radar-1988,doppler,WSR-88D)[11]。通常飞行能力弱的非迁飞小型昆虫在X波段雷达上的横截面约为10-10~10-7m2,而大型昆虫为10-5m2以上[11],因此典型的天气雷达因其分辨率空间(resolution volume)大而观测不到单个的昆虫,但当昆虫的密度足够大时作为体积散射目标确实可以被这些天气雷达“看”到[11]。
在陆地上空从春到秋(甚至在冬天气温超过10℃时)昆虫回波经常出现在晴朗的天空,它们可以被看做不期望的杂波,也可以为被看做是需要的(尽管不是完美的)晴空示踪者来测风[11,13]。使用昆虫回波测风的可靠性取决于:(1)用一个正确的方法将昆虫回波标记出来,从而最大限度地得到好的数据和减少杂波干扰;(2)如何估计测风的误差,包括由昆虫自主飞行产生的测风偏差[14]。气象雷达学家近年来的研究主要集中在如何识别空中生物回波以及昆虫回波是否可以用来测风上[13-14]。
1.2 天气雷达鉴定空中生物的原理
一部多普勒测云雷达通常测量雷达基本反射率(base reflectivity)、径向速度(radial velocity)和径向速度的多普勒谱宽(width of spectrum)[15-16],如美国的 WSR-88D雷达网的二级数据[16]。用这些信息可以探测到空中的生物,如鸟、蝙蝠和昆虫。
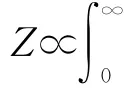
径向速度是用于测量目标朝向或背离雷达的运动分量[16],是根据多普勒效应计算得来的。所谓多普勒效应就是当声音、光和无线电波等振动源与观测者以相对速度V运动时,观测者所收到的振动频率与振动源所发出的频率有所不同。由多普勒效应所形成的频率变化叫做多普勒频移,它与相对速度V成正比,与振动的频率成反比[2]。速度方位显示(velocity azimuth display,VAD)是(假设风场是线性的)对每一个距离档上各个高度的径向速度进行正弦曲线拟合得到的[13]。利用VAD与当地的风速进行比较可以鉴别回波是否为昆虫[13]。使用该方法测到1.2~1.4km高度上的目标为昆虫,它们的速度为18m/s,比同高度上的雨滴快3m/s[17]。
多普勒谱宽用于测量脉量内目标的多普勒速度的变异,如果一个脉量内所有的目标都以相同径向速度飞行,谱宽将达到最小值,如果一个脉量内有多种生物目标以不同的径向速度飞行,谱宽就会很大[15-16]。大多数昆虫的空中速率都较小而鸟的飞行速率较大,因而大气中以昆虫为主时谱宽较小,以鸟为主时谱宽较大[16]。
双极化(dual-polarization)雷达发射和接收水平(H)和垂直(V)极化的电磁波,这使得我们可以估计雷达目标的几何形状[18-20],因此双极化雷达比单极化雷达能更有效地鉴定目标,而且可确定昆虫是否具有定向行为以及定向是否与风向一致[13-14,21]。用于区分生物目标的双极化变量包括差反射率因子(differential reflectivity)ZDR、微 分 相 位 (differential phase)δ、相互垂直的极化回波的相关系数[16,20,22]。
差反射率因子ZDR指水平和垂直频道的反射率比值并以对数表示,即ZDR=10log(Zh/Zv)(dB),其中Zh和Zv分别指水平和垂直极化的反射率因子[9-10]。对于M次脉冲每一分辨率空间ρhv就有M个谱系数的测量值[22]。令κ为一系列取值从1到M的谱系数的序号,κ可以转化为相对应的径向速度,然后,Sh(κ)和Sv(κ)分别为水平和垂直频道的包含了信号与噪音在内的复杂谱系数,那么微分相位δ(κ)=arg(〈Sh(κ)Sv(κ)〉)[20],其中,双极化雷达ρhv可由水平和垂直频道的复杂谱系数的三点平均求得[18,22]。
由于昆虫和鸟是似椭圆体的目标,鸟的宽与长之比为1∶2~1∶3,昆虫的宽与长之比为1∶3~1∶10[9],垂直极化将给出一个各向同性的扫描,而水平极化扫描昆虫侧面时得到较强的信号,因此鸟和昆虫的ZDR值比水滴小。一般来说,降雨的ZDR值为0~2dB[10],鸟的ZDR值为-1~3dB[20],昆虫回波的ZDR值为5~10dB[9-10]。当昆虫具有共同定向时,ZDR值可用于将昆虫从其他目标中区分出来[13],而且比较ZDR值还可确定昆虫是否在进行迁飞[9]。
使用10cm波长的垂直分辨率为2.5m的高分辨率S波段调频连续波雷达(FMCW)可以将小型昆虫(<10mm)从布拉格散射体中分离出来[11]。这次FMCW观测结果与毫米波雷达观测到的结果[23-24]一致,但观测到的平均反射率因子远小于其他观测[9-10],可能是由于其他观测正好赶上昆虫密度特别大的时期[11]。
毫米波雷达(3mm和8.6mm或94GHz和35GHz)由于时间和空间分辨率高,使气象学家可以探测到更小的冰晶来研究大气的微观结构[12],也同样有利于观测微小昆虫。Luke等人用35GHz毫米波测云雷达(MMCRs)辅以气球和风筝携带的遥控捕虫装置证明从地面至700m高度充满了小型昆虫(体长约2mm,翅长4~5mm,翅宽1~2mm),发明出一种以35GHz毫米波测云雷达MMCR记录的多普勒谱、描述昆虫谱的特征提取机和神经网络算法提取小型昆虫回波的技术[12]。只用 MMCR多普勒谱作为输入时这项技术能鉴定出92%以上的昆虫回波。
1.3 使用天气雷达区分鸟与昆虫
大量的文献表明鸟和昆虫都是晴空回波的主要来源,气象学家区分鸟与昆虫是由于他们认为迁飞的鸟会对测风造成严重的偏差,而认为昆虫造成的偏差较小[25]。可是没有一种可靠的简单方法把昆虫与鸟区分开,鸟类学家用雷达时经常假设他们的目标是鸟,而昆虫学家则经常假设为昆虫[25]。使用天气雷达在一定程度上可以区分鸟与昆虫,要达到百分之百的鉴定昆虫目标是不可能,因为昆虫回波的所有参数都与其他类型目标的参数有重叠[13]。
每一分辨率空间可以用极化雷达的一系列极化参数进行描述,根据这一空间内目标的种类建立各种参数的描述函数,可以对不同目标进行分类[18]。根据反射率大小似乎可以区分鸟与昆虫,但由于昆虫在空中的密度会非常大,高密度的昆虫可以产生很高的反射率,也就是说较低的反射率可以排除鸟的可能,但高的反射率却不能排除昆虫的可能性,因此单以反射率不能区分鸟与昆虫[25]。
在早期的利用极化雷达鉴定空中目标的研究中发现,ZDR值可用于区分鸟与昆虫[20,26],但是 Mueller认为鸟的ZDR值高于昆虫,而后来的研究却认为恰恰相反,即昆虫的ZDR值大于鸟[18,20,22]。这是由于昆虫和鸟的体型都有很多变化,他们的ZDR值的变化范围相互重叠,很难说哪一类生物的体型更长[18,25]。同样原因δ值也无法将昆虫与鸟区分开[16]。如果两种或更多种散射体同时存在于同一个分辨率空间,这个空间的极化变量取决于混合的比例、它们的排列、生物目标的定向、在这一空间内的位置(均匀填充、成层、成团或其他)。因此当多种类型的目标同时存在于一个分辨率空间时,极化变量的变异范围就会变大,假设一种目标为主体得到的模糊分类标准就难以奏效[16,18]。
最近建立的双极化雷达的速度方位显示谱图(SVAD)与三维(方位、多普勒频移和极化谱密度)方 位 谱 图 (3-D spectral azimuthal histogram,3DASH)为区分目标种类提供了新的视觉分析技术[18,22]。SVAD是在VAD图基础上以不同灰度表示出多普勒系数以及极化变量(ZDR、δ、ρhv)的谱密度图,这种谱图可以在同一取样空间内同时存在鸟与昆虫时将两者的回波区分开[22]。3DASH是一定距离档上的方位—速度—δ分布的三维图,并用灰度表示发生概率。它可以投影成方位—速度、方位—δ和速度—δ图,通过设置不同的透明度可以除去干扰看清目标[18]。3DASH的螺旋结构印迹代表了不同目标的特性,甚至有望区分不同类型的昆虫[18]。
不同波段的雷达同时观测,根据目标在不同波段的雷达上的反射率差异可以确定目标的种类并容易将昆虫与鸟区分开。例如DOW3(3.198cm)、WSR-88D(10.0~11.1cm)和 UMASS(0.315 7cm)的联合使用[25],S波段(10.7cm)、X 波段(3.2cm)和 C波段(5.4~5.5cm)雷达的联合使用[9]。激光雷达的波长比雨滴和昆虫小,对由高密度的微小雨滴(10μm)组成的云非常敏感,但一般探测不到低密度的昆虫。因此英国人曾利用两部毫米波(35GHz和94GHz)垂直雷达、一部厘米波垂直昆虫雷达与激光雷达同时使用来区分昆虫目标[15]。
1.4 天气雷达观测到的空中昆虫行为
昆虫,尤其是自主飞行能力很弱的微小昆虫(通常体重为1mg[15]或体长小于10mm[12])白天借助由于地面受热产生的上升气流升空,其上限是对流边界层(convective boundary layer,CBL)顶[13]。晚上地面冷却形成逆温层,行星边界层比较稳定,夜间迁飞的昆虫个体一般较大(>40mg),有较强的自主飞行能力,用来测风误差较大,因此大多数气象雷达对昆虫的观测都集中在白天[13]。
英国的一部毫米波垂直多普勒雷达观测到小型昆虫的上升速率可达4m/s,可上升到高出CBL 800m以上的高度[15]。大多数小型昆虫(蚜虫、飞虱)自主飞行的上升速率都小于0.2m/s,即使是大型昆虫(蝗虫和夜蛾)其上升速率也只有0.4~0.5m/s,那么观测到的4m/s的上升速率肯定是昆虫与上升气流的速率之和[15]。飞行高度超出CBL表明昆虫是一种主动飞行者而不是单纯靠上升气流飞行的[15]。
还有研究表明尽管循环气流可以将昆虫带到更高的地方,但昆虫层顶却在1 800mAGL,那儿的温度为10~15℃,低于这个温度蝗虫是不飞行的[10]。如果高于这个高度,由于昆虫对气温的反应,会通过折叠翅膀停止飞行甚至向下飞行主动下降使自己保持在细线(气象雷达观测到的昆虫带或层)内[10]。机载W波段(3mm)垂直雷达的观测结果却表明使CBL内昆虫产生反应的是突然的垂直运动而不是温度[24]。
气象雷达曾多次观测到大量迁飞的昆虫具有共同定向特性,而且头向与风向不一致[10,13,19]。有趣的是英国用多普勒雷达观测到昆虫的定向在6月份更偏北一些,而在8月份更偏南一些,这与迁飞的季节性偏好方向一致[13]。
2 昆虫雷达在昆虫迁飞研究中的应用
由于用传统的昆虫学方法很难观测高空中迁飞的昆虫,20世纪60年代末开始使用专门设计建造的昆虫雷达,这极大地丰富了我们对昆虫迁飞行为的认识[1,5,7-8,27]。目前各国普遍采用的昆虫监测雷达(insect monitoring radar,IMR)为能长期自动化运行的旋转极化的垂直昆虫雷达(vertical looking radar,VLR)[28-31]。
2.1 旋转极化的垂直昆虫雷达
昆虫监测雷达或垂直昆虫雷达(IMR或VLR)的天线不像传统雷达那样以各个仰角进行360°扫描并将结果在平面显示器(plan position indicator,PPI)上显示出来,而是产生一个垂直的波束,当昆虫飞过波束时对昆虫进行测量[28]。实际上1949年Crawford首次证明雷达能探测到昆虫回波时就是这样做的,这种新型昆虫雷达的原理和技术在20世纪80年代初就确定了下来,90年代的“数字革命”给昆虫雷达的发展带来新生,迅速发展的廉价数字计算机以及电子设备使制造低成本的自动运行的昆虫雷达成为现实[28]。美国的Beerwinkle首先研制了一个简单的垂直雷达并无人值守地运行了两年,英国和澳大利亚也紧跟着研制出更为先进的垂直雷达,并从上世纪90年代末开始了长期的自动观测[28,32]。
旋转极化的垂直昆虫雷达发射一个环形对称垂直向上看的波束,并通过机械转动波导馈电使波束内的线性极化平面旋转。馈电与垂直方向有一个非常小的0.2°的偏角,这使得波束产生轻微的摆动。于是整个雷达波束就像陀螺旋转变慢时一样产生旋转的锥形扫描[29]。由于馈电的垂直偏角非常小,能以300r/min的转速高速转动,因此当昆虫穿越垂直的雷达波束时可以完成几个完整的旋转扫描[28]。由于波束的位置和极化在任何时候都是已知的,接收到的回波信号包含了目标的轨迹(相对于垂直方向的位置)和由极化平面旋转带来的背向散射截面的变化两方面的信息[28]。因此这种扫描的效果相当于一个目标飞过头顶时以不同角度去反复“切割”它,得到昆虫不同截面的信息。
这种昆虫雷达的旋转极化配置(多个“切面”)与天气雷达的双极化配置(两个“切面”)相比,能采集到目标更丰富的体形信息,大大增强了对昆虫的识别能力。这种雷达检测到的回波能量可以用一个非常复杂的公式表示,包含了目标的水平速率、位移方向、身体的轴向、到波束中心线的最近距离、昆虫的大小和两个代表昆虫体形的参数在内的7个参数[28-29]。由于这种雷达每一圈扫描都可以得到64个回波能量数据,这些数据足够用于拟合7个参数,拟合的回归系数可达0.9以上[28-29]。具体的数据拟合方法十分复杂[33-35],但随着计算机技术的突飞猛进,能在现代微型计算机上运行的C++写成的程序可以在4min内自动分析5万个昆虫信号[35]。将模拟的昆虫回波信号与实测的昆虫回波信号相减得到的残差并不为0,也不是随机分布的,而是出现有规律的大小相似的波动,每一个波峰就代表了一次振翅,根据这一原理可以将昆虫的振翅频率提取出来[36]。
1.3观察指标 根据线性视觉模拟评分标尺对患者舒适程度进行评估[3],分为无症状、轻度、中度、重度不舒适,分别为0分、1~4分、5~7分、8~10分。自行设计调查表,对患者进行护理满意率调查。
由于垂直雷达是垂直向上照射的,假设昆虫在飞过雷达波束时不主动(或因气流颠簸被动)地改变倾角、飞行偏角和滚动倾斜角,所有的昆虫都以相同的姿势飞过电磁波束,因此提取出的4个关于昆虫本身特性的参数(昆虫的个体大小、振翅频率以及2个体形参数)就可以有效地鉴别昆虫的种类[37-39]。英国昆虫学家则使用最大背向散射截面δxx与最小截面δyy(与δxx垂直)的比值来鉴定昆虫种类[38-39]。较大的比值代表较细长的体形,如15∶1代表草蛉,较小的比值代表较短的体形,如5∶1代表步甲,而1∶1代表瓢虫[29]。澳大利亚科学家用两个体形参数(α2和α4)描述昆虫的共极的线性极化类型(copolar-linear polarization pattern,CLPP),理论预测与实际观测的结果一致[37]。用α2和α4描述的CLPP由于有丰富的图形变化(2个瓣至4个瓣及瓣的不同形态)可能代表了更为丰富的昆虫体形信息,可用于昆虫种类的详细鉴定[37]。但实验室测定一种昆虫时得到的CLPP形态变化也较大,这就给CLPP鉴定种类带来了困难,还需要进一步的研究。
现行的IMR或VLR还不能连续地处理信号,即不能连续的观测垂直空间,而是把垂直高度分为若干个距离门(range gate)即高度带,两个距离门之间有一定的间隔[29]。澳大利亚的IMR在垂直方向上有8个距离门,每个距离门的高度(即分辨率)为50m,距离门的间隔为150m,由于第一个距离门含有大量地物回波干扰成为无效的距离门,因此其有效的观测高度为200~1 400m[28,34]。英国 VLR有15个有效的距离门,门高45m,间隔26m,有效观测高度为150~1 166m[29]。因昆虫的大小不同实际能探测到的高度(距离)也不同,1mg的昆虫只能在200m以下被探测到(但由于地物干扰实际上是探测不到),2mg的昆虫的探测高度可达350m,5mg的可达550m,15mg的可达1 200m[29]。
随着现代通讯技术的发展,远程遥控的多机联网自动观测、甚至与现行的天气雷达或雷达网进行联合或联网观测也成为现实,监测结果可通过Internet向大众实时发布[40]。
2.2 传统扫描雷达的新生
旋转极化的垂直昆虫雷达虽然在目标鉴定上有着无可比拟的优势,但它的取样空间小而且200m以下无法观测。传统的扫描昆虫雷达取样空间大,但信号数字化和分析困难,这也是国际雷达昆虫学界转而开发垂直昆虫雷达的主要原因。近年来随着信息技术的迅速发展扫描雷达的数字化取得了长足的进步,使传统扫描昆虫雷达重获新生[1,41-42]。
传统扫描昆虫雷达依靠对雷达屏幕显示的回波进行照相和摄像来记录数据,以供后期人工分析用。澳大利亚的Drake博士详细描述了手动分析扫描昆虫雷达数据的方法和过程[43]。1998年我国第2部昆虫雷达(KC-2)建成,为了克服信号采集与分析的困难,在郭予元院士和吴孔明博士的大力支持下程登发博士开发出了扫描昆虫雷达实时数据采集、分析系统[41-42]。
KC-2昆虫雷达的显示单元提供了一个8针VGA信号接口,供连接其他计算机显示器用。使用基于PCI总线的8bit高速度图像采集卡可直接从雷达主机将PPI采集为分辨率为768×536的8位黑白图像。使用VC++编程实现了PPI图像的采集、存贮与再现,并根据Drake的方法实现了昆虫回波的自动识别和密度统计。该系统还可将连续采集的3张图片叠加在一起分别以红、绿、蓝3种不同的颜色显示回波点,使用户可以清楚地分辨出地物回波(白色)以及昆虫的位移轨迹,并能测量出位移速度[1,41-42]。将测到的昆虫位移速度与同时实地测得的风速进行矢量差运算得到昆虫的定向[1,44]。
由于这套系统大大提高了扫描昆虫雷达的工作效率,使我国昆虫学家得以进行长期的雷达昆虫学研究,先后观测了甜菜夜蛾、草地螟、棉铃虫和黏虫等重要害虫以及黄蜻和毛婪步甲等天敌在我国华北地区的迁飞行为,获得了重要的行为参数[1,44-54]。这部雷达以及数据采集与分析系统直到今天还在正常运行,是标志着我国独立进行雷达昆虫学研究的一个里程碑。
无独有偶,加拿大科学家最近开发了一个开源平台radR,用于采集、存贮和分析航海雷达以监测模式观测到的生物目标[55]。底层数据处理是由C代码实现的,用户定义的函数是由R语言写成的,可以在计算时按预设的步骤调用。输出数据格式设计的可以在将来增加数据而不必改变C代码。目前这个开源平台支持两种采集卡,可以在 Windows XP和Linux两个操作系统下运行。
2.3 迁飞昆虫的定向与成层
昆虫的共同定向是指高空迁飞的昆虫头朝着相同的方向飞行的行为,是迄今为止昆虫雷达观测到的最神奇且神秘的行为现象[8,27,29,56]。自从 1968年Scheafer首次用雷达观测到夜间迁飞昆虫具有共同定向以来,这种行为在各个地区和各种夜间迁飞的昆虫身上反复地被观测到[27,56-59]。由于在夜间高空飞行,视觉的定向是不可能的,两种最可能的机制是“风定向”和“罗盘定向”[8,57-59]。
风定向机制认为昆虫可以用触角探测到风向,以便始终向下风方向定向[8],那么当不同高度上的风向发生转变时,昆虫的定向也应跟着转变。有趣的是一个模拟模型预测的结果是不同高度上风向的转变(Ekman螺旋)误导昆虫在北半球向右偏而在南半球向左偏[60]。英国观测到的数据支持模型预测的结果,即北半球昆虫顺风定向时向右偏[60],但澳大利亚的观测结果却不支持模型的预测结果(王海扣,个人交流)。同样处在北半球的中国观测到的棉铃虫秋季回迁中当在下风方向大于225°(西南方向)时向左偏,而当下风方向小于225°时向右偏[47],这同样也不支持模型的预测结果。如果昆虫总是顺风定向那么迁到高纬度地区繁殖的昆虫的后代,很可能由于所谓的Pied Piper效应而滞留在高纬度地区,无法安全回到低纬度地区越冬,然而近年观测到昆虫回 迁 的 事 例 越 来 越 多[44-45,47-48,51,61-62],这 表 明昆虫不是盲目顺风定向、顺风迁飞的。
罗盘定向机制认为昆虫以某种罗盘识别地理方向,迁飞时总是向着自己偏好的方向定向。近年来越来越多的观测表明夜间迁飞的昆虫具有这种非常惊人的罗盘定向能力[44,47-48,51,62-64]。我国的草地螟春季迁飞时向东北定向,黏虫秋季回迁时向西南定向,当风力较弱时这种主动定向可以对迁飞轨迹产生较大影响,但当风力较强时这种校正起到的效果甚微[48-49]。除了黄蜻飞行能力较强可以侧顶风飞行外[51],总体上昆虫的飞行能力较弱,选择与下风方向呈锐角顺风飞行。但这并非是被动顺风飞行,棉铃虫秋季总是向着西南方向偏转,而且选择有适合的风向出现的夜晚或高度飞行[45,47]。昆虫的定向是有季节适应性的,即在北半球春季向北而秋季向南[62],它的生态学意义是使昆虫能更快地到达适生区繁殖或安全越冬[61]。
夜间迁飞昆虫的成层也是使用昆虫雷达经常观测到的一个现象[8]。在高纬度地区的观测发现昆虫的成层与逆温有关,由于昆虫有最适飞行温度,在温度较低的环境里经常聚集在逆温层顶飞行[49,51,65-67]。当温度较高时夜间迁飞的昆虫选择风速最大的高度成层,这使昆虫以最小的能量实现最大位移[45,47,61-62,64],我国还发现昆虫的成层与风向和风切变有关[45,47]。因此,昆虫的成层可能与温度、风向、风速和风切变等多个因素有关。
3 谐波雷达与近地昆虫行为研究
3.1 谐波雷达原理
昆虫谐波雷达的原理是发射高能的微波照射粘在单个昆虫身上的转发器(transponder)并接收由转发器转发回来的二次调谐波来探测到昆虫目标[7,68]。由于植物、地物和海洋不产生高次谐波而由它们产生的基波又被抑制,故谐波雷达具有很强的抗杂波能力,能有效地排除地物杂波的干扰[69]。根据这一原理现在已建成了用于研究昆虫爬行行为的便携式谐波雷达和用于研究昆虫近地飞行行为的基站式谐波雷达[69]。
使用谐波雷达需要在昆虫身上粘一个转发器,类似于标记回收试验所用的标记物,也称标签(tag)。这种标签由一个肖特基二极管和一段天线组成[68,70],肖特基二极管能够吸收高频电磁波的部分能量并反射出二次谐波,不需自带电源,故可做得很小很轻[7,69],不会影响昆虫本身的行为[71-73]。
所有使用谐波雷达的昆虫学家都要自制标签,在标签的探测性能与耐久性之间找到一个平衡。自制或修改标签包括改变二极管、天线、二极管—天线配置和天线配置(直线形、圆形和卵圆形)[74]。天线长度理论上是雷达波长的一半,提高雷达的频率可以有效地缩短标签的长度。利用经济的航海雷达主机制造的昆虫雷达系统由于使用的频率是9.4GHz,因此可以使用重量小于3mg,长度约十几毫米的标签[7,68,73,75]。到目前为止最轻的标签(0.4mg)是由Roland等制作的,尽管这种标签在野外的寿命是非常有限的[69]。
虽然标签越小对昆虫行为的影响越小,但使用较小的标签就要提高雷达的频率,频率越高的电磁波穿透力就越弱,因此要根据昆虫的行为特点及所要研究的特性来决定使用什么样的谐波雷达和标签。
3.2 便携式谐波雷达
常用的便携式RECCO收发系统,重1.6kg,是寻找雪崩遇难者的雷达系统(www.recco.com),使用917MHz的初始电磁波,可穿透作物、雪和土壤,使用RECCO配发的标签可以探测到土壤下1m处的目标[70,74]。谐波雷达的探测性能与标签有关,使用O’Neal等的16cm标签,RECCO只能探测到土下30cm处的目标[74]。由于商用的RECCO收发系统无法改变电磁波频率,这就限制了使用更小的转发器。如果研究的对象个体较大,运动速度不快且有隐藏于土壤或茂密的作物丛中的习性如步甲,使用商用的RECCO收发系统和十几厘米长的标签能得到令人满意的结果[74,76-79]。2004年 Williams等人还在北京研究了光肩星天牛的飞行行为取得成功,其中最远移动距离超过了90m[70]。
为了研究马铃薯甲虫的运动行为,加拿大的Colpitts研究小组研制了频率为9.4GHz的便携式谐波昆虫雷达和相应的重3mg长16mm的电子标签,并详细描述了标签的制作和标定方法,这种系统还可用于研究与马铃薯甲虫相似的多种有益昆虫的行为[68,75,80]。使用这套系统在马铃薯田探测113头被电子标签标记过的马铃薯甲虫的发现率为87%,平均每头用时(165±15)s,在距离苹果树3m处能发现78%的被标记的李象鼻虫,当被标记的玉米根萤叶甲在正对雷达的玉米叶、雄穗和茎秆上时,在6~20m距离处的发现率可达80%~100%,但当玉米萤叶甲在玉米雌穗或茎秆背后时几乎不能被发现,在第二行玉米上的叶甲的发现率也明显下降,但如果使用者可以绕过作物进行扫描,发现率会非常高[81]。
昆虫行为是否会因虫体粘上了电子标签而受到影响一直是广受质疑的。Boiteau等的研究发现,被标记的3种甲虫的确会因增加了重量负担或标签的缠绕爬行速度受到一定的影响,影响因种类而异。但这种负面影响远比环境本身对昆虫行为的影响小,而且这轻微的影响与使用此技术获得的昆虫的重要生态学信息相比是微不足道的,便携式谐波雷达不失为一种优良的研究手段[72,82]。桂连友最近使用这种技术研究了饥饿马铃薯甲虫的行为,发现饥饿能增加甲虫的运动距离而且这一影响比试验室研究的结果更强[83]。他从加拿大学习回国后也建造了相同的便携式谐波昆虫雷达,正在开展橘大实蝇行为的研究[84]。
3.3 基站式谐波雷达
便携式谐波雷达从某种意义上讲是一种“方向发现”装置,即发现标记昆虫的方向,它不能很快地绘出昆虫飞行的轨迹,因为它的探测距离一般小于50m,操作者必须能够跟得上昆虫的爬行或飞行才行[7]。1995年英国的Riley领导的雷达昆虫学研究团队研制了世界首部真正的谐波昆虫雷达。这部基站式谐波昆虫雷达有两个碟形的天线,下部的一个直径1.5m的抛物面天线用于发射功率25kW、脉冲宽度0.1μs、波长3.2cm的电磁波,上部的第二个直径0.7m的抛物面小天线用于接收反射回来的1.6cm波长的谐波,两个天线都以20r/min的转速旋转,可以实时绘制出半径约900m范围内飞行高度在3m以下的标记昆虫的飞行轨迹,空间解析度为5m[8,73,85]。
由于社会性昆虫蜜蜂(或其他的蜂)有“家”的概念,每次外出都要重新回家,用电子标签标记蜜蜂一般不会丢失,同时蜜蜂的行为也充满了不解之谜,所以成为基站式谐波昆虫雷达研究的主要对象[8]。研究发现,初次出巢的工蜂通过反复飞行训练逐渐增加离巢距离从而保持回巢的能力,这种距离的增加是由于飞行速度的增加而不是飞行时间的增加,这种飞行行为的渐变与出巢的次数有关而与工蜂的年龄无关。熊蜂和蜜蜂都能正确地从觅食处径直回到蜂巢,即使正在觅食的工蜂被人为移动了位置也会自动按既定的方向和路线飞行,飞行过程中可以克服风的飘移,当蜜蜂准确定位蜂巢或食物源时进行随机寻找。研究还证明了von Frisch的蜜蜂舞蹈理论的正确性,即舞蹈交流包含了方向和距离两方面的信息[8]。由于这种谐波雷达的出色表现,最近德国也请洛桑研究院的雷达工程师为他们建造了世界第二部基站式谐波雷达,用于蜜蜂行为的研究,明确了蜜蜂使用一系列地面标志物作为测量距离的标尺[86]。
尽管许多黄地老虎飞出雷达的视野,用于研究几百米范围内雄蛾寻找性信息素源的行为也取得了成功,发现当气味浓度足够大时,顺风飞行的雄蛾转为侧风飞行而侧风飞行的则转为逆风飞行[87-88]。首次用谐波雷达研究5种蝴蝶近地面飞行行为发现了快速的直线型和慢速的包括循环觅食的非直线型的两种飞行轨迹,展示了用于蝴蝶飞行研究的潜力[89]。庆网蛱蝶(Melitaeacinxia)的飞行能力不强,2h飞行距离的中位数是488m(最短9m,最长2 730m),而且喜欢在开阔地和林地的边缘飞行,因此试验在由一圈树围成的开阔草地上进行,成功证明了生活在片段化生境下的种群比生活在连续生境下的种群飞行扩散能力更强[90-91]。
4 探地雷达在蚁巢探测中的应用
探地雷达(ground penetrating radar,GPR)又称地质雷达或透地雷达,是用无线电波来确定地下介质分布的一种方法,在考古、市政建设、建筑、铁路、公路、水利、电力、采矿、航空等领域都有广泛应用。中国科学家创造性地将其应用于堤坝白蚁巢的探测,在堤坝隐患排查中发挥着作用[92-94]。
探地雷达由主机、收发天线、图像处理显示系统几个部分组成。实际探测过程中天线沿地面移动,脉冲信号不断地被发射和接收,显示系统经A/D转换后得到的数据信号按一定方式进行编码排列及处理,以二维形式给出连续的地下纵向剖面图像[92]。电磁波在地下介质中传播时遇到存在电性差异的界面时发生反射,根据接收到电磁波的波形、振幅强度和时间的变化特征推断地下介质的空间位置、结构、形态和埋藏深度。根据探地雷达的影像特征可以区分出蚁巢、菌圃群、蚁道、树根、红火蚁巢、金属物等不同的目标[92,95]。
目前用于探测堤坝蚁巢的探地雷达都是由美国地球物理勘探设备公司(http:∥www.geophysical.com)生产的SIR(subsurface interface radar),该雷达可配备16~2 600MHz范围内不同波段的天线。探测深度与土壤结构和含水量等因素有关,同样条件下使用的电磁波频率越高探测深度越浅[92,94]。使用SIR-3000GPR 400MHz的天线可探测到地下半米左右深度的直径在10cm左右的蚁巢[94-95]。
5 我国昆虫雷达学的机遇与挑战
几乎所有的雷达昆虫学研究都是以控制重要经济害虫为主要目的的研究项目的副产品[96]。我国的雷达昆虫学研究也不例外,新中国成立后东北黏虫的变幻莫测促使我国昆虫学家建立了第一部扫描昆虫雷达,棉铃虫的大暴发促生了第二部昆虫雷达,进入新世纪草地螟和稻飞虱大暴发,我国又投资建造了多部垂直昆虫雷达、毫米波昆虫雷达和多普勒调频连续波垂直雷达[4,6,53,54,97-102]。经过十几年的迅速发展,我国已形成了以垂直昆虫雷达为主要机型的昆虫雷达网[6],同时雷达昆虫学研究团队也不断壮大,观测资料不断积累。在今后的发展中应充分发挥多种雷达共同探测和联网探测的优势,努力完善新型雷达软件系统,深入分析已有的观测资料,不断提高我国雷达昆虫学研究和应用的水平。
5.1 新型昆虫雷达(网)的建设
出于不同的需求我国近年来研制了垂直昆虫雷达(包括简单的垂直雷达、旋转极化的垂直雷达和收发分置的多普勒垂直雷达)。但这些雷达的信号采集与分析却一直存在问题,如只能测到昆虫的高度与密度[97-101],要得到昆虫的更多信息必须进一步完善数字化采集与分析系统。最近发展起的开源radR平台[55]有助于改进我国扫描雷达的信号采集与分析系统。我国首创了探地雷达探测蚁巢的方法,如果能进一步研究用来调查地下害虫(或在地下的虫态)的密度与变化规律,将对地下害虫以及害虫地下越冬规律的研究起到不可估量的推动作用。
我国目前虽然拥有世界上最庞大的昆虫雷达网,但由于没有支撑其运行的软件,雷达网尚未得到充分利用。即便是有软件支持运行,由于各部雷达相距太远[6],也无法形成有效的覆盖。天气雷达网经常探测到的昆虫,对气象学研究是污染或干扰,但对昆虫学家却是非常有用的[103]。虽然现行的气象雷达不能识别昆虫的种类,但可以监测到昆虫是无定向的近距离扩散还是进行有定向的远距离迁飞,单部雷达可监测10~30km甚至50km远和1~2km高[13-14],其覆盖范围也远比昆虫雷达网广。
早在1988年芬兰人就开始用天气雷达、诱虫网和轨迹分析模型研究蚜虫和小菜蛾向芬兰的迁飞,并利用手机短信和计算机网络向试验用户发送害虫入侵警报[104,105]。事实上,美国的天气预警雷达网也越来越多地用于昆虫密度和昆虫迁飞的常规观测[24]。因此,可以设想将我国的天气雷达网、昆虫雷达网和诱虫灯网相互结合形成能预测害虫发生和迁飞的实用网络[15,106],但如何协调各部门联动是一个新的挑战。
5.2 毫米波与厘米波昆虫雷达的协同观测
2007年我国第一部波长为8.0mm的扫描昆虫雷达建成以来,不但观测到了稻飞虱的空中密度、飞行高度、空中密度随时间的变化等预期现象,而且还观测到了哑铃形回波[97-99]。但关于毫米波雷达观测到的哑铃形回波是不是由稻飞虱的共同定向产生的存在很大争议。
Mie散射能计算出任何物体在任何雷达波长下的回波强度,当粒子小于波长的1/10时近似于Rayleigh散射,即回波强度与球体直径的6次方成正比[25]。对于传统的3.2cm昆虫雷达,一般昆虫的头尾方向的截面都落入Rayleigh散射区,雷达回波会因反射截面的减小而骤减,因而当昆虫具有定向时会在雷达屏幕上产生哑铃形回波[7,56]。毫米波雷达的波长为8mm,一般的迁飞蛾类(如黏虫、稻纵卷叶螟)不论身体的侧面还是正面对准雷达,都不会落入Rayleigh散射区,回波强度的变化不会差别很大,因此这类目标即使是有共同定向,在毫米波雷达上也不应表现出哑铃形。稻飞虱由于体长与毫米波雷达的波长相似,其头尾方向的截面落入Rayleigh散射区,理论上可产生哑铃形回波。有趣的是英国用多普勒雷达也观测到微小昆虫的共同定向,而且定向在6月份更偏北一些,在8月份更偏南一些,这与迁飞的季节性偏好方向一致[13]。因此几乎可以断定我国毫米波雷达观测到的哑铃形回波是稻飞虱产生的,即稻飞虱具有共同定向特性。
根据不同波长雷达探测到的信号差来确定回波的种类在气象雷达鉴定晴空回波的实践中曾多次使用[9,25]。根据雷达原理厘米波昆虫雷达“看”不到稻飞虱,而毫米波雷达却可以“看”到稻飞虱,如果同时使用厘米波和毫米波雷达进行观测,可以很容易地鉴定出哑铃形分布是稻飞虱产生的还是迁飞的蛾类产生的。当然空中捕捉能得到更直接的证据。
5.3 基站式和便携式谐波雷达的联合使用
基站式扫描谐波雷达非常适合在开阔地带研究昆虫的近距离飞行行为,尤其是觅食行为,在蜜蜂神经行为学、授粉昆虫生态学、气味介导的趋风性、最优搜寻策略和近距离扩散研究中发挥了重要作用[8]。但当昆虫飞入茂密的作物冠层或树林里,或者飞出监测范围,就很难再发现和追踪目标。由于谐波雷达不能区分不同的个体,研究时只能同时放飞1头或几头昆虫。尽管蜜蜂有“家”的概念,标记的蜜蜂也时有丢失,或者因为飞出监测范围长时间不归,观测者只能等待。这时如果继续释放新的标记昆虫,前面标记过的昆虫的意外归来会干扰试验,这种等待有时可持续半天或更长。便携式谐波雷达由于体积小重量轻、可以移动,更适合于寻找进入作物冠层活动的昆虫。由于两种雷达可以使用同样的标签,如果在农业昆虫研究中能将两种类型的雷达同时应用,即根据基站式雷达指示的方位使用便携式雷达到较远或茂密的作物丛中定位昆虫,也许会得到意想不到的结果。
5.4 现有观测数据的分析与新的挑战
昆虫定向与成层行为机理的研究在本世纪逐渐成为雷达昆虫学研究的热点,也取得了一些进展,但相互矛盾和不解之谜甚多。我国经过十年的持续观测积累了大量的夜间昆虫迁飞的资料,而且都有同时测到的高空风温场资料,这是一笔宝贵的资源。我们已初步建立了用风温场模拟昆虫成层的方法,现正在建立计算机分析程序,以期对历史资料进行更深入的分析,明确我国迁飞昆虫的成层机理。但昆虫学工作者缺乏编程经验,而程序员不懂昆虫,交叉学科人才不足给这项看似简单的工作平添几分困难。
越来越多的观测表明昆虫具有罗盘定向能力,但它们是使用什么罗盘还不清楚。使用雷达虽然可以准确观测昆虫的定向,但难以控制几百米高空的磁场、气压和风温等环境条件。红外三维轨迹捕捉及分析系统的尝试也未取得好的结果,飞行模拟器可以在模拟环境下测出昆虫的定向,但并不是所有的昆虫都适合吊飞[107-108]。最近小型昆虫的定向和成层也逐渐被认识[13,97],为昆虫的成层和定向机理研究提出了新的课题和挑战。
[1]程登发,封洪强,吴孔明.扫描昆虫雷达与昆虫迁飞监测[M].北京:科学出版社,2005.
[2]百度百科.http:∥baike.baidu.com,2011.
[3]维基百科.雷达[EB/OL].http:∥zh.wikipedia.org/zh-cn/%E9%9B%B7%E9%81%94,2011.
[4]翟保平.追踪天使—雷达昆虫学30年[J].昆虫学报,1999,42(3):315-326.
[5]翟保平.昆虫雷达让我们看到了什么?[J].昆虫知识,2005,42(2):217-226.
[6]封洪强.雷达昆虫学40年研究的回顾与展望[J].河南农业科学,2009(9):121-126.
[7]Riley J R.Riding on the wind:a radar perspective of insect flight[M].Inaugural Lecture Series,the University of Greenwich,London,1999:31.
[8]Chapman J W,Drake V A,Reynolds D R.Recent insights from radar studies of insect flight[J].Annu Rev Entomol,2011,56:337-356.
[9]Wilson J W,Weckwerth T M,Vivekanandan J,et al.Boundary layer clear-air radar echoes:origin of echoes and accuracy of derived winds[J].Journal of Atmospheric and Oceanic Technology,1994,11:1184-1206.
[10]Achtemeier G L.The use of insects as tracers for“clear-air”boundary-layer studies by Doppler radar[J].Journal of Atmospheric and Oceanic Technology,1991,8:746-765.
[11]Contreras R F,Frasier S J.High-resolution observations of insects in the atmospheric boundary layer[J].Journal of Atmospheric and Oceanic Technology,2008,25:2176-2187.
[12]Luke E P,Kollias P,Johnson K L,et al.A technique for the automatic detection of insect clutter in cloud radar[J].Journal of Atmospheric and Oceanic Technology,2008,25:1498-1513.
[13]Rennie S J,Illingworth A J,Dance S L,et al.The accuracy of Doppler radar wind retrievals using insects as targets[J].Meteorological Applications,2010,17(4):419-432.
[14]Rennie S J,Illingworth A J,Dance S L,et al.Utilization of Doppler radar wind measurements from insect returns[C]∥ERAD 2008-The Fifth European Conference on Radar in Meteorology and Hydrology,Helsinki,Finland,30June-4July 2008.Finnish Meteorological Institute:Helsinki,2008.
[15]Wood C R,O’Connor E J,Hurley R A,et al.Cloud-radar observations of insects in the UK convective boundary layer[J].Meteorological Applications,2009,16(4):491-500.
[16]Gauthreaux Jr S A,Livingston J W,Belser C G.Detection and discrimination of fauna in the aerosphere using Doppler weather surveillance radar[J].Integr Comp Biol,2008,48(1):12-23.
[17]Leskinen M.Observed polarimetric signals of insects[C]∥ERAD 2008-The Fifth European Conference on Radar in Meteorology and Hydrology,Helsinki,Finland,30June-4July 2008.Finnish Meteorological Institute:Helsinki,2008.
[18]Bachman S,Zrnic'D.Three-dimensional attributes of clear-air scatterers observed with the polarimetric weather radar[J].IEEE Geosciences and Remote Sensing Letters,2008,25:231-235.
[19]Mueller E A,Larkin R P.Insects observed using dual-polarization radar[J].Journal of Atmospheric and Oceanic Technology,1985,2:49-54.
[20]Zrnic D S,Ryzhkov A V.Observations of insects and birds with a polarimetric radar[J].IEEE Transactions on Geoscience and Remote Sensing,1998,36(2):661-668.
[21]Rennie S,Illingworth A,Dance S.On differentiating ground clutter and insect echoes from Doppler weather radars using archived data[J].Atmospheric Measurement Techniques Discussions,2010,3:1843-1860.
[22]Bachman S,Zrnic'D.Spectral density of polarimetric variables separating biological scatterers in the VAD display[J].Journal of Atmospheric and Oceanic Technology,2007,24:1186-1198.
[23]Geerts B,Miao Q.The use of millimeter Doppler radar echoes to estimate vertical air velocities in the fair-weather convective boundary layer[J].Journal of Atmospheric and Oceanic Technology,2005,22:225-246.
[24]Geerts B,Miao Q.Airborne radar observations of the flight behavior of small insects in the atmospheric convective boundary layer[J].Environ Entomol,2005,34:361-377.
[25]Martin W J,Shapiro A.Discrimination of bird and insect radar echoes in clear air using high-resolution radars[J].Journal of Atmospheric and Oceanic Technology,2007,24:1215-1230.
[26]Mueller E A.Differential reflectivity of birds and insects[C]∥Preprints,21st Conference on Radar Meteorology,Edmonton,AB,Canada:American Meteorological Society,1983:465-466.
[27]Schaefer G W.Radar observations of insect flight[C]∥Symposium of the Royal Entomology Society,London,1976,7:157-197.
[28]Drake V A.Automatically operating radars for monitoring insect pest migrations[J].Entomologia Sinica,2002,9(4):27-39.
[29]Chapman J W,Reynolds D R,Smith A D.Vertical-looking radar:A new tool for monitoring high-altitude insect migration[J].BioScience,2003,53(5):503-511.
[30]Chapman J W,Smith A D,Woiwod I P,et al.Development of vertical-looking radar technology for monitoring insect migration[J].Computers and Electronics in Agriculture,2002,35(2-3):95-110.
[31]翟保平.昆虫雷达:从研究型到实用型[J].遥感学报,2001,5(3):231-240.
[32]Smith A D,Reynolds D R,Riley J R.The use of vertical-looking radar to continuously monitor the insect fauna flying at altitude over southern England[J].Bull Entomol Res,2000,90(3):265-277.
[33]Smith A D,Riley J R,Gregory R D.A method for routine monitoring of the aerial migration of insects by using a verticallooking radar[J].Philosophical Transactions of the Royal Society of London.Series B:Biological Sciences,1993,340(1294):393-404.
[34]Drake V A,Harman I T,Wang H K.Insect monitoring radar:stationary-beam operating mode[J].Computers and Electronics in Agriculture,2002,35(2-3):111-137.
[35]Harman I T,Drake V A.Insect monitoring radar:analytical time-domain algorithm for retrieving trajectory and target parameters[J].Computers and Electronics in Agriculture,2004,43(1):23-41.
[36]Wang H K,Drake V A.Insect monitoring radar:retrieval of wingbeat information from conical-scan observation data[J].Computers and Electronics in Agriculture,2004,43(3):209-222.
[37]Dean T J,Drake V A.Monitoring insect migration with radar:the ventral-aspect polarization pattern and its potential for target identification[J].International Journal of Remote Sensing,2005,26(18):3957-3974.
[38]Reynolds D R,Chapman J W,Brooks S J,et al.Seasonal variation in the migration strategies of the green lacewingChrysoperlacarneaspecies complex[J].Ecological Entomology,2006,31(4):378-388.
[39]Chapman J W,Reynolds D R,Smith A D,et al.High-altitude migration of the diamondback mothPlutellaxylostellato the U.K.:a study using radar,aerial netting,and ground trapping[J].Ecological Entomology,2002,27(6):641-650.
[40]Drake V A,Wang H K,Harman I T.Insect monitoring radar:remote and network operation[J].Computers and Electronics in Agriculture,2002,35(2-3):77-94.
[41]Cheng D F,Wu K M,Tian Z,et al.Acquisition and analysis of migration data from the digitised display of a scanning entomological radar[J].Computers and Electronics in Agriculture,2002,35(2-3):63-75.
[42]程登发,吴孔明,田喆,等.扫描昆虫雷达实时数据采集、分析系统[J].植物保护,2004,30(2):41-46.
[43]Drake V A.Quantitative observation and analysis procedures for a manually operated entomological radar[R].CSIRO Australia Division of Entomology Technical Paper No.19,1981:1-41.
[44]Feng H Q,Wu K M,Cheng D F,et al.Radar observations of the autumn migration of the beet armywormSpodopteraexigua(Lepidoptera:Noctuidae)and other moths in northern China[J].Bull Entomol Res,2003,93(2):115-124.
[45]Feng H Q,Wu X F,Wu B,et al.Seasonal migration ofHelicoverpaarmigera(Lepidoptera:Noctuidae)over the Bohai Sea[J].J Econ Entomol,2009,102(1):95-104.
[46]Feng H Q,Wu K M,Cheng D F,et al.Northward migration ofHelicoverpaarmigera(Lepidoptera:Noctuidae)and other moths in early summer observed with radar in northern China[J].J Econ Entomol,2004,97(6):1874-1883.
[47]Feng H Q,Wu K M,Ni Y X,et al.Return migration ofHelicov-erpaarmigera(Lepidoptera:Noctuidae)during autumn in northern China[J].Bull Entomol Res,2005,95(4):361-370.
[48]Feng H Q,Zhao X C,Wu X F,et al.Autumn migration ofMythimnaseparata(Lepidoptera:Noctuidae)over the Bohai Sea in northern China[J].Environ Entomol,2008,37(3):774-781.
[49]Feng H,Wu K M,Cheng D F,et al.Spring migration and summer dispersal ofLoxostegesticticalis(Lepidoptera:Pyralidae)and other insects observed with radar in Northern China[J].Environ Entomol,2004,33(5):1253-1265.
[50]Feng H Q,Wu K M,Ni Y X,et al.High-altitude windborne transport ofHelicoverpaarmigera(Lepidoptera:Noctuidae)and other moths in mid-summer in Northern China[J].Journal of Insect Behavior,2005,18(3):335-349.
[51]Feng H Q,Wu K M,Ni Y X,et al.Nocturnal migration of dragonflies over the Bohai Sea in northern China[J].Ecological Entomology,2006,31(5):511-520.
[52]Feng H Q,Zhang Y H,Wu K M,et al.Nocturnal windborne migration of ground beetles,particularlyPseudoophonusgriseus(Coleoptera:Carabidae),in China[J].Agricultural and Forest Entomology,2007,9(2):103-113.
[53]吴孔明,翟保平,封洪强,等.华北北部地区二代棉铃虫成虫迁飞行为的雷达观测[J].植物保护学报,2006,33(2):163-167.
[54]吴孔明,程登发,徐广,等.华北地区昆虫秋季迁飞的雷达观测[J].生态学报,2001,21(11):1833-1838.
[55]Taylor P D,Brzustowski J M,Matkovich C,et al.radR:an open-source platform for acquiring and analysing data on biological targets observed by surveillance radar[J].BMC Ecol,2010,10:22.
[56]Riley J R.Collective orientation in night-flying insects[J].Nature,1975,253(5487):113-114.
[57]Riley J R,Reynolds D R.Orientation at night by high-flying insects[M]∥Danthanarayana W,ed.Insect Flight:Dispersal and Migration.Berlin:Springer-Verlag,1986:71-87.
[58]Reynolds D R,Riley J R.The flight behaviour and migration of insect pests:radar studies in developing countries[R].Chatham (United Kingdom):Natural Resources Institute.NRI Bulletin no.71,1997.
[59]Drake V A.Collective orientation by nocturnally migrating Australian plague locusts,Chortoicetesterminifera(Walker)(Orthoptera:Acrididae):a radar study[J].Bull Entomol Res,1983,73(04):679-692.
[60]Reynolds A M,Reynolds D R,Smith A D,et al.A single wind-mediated mechanism explains high-altitude‘non-goal oriented’headings and layering of nocturnally migrating insects[J].Proceedings of the Royal Society B:Biological Sciences,2010,277(1682):765-772.
[61]Chapman J W,Nesbit R L,Burgin L E,et al.Flight orientation behaviors promote optimal migration trajectories in highflying insects[J].Science,2010,327(5966):682-685.
[62]Chapman J W,Reynolds D R,Hill J K,et al.A seasonal switch in compass orientation in a high-flying migrant moth[J].Curr Biol,2008,18(19):R908-909.
[63]Chapman J W,Nesbit R L,Burgin L E,et al.Flight orientation behaviors promote optimal migration trajectories in highflying insects[J].Science,2010,327(5966):682-685.
[64]Chapman J W,Reynolds D R,Mouritsen H,et al.Wind selection and drift compensation optimize migratory pathways in a high-flying moth[J].Curr Biol,2008,18(7):514-518.
[65]Reynolds D R,Chapman J W,Edwards A S,et al.Radar studies of the vertical distribution of insects migrating over southern Britain:the influence of temperature inversions on nocturnal layer concentrations[J].Bull Entomol Res,2005,95(3):259-274.
[66]Wood C R,Chapman J W,Reynolds D R,et al.The influence of the atmospheric boundary layer on nocturnal layers of noctuids and other moths migrating over southern Britain[J].Int J Biometeorol,2006,50(4):193-204.
[67]Wood C R,Clarke S J,Barlow J F,et al.Layers of nocturnal insect migrants at high-altitude:the influence of atmospheric conditions on their formation[J].Agricultural and Forest Entomology,2010,12:113-121.
[68]Colpitts B G,Boiteau G.Harmonic radar transceiver design:miniature tags for insect tracking[J].IEEE Transactions on Antennas and Propagation,2004,52(11):2825-2832.
[69]桂连友,黄秀琴,李传仁.昆虫谐波雷达的发展和利用[J].应用昆虫学报,2011,48(3):732-738.
[70]Williams D W,Li G,Gao R.Tracking movements of individualAnoplophoraglabripennis(Coleoptera:Cerambycidae)adults:application of harmonic radar[J].Environmental Entomology,2004,33(3):644-649.
[71]Riley J R,Smith A D.Design considerations for an harmonic radar to investigate the flight of insects at low altitude[J].Computers and Electronics in Agriculture,2002,35(2-3):151-169.
[72]Boiteau G,Vincent C,Meloche F,et al.Harmonic radar:assessing the impact of tag weight on walking activity of Colorado potato beetle,Plumcurculio,and western corn rootworm[J].J Econ Entomol,2010,103(1):63-69.
[73]Riley J R,Smith A D,Reynolds D R,et al.Tracking bees with harmonic radar[J].Nature,1996,379:29-30
[74]O’Neal M E,Landis D A,Rothwell E,et al.Tracking insects with harmonic radar:a case study[J].American Entomologist,2004,50(4):212-218.
[75]Boiteau G,Colpitts B.The potential of portable harmonic radar technology for the tracking of beneficial insects[J].International Journal of Pest Management,2004,50(3):233-242.
[76]Mascanzoni D,Wallin H.The harmonic radar:a new method of tracing insects in the field[J].Ecological Entomology,1986,11(4):387-390.
[77]Wallin H,Ekbom B S.Movements of carabid beetles(Coleoptera:Carabidae)inhabiting cereal fields:a field tracing study[J].Oecologia,1988,77(1):39-43.
[78]Wallin H,Ekbom B.Influence of hunger level and prey densities on movement patterns in three species of pterostichus beetles(Coleoptera:Carabidae)[J].Environmental Entomology,1994,23(5):1171-1181.
[79]Charrier S,Petit S,Burel F.Movements ofAbaxparallelepipedus(Coleoptera,Carabidae)in woody habitats of a hedgerow network landscape:a radio-tracing study[J].Agriculture,Ecosystems & Environment,1997,61(2-3):133-144.
[80]Bliteau G,Colpitts B.Electronic tags for the tracking of insects in flight:effect of weight on flight performance of adult Colorado potato beetles[J].Entomologia Experimentalis et Applicata,2001,100:187-193.
[81]Boiteau G,Vincent C,Meloche F,et al.Harmonic radar:efficacy at detecting and recovering insects on agricultural host plants[J].Pest Manag Sci,2011,67(2):213-9.
[82]Boiteau G,Vincent C,Meloche F,et al.Evaluation of tag entanglement as a factor in harmonic radar studies of insect dispersal[J].Environ Entomol,2011,40(1):94-102.
[83]Gui L Y,Boiteau G,Colpitts B G,et al.Random movement pattern of fed and unfed adult Colorado potato beetles in bareground habitat[J].Agricultural and Forest Entomology,2011.In press.
[84]Gui L Y,Huang X Q,Li C R,et al.Validation of harmonic radar tags to study the movement of Chinese citrus fly[J].The Canadian Entomologist,2011:In press.
[85]Carreck N L,Osborn J L,Capaldi E A,et al.Tracking bees with radar[J].Bee World,1999,80(3):124-131.
[86]Menzel R,Fuchs J,Nadler L,et al.Dominance of the odometer over serial landmark learning in honeybee navigation[J].Naturwissenschaften,2010,97(8):763-767.
[87]Svensson G P,Valeur P G,Reynolds D R,et al.Mating disruption inAgrotissegetummonitored by harmonic radar[J].Entomologia Experimentalis et Applicata,2001,101(2):111-121.
[88]Reynolds A M,Reynolds D R,Smith A D,et al.Appetitive flight patterns of maleAgrotissegetummoths over landscape scales[J].J Theor Biol,2007,245(1):141-149.
[89]Cant E T,Smith A D,Reynolds D R,et al.Tracking butterfly flight paths across the landscape with harmonic radar[J].Proc Biol Sci,2005,272(1565):785-790.
[90]Ovaskainen O,Smith A D,Osborne J L,et al.Tracking butterfly movements with harmonic radar reveals an effect of population age on movement distance[J].Proc Natl Acad Sci U S A,2008,105(49):19090-19095.
[91]Wang R,Ovaskainen O,Cao Y D,et al.Dispersal in the Glanville fritillary butterfly in fragmented versus continuous landscapes:comparison between three methods[J].Ecological Entomology,2011,36:251-260.
[92]徐兴新,李栋.探地雷达探测堤坝白蚁巢研究[J].昆虫学报,1996,39(1):46-52.
[93]Henderson G.The termite menace in New Orleans:did they cause the floodwalls to tumble?[J].American Entomologist,2008,54(3):156-162.
[94]陈家银,李富长,司马军.白龟山水库白蚁隐患的探地雷达检测试验[J].河南水利与南水北调,2007,2007(1):54-55.
[95]Yang X H,Henderson G,Mao L X,et al.Application of ground penetrating radar in detecting the hazards and risks of termites and ants in soil levees[J].Environ Entomol,2009,38(4):1241-1249.
[96]Drake V A,Reynolds D R.Radar entomology:observing insect flight and migration[M].CABI,2011:304.
[97]齐会会.稻飞虱的雷达监测与迁飞轨迹分析[D].北京:中国农业科学院,2011.
[98]齐会会,张云慧,程登发,等.褐飞虱2009年秋季回迁的雷达监测及轨迹分析[J].昆虫学报,2010,53(11):1256-1264.
[99]杨秀丽,陈林,程登发,等.毫米波扫描昆虫雷达空中昆虫监测的初步应用[J].植物保护,2008,34(2):31-36.
[100]高月波,陈晓,陈钟荣,等.稻纵卷叶螟(Cnaphalocrocismedinalis)迁飞的多普勒昆虫雷达观测及动态[J].生态学报,2008,28(11):5238-5247.
[101]张云慧,陈林,程登发,等.草地螟2007年越冬代成虫迁飞行为研究与虫源分析[J].昆虫学报,2008,51(7):720-727.
[102]张云慧,乔红波,程登发,等.垂直监测昆虫雷达空中昆虫监测的初步应用[J].植物保护,2007,33(3):23-26.
[103]Zhang P F,Ryzhkov A,Zrnic D.Polarimetric prototype of the WSR-88Dradar observation of birds and insects[C]∥Preprints CD,12th Conference on Aviation,Range,and Aerospace Meteorology,Atlanta,GA,28January-3February 2006.American Meteorological Society:Boston,2006.
[104]Leskinen M,Markkula I,Koistinen J,et al.Pest insect immigration warning by an atmospheric dispersion model,weather radars and traps[J].Journal of Applied Entomology,2010,135(1-2):55-67.
[105]Nieminen M,Leskinen M,Helenius J.Doppler radar detection of exceptional mass-migration of aphids into Finland[J].Int J Biometeorol,2000,44(4):172-181.
[106]Markkula I,Ojanen H,Tiilikkala K,et al.Insect migration case studies by polarimetric radar[C].In ERAD 2008-The Fifth European Conference on Radar in Meteorology and Hydrology,Helsinki,Finland,30June-4July 2008.Finnish Meteorological Institute:Helsinki,2008.
[107]吴博,封洪强,赵奎军,等.视频轨迹分析技术在昆虫行为学研究中的应用[J].植物保护,2009,35(2):1-6.
[108]施翔宇,封洪强,刘中芳,等.实验条件下黏虫、棉铃虫和小地老虎的定向行为比较[J].植物保护,2010,36(4):60-63.
Applications of radar in entomological research
Feng Hongqiang
(HenanKeyLaboratoryofCropPestControl,InstituteofPlantProtection,HenanAcademyofAgriculturalSciences,Zhengzhou450002,China)
Insect radar has been used as an excellent tool in entomological research,such as flight behavior of insects during high-altitude and long-distance migration or just above ground,because of its fascinating ability in detecting flight behavior of insects in natural free flying from a long distance with little disturbance to them.Weather radar has increasingly been used to monitor birds and insects,and GPR (Ground Penetrating Radar)has also been used to detect nests of termites and ants in soil levees.This review evaluated the capability of different types of radars and their performance in entomological research,to tell what the radar can do and the challenges in using radar from the view of an entomologist.
radar; insect; migration; target identification; application
S 431.9
A
10.3969/j.issn.0529-1542.2011.05.001
2011-09-05
国家自然科学基金项目(30971873);河南省杰出青年基金项目(094100510024);河南省重大公益项目(081100911300)
* 通信作者E-mail:feng_hq@163.com
猜你喜欢
玩具世界(2022年3期)2022-09-20 01:48:20
林业科学(2021年4期)2021-05-21 08:10:22
甘肃教育(2020年2期)2020-09-11 08:01:48
阅读(快乐英语高年级)(2020年3期)2020-07-16 03:41:38
小学生优秀作文(低年级)(2018年9期)2018-09-10 05:47:38
长春师范大学学报(2018年8期)2018-08-17 12:40:20
小学生必读(低年级版)(2018年11期)2018-03-13 00:54:24
小学生导刊(2017年13期)2017-06-15 20:29:38
中国学术期刊文摘(2015年16期)2015-11-01 01:34:22
中国学术期刊文摘(2015年24期)2015-02-28 01:39:26
