
细粒棘球蚴重组抗原rEgferritin诱导宿主抗感染免疫应答的初步研究*
2011-08-21 10:23王娅娜王淑静
中国人兽共患病学报 2011年10期
王娅娜,王 洁,王淑静,张 焱,赵 巍
2.宁夏医科大学医学遗传学与细胞生物学教研室;
3.宁夏回族自治区遗传与生殖重点实验室
细粒棘球蚴重组抗原rEgferritin诱导宿主抗感染免疫应答的初步研究*
王娅娜1,2,3,王 洁1,王淑静1,张 焱1,赵 巍1,2,3
目的 初步探讨细粒棘球蚴重组铁蛋白rEgferritin免疫小鼠产生的抗细粒棘球蚴感染的免疫机制。方法 rEgferritin免疫ICR小鼠3次、同时设立佐剂组和对照组,第12w用细粒棘球蚴原头蚴对3组小鼠进行攻击感染,5个月后剖杀小鼠,检查各组小鼠腹腔内包囊个数,根据公式计算保护力。分别于0w,12w,32w取血,制备血清,同时取脾,制备淋巴细胞悬液。通过ELISA检测血清中的IgG及其抗体亚型、IgE及MTT;通过流式细胞仪检测脾淋巴细胞中CD+4及CD+8亚群百分比。结果 rEgferritin组小鼠血清中的IgG,IgG2a,IgG2b持续增高,与佐剂组相比具有显著性差异(P<0.01);3组IgG3无差异。rEgferritin组免疫小鼠可诱导小鼠产生85.6%的保护力抵御细粒棘球蚴原头蚴的再次感染,与对照组相比具有显著性差异(P<0.01);rEgferritin组和佐剂组CD+4T细胞和CD+8T细胞均增加,rEgferritin组CD+4T细胞增加更为明显,与佐剂组相比有显著性差异(P<0.01)。rEgferritin组脾细胞在受到rEgferritin抗原和天然抗原刺激后,细胞增殖明显。结论 细粒棘球蚴重组抗原rEgferritin通过诱导宿主产生体液免疫级细胞免疫应答的方式抵御细粒棘球蚴感染的,说明rEgferritin是具有研究价值和潜力的包虫病候选疫苗分子。
细粒棘球蚴;铁蛋白;免疫应答
2.宁夏医科大学医学遗传学与细胞生物学教研室;
3.宁夏回族自治区遗传与生殖重点实验室
细粒棘球蚴病又称包虫病,是一种人兽共患寄生虫病。它是导致各国尤其是发展中国家[1]的致残率和致死率较高的原因之一。许多国家已经实施对细粒棘球蚴虫终宿主病狗及其粪便的处理等防控措施,在一定程度上减少了细粒棘球蚴的虫卵从狗向人的传播,可是包虫病在世界某些地区的发病率仍然高发甚至有回升趋势[2]。近年来,用疫苗防控寄生虫病成为研究的热点。铁蛋白作为寄生虫疫苗的候选基因受到了越来越多的关注。曼氏血吸虫[3],日本血吸虫[4-5],牛带绦虫[6],细粒棘球绦虫[7]及细粒棘球蚴中国株的铁蛋白基因已相继被克隆[8],本课题组前期已经构建了细粒棘球蚴重组表达质粒Eg.ferritin/pET-28a,并纯化出重组铁蛋白[9]。本实验在此基础上,通过体内试验,初步研究细粒棘球蚴铁蛋白作为抗原免疫小鼠后进行细粒棘球蚴攻击感染实验,观察小鼠的相关体液免疫和细胞免疫指标。
1.1 材料
1.1.1 实验动物 6~8周龄雄性ICR小鼠,体重(18±2)g,共30只,购自宁夏医科大学实验动物中心。
1.1.2 细粒棘球蚴原头蚴 从宁夏医科大学第一附属医院肝胆外科包虫病患者手术摘除的完整包囊中无菌条件下抽取囊液,分离原头蚴,经PBS(pH 7.2)洗涤3次后,弃去上清液,用0.5%伊红染色计数原头蚴的着色率及形态,原头蚴的存活率>90%,用PBS配成1.5×104个原头蚴/mL悬液,用于攻击感染ICR小鼠。
1.1.3 细粒棘球重组表达菌株Eg.ferritin/pET-28a/BL21plysS由本室提供。
1.1.4 主要试剂 辣根过氧化物酶(HRP)标记的羊抗鼠IgG、IgG1、IgG2a、IgG2bIgG3、IgE及 ConA(刀豆蛋白A)、MTT购自华美生物工程公司北京分公司。RPMI 1640培养基为GIBiCO公司产品;PE标记的大鼠抗小鼠CD+8T以及FITC标记的大鼠抗小鼠CD+3T细胞亚群单克隆抗体为BD Biosciences产品,购自北京利文商贸有限责任公司。
1.2 方法
1.2.1 动物分组免疫及攻击感染 将ICR小鼠随机分为A、B、C 3组,每组15只。A组为rEgferrtin免疫组;B组佐剂组;C组为对照组:不免疫攻击感染组。A组:免疫注射50μg rEgferritin和等量弗氏佐剂的乳化剂,B组:注射与A组等量弗氏佐剂与PBS的乳化剂,第1次免疫为基础免疫注射弗氏完全佐剂,以后2次为加强免疫,注射弗氏不完全佐剂。两组小鼠在0、2、4w各免疫1次,共3次,均采用背部皮下多点注射免疫,每次注射量为100μL/只。12w时,A、B、C组小鼠各用1500枚/只活棘球蚴原头蚴腹腔内注射进行攻击感染。分别于免疫前(0w)攻击感染 (12w)和剖杀(32w)小鼠眼眶采血,将血液4 000r/min离心10min,吸取血清,-85℃保存。
1.2.2 各组小鼠免疫保护力的计算 攻击感染后20w(即32w)剖杀小鼠,打开小鼠腹腔,对各组小鼠腹腔内包囊个数进行计数并测量包囊直径,根据公式计算出各组小鼠的保护力,免疫保护力(%)=(1-实验组平均包囊数/对照组平均包囊数)× 100%[10]。
1.2.3 ELISA检测IgG及其亚型和IgE抗体 用终浓度为10μg/mL rEgferritin—碳酸缓冲液(pH 9.6)包被96孔酶标板,100μL/孔,4℃过夜;次日用PBS洗板3次,5%脱脂奶粉37℃封闭2h;加1∶100稀释的A、B、C组鼠血清,100μL/孔,37℃反应2h,PBS洗板3次;加入1∶1 000稀释的羊抗鼠Ig-HRP(IgG1,IgG2a,IgG2b,IgG3及IgE)100μL/孔,37℃反应2h,PBS洗板3次;加四甲基联苯胺(TMB 40mg/mL)底物溶液,15min后用2mol/L H2SO4(50μL/孔)终止反应,用酶标分析仪测定OD490nm每孔的吸光值。
1.2.4 脾细胞悬液制备 剖杀A、B、C组小鼠,置75%乙醇中消毒3min;无菌取脾,匀浆,用2mL PBS冲洗,200目尼龙膜过滤,滤液置10mL离心管;加5~10倍的红细胞裂解液来溶解红细胞,离心5min,2 000r/min;去上清,用PBS洗2次,离心5min,2 000r/min;去上清,加2mL PBS,计数后用含10%BSA的RPMI1640培养液调整细胞浓度为5×106/mL;台盼蓝染色后发现脾细胞存活率>95%。
1.2.4.1 脾细胞CD+4T、CD+8T亚群测定 将上述制备好的脾细胞悬液5×106/mL,取200μL,每份标本分两组,4℃离心去上清,先用含FITC标记的CD+3单抗进行标记,然后用PE标记的CD+8标记,静置30min后,再次离心,以2mL冷PBS缓冲液(含小牛血清及NaN3)洗2次,加入1%多聚甲醛0.5mL,4℃保存。流式细胞仪检测脾CD+4、CD+8淋巴细胞亚群的百分比,用B-D流式细胞仪523nm波长下测定细胞荧光,每次测定1×105个细胞,分析软件为Mod Fit LT。
1.2.4.2 T淋巴细胞增殖试验(MTT法) 将A、B、C各组小鼠的脾细胞悬液5×106/mL,加入96孔细胞培养板,分别用培养液、ConA、重组抗原(rEgferritin)及天然抗原(原头蚴匀浆后制备的上清)刺激后检测T淋巴细胞活性,刺激物的工作浓度皆为5μg/mL,每份标本分无刺激、ConA、rEgferritin和天然抗原4小组,均重复3孔,置37℃5%CO2条件下培养72h,培养结束前4h,加入20μL MTT(5mg/mL),继续培养4h,结束后2 000r/min,离心10min,弃100μL上清,加100μL二甲基亚砜(DMSO)振荡,轻轻反复吹打至甲簪(formazane)充分溶解,测定吸光度(OD490nm)。
1.2.5 统计学处理 采用SPSS 11.5软件进行分析,组间比较用单因素方差分析(ANOVA)。
2 结 果
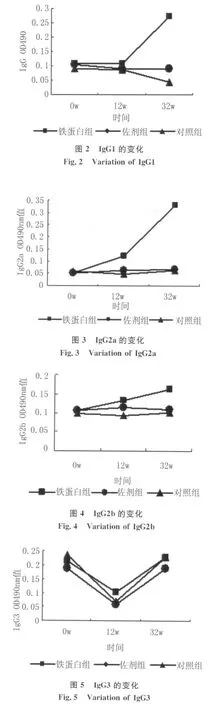
2.1 各组血清中IgG1,IgG2a,IgG2b,IgG3及IgE的测定 铁蛋白组IgG随时间的延长一直上升,并且在攻击感染后(12w)继续上升,与佐剂和对照组相比有显著差异,佐剂组IgG和对照组在攻击感染后(12w)有所上升,见图1。铁蛋白组IgG1攻击感染后明显上升,与佐剂组和对照组有显著差异,佐剂组随时间无明显变化,对照组IgG1在剖杀前(32w)与攻击前(12w)和免疫前(0w)相比有所下降,与佐剂组相比有差异,见图2。铁蛋白组IgG2a随时间的延长不断增加,与佐剂组和对照组相比有显著差异,佐剂组与对照组之间无明显差异,见图3。铁蛋白组IgG2b随免疫次数的增加而增加,与佐剂组及对照组相比有显著性差异。佐剂组IgG2b12w稍微增高,之后下降为正常,与对照组之间无差异,见图4。铁蛋白组、佐剂组和对照组IgG3均随时间下降,至在12w达到最低,之后上升为免疫前水平,3组之间相比无统计学差异,见图5。铁蛋白组及佐剂组的IgE值持续上升,2组之间无差异,与对照组相比有显著差异,而对照组在12w后有所上升,见图6。
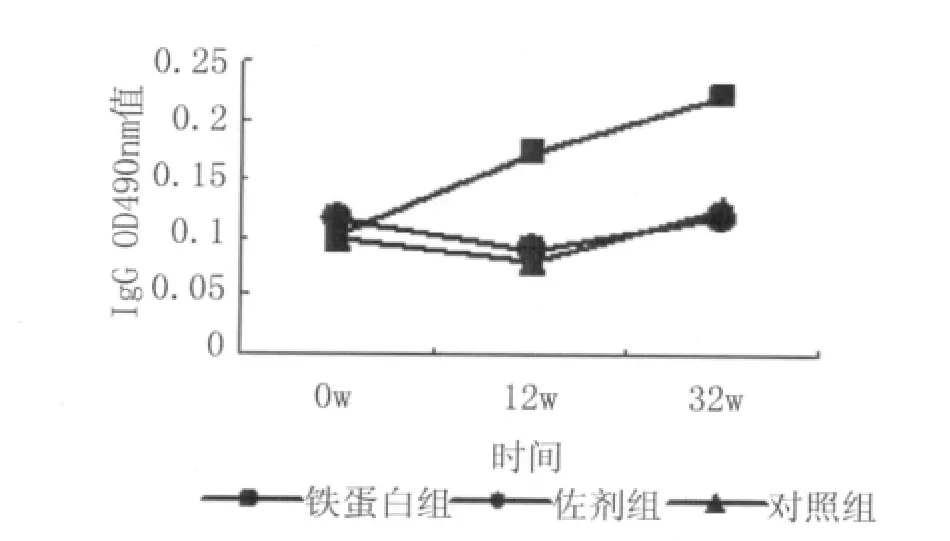
图1 IgG的变化Fig.1 Variation of IgG
图6 IgE的变化Fig.6 Variation of IgE
2.2 免疫保护力的测定 3组小鼠腹腔内注射细粒棘球蚴原头蚴1500枚/只,除对照组1只小鼠攻击感染后死亡,其余3组小鼠,5个月后(32w)剖杀,检查小鼠腹腔内各个器官,计数包囊个数及直径。发现铁蛋白免疫组小鼠体内的包囊数及包囊大小与佐剂组和对照组相比明显少,并具有显著性差异(P<0.01)。根据公式计算出相对于对照组,铁蛋白可诱导小鼠产生85.6%的免疫保护力抵御攻击感染,佐剂组可产生56.3%,见表1。
2.3 脾细胞中CD+4T及CD+8T细胞亚群的测定原头蚴攻击感染后20周,铁蛋白组的脾淋巴细胞CD+4T和CD+8T均有增高,CD+4T与佐剂组和对照组相比明显增加,有显著性差异(P<0.01),CD+8T与佐剂组的之间差异显著(P<0.05),与对照组相比无明显差异。佐剂组与对照组相比无显著差异(P>0.05),见表2。

表1 各组小鼠产生的包囊数及免疫保护力Table 1 Number of hydatid cysts and protective immunity in three groups

表2 原头蚴攻击感染后各组小鼠脾淋巴细胞亚群的检测Table 2 Detection on subsets of spenocytes of three group mice after challenged with Egprotoscoleces
2.4 各组脾细胞增殖实验rEgferritin抗原 3组小鼠脾淋巴细胞与四种物质共同培养,发现用ConA刺激可使3组脾淋巴细胞增殖能力均提高,与同组无刺激物相比具有统计学差异,但3组之间无统计学差异(图7)。用rEgferritin抗原刺激后,铁蛋白组脾淋巴细胞与其它两组相比明显增加(P<0.05),与同组无刺激物和conA刺激相比具有显著性差异,佐剂组与对照组之间无明显差异。用天然抗原原头蚴刺激后,铁蛋白组脾淋巴细胞增加更为明显与其它两组相比具有显著性差异(P<0.01),与同组无刺激物和conA刺激相比具有显著性差异,佐剂组与对照组之间无明显差异。提示铁蛋白组的脾淋巴细胞在体外能被rEgferritin蛋白诱导产生较好的免疫记忆能力,被特异性的刺激增生。
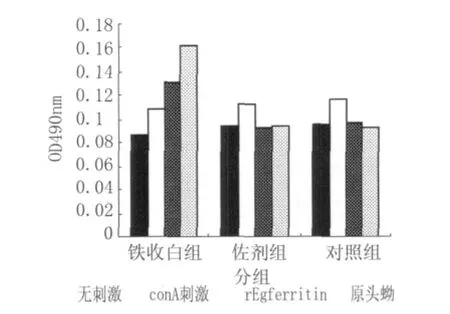
图7 3组小鼠淋巴细胞的增殖Fig.7 Three groups of lymphocytes cell growth with different stimulation
3 讨 论
本实验室前期工作已经显示,细粒棘球蚴重组铁蛋白具有较好的抗原性及免疫原性,诱导小鼠可产生较高的免疫能力抵抗细粒棘球蚴原头蚴的感染。但是对于重组铁蛋白诱导小鼠产生怎样的免疫机制抵抗感染目前未见相关报道。本次实验重组铁蛋白诱导小鼠产生的免疫机制进行初步研究。
目前一些研究表明在抵抗细粒棘球蚴感染中体液免疫和细胞免疫均发挥重要的作用[11-12]。在2次Eg感染小鼠的实验中发现,感染前3d内,腹腔注射的PSC周围很快被细胞浸润。早期血清中可检测到微量的IgG1,IgG2a和IgG3。后期血清中的IgG1,IgG2a和IgG3达到高水平[13]。感染肠道寄生虫的宿主血清中通常都能检测到高水平的IgE[14-15]。IgE依赖肥大细胞,聚集嗜酸性粒细胞至寄生虫周围,增强嗜酸性粒细胞抗寄生虫的功能。本实验关于小鼠血清IgG及其亚型和IgE的检测显示:铁蛋白组IgG在免疫后及攻击感染后持续增加,佐剂组在攻击感染后IgG增加这与我们前期实验[9]和Liu[16]等观察的细粒棘球蚴感染小鼠的结果相似。铁蛋白组IgG1在攻击感染后增加明显,而对照组IgG1在攻击感染后下降,这提示铁蛋白诱导小鼠产生高的IgG1具有抵抗细粒棘球蚴生长的作用,对照组IgG1下降,有利于寄生虫的生长。铁蛋白组IgG2a,IgG2b在免疫后和攻击感染后持续增加,与佐剂组和对照组相比均具有显著性差异,说明IgG1,IgG2a,IgG2b在抗寄生虫感染中发挥着重要的作用。3组IgG3变化一致,无明显变化,IgG3参与免疫逃逸机制。铁蛋白组及佐剂组的IgE值持续上升,两组之间无统计学差异,与对照组相比有显著性差异,而对照组在12w后有所上升,这些可能与IgE参与致敏反应有关。
淋巴细胞包括两个主要亚群:CD+4+和CD+8T细胞,在寄生虫感染中,两种细胞都可参与宿主保护力和病理变化的形式。Riley等[17]发现感染Eg原头蚴的小鼠淋巴结CD+4T细胞亚群降低,CD+8T细胞亚群升高。本研究中rEgferritin蛋白免疫小鼠,原头蚴攻击感染后20周CD+4T细胞与佐剂组和对照组相比显著增高(P<0.01),CD8+T细胞与佐剂组相比显著升高(P<0.05),但佐剂组和对照组相比较无显著性差异(P>0.05)。推测该重组抗原刺激CD+4T细胞可能向Th1细胞分泌,介导细胞免疫应答,还可能激活巨噬细胞和NK细胞,增加它们杀伤细粒棘球蚴的作用。同时CD+8T的杀伤性T细胞也被抗原激活,形成细胞毒性T(Tc)淋巴细胞,可能通过细胞裂解和细胞凋亡杀伤细粒棘球蚴。佐剂组的小鼠在佐剂的刺激下产生了一定的非特异性的抗体,可在感染早期,刺激机体产生高的CD+4T杀伤细粒棘球蚴,形成保护性免疫,细粒棘球蚴增长缓慢,感染后期以CD+4T下降,使得细粒棘球蚴出现明显增殖,宿主体内感染严重。这与Sakamoto等[18]报道基本一致。
淋巴细胞增殖实验是反映细胞免疫应答的传统方法之一,MTT法检测证实rEgferritin免疫组的小鼠脾淋巴细胞,用rEgferritin和天然抗原刺激培养后,脾淋巴细胞的数目远高于原液和conA刺激的值,具有显著差异(P<0.05)。推测rEgferritin可诱导小鼠脾细胞产生较好的记忆能力。而且用天然抗原刺激与rEgferritin刺激相比也有显著差异(P<0.05)。考虑由于rEgferritin是包涵体,在纯化的过程中,蛋白的天然构象发生了一些改变。所以天然抗原比重组抗原更容易刺激细胞的增殖。
综上所述,寄生虫感染的免疫机制十分复杂,其中包括多种免疫细胞和细胞因子的参与,它们互相作用,互相制约形成调节网络。研究寄生虫感染的免疫机制及各类淋巴细胞的作用有助于制定有效的防治措施,对研制寄生虫病疫苗更具有重要意义。但由于寄生虫抗原的复杂性及免疫系统调节的精密性,各类淋巴细胞在各种寄生虫感染中的免疫作用及其相互间的调节机制还不清楚,尚需进一步研究。通过本实验初步证明rEgferritin可诱导小鼠产生85.6%的免疫保护力,通过体液免疫及细胞免疫应答方式抵御细粒棘球蚴感染的,是具有研究价值和潜力的包虫病候选疫苗分子。下一步将对其免疫机制进行详细研究,为寄生虫免疫防御提供一定的理论依据。
[1]Achussler P,Potters E,Winnen T,et al.An isoform of ferritin as a component of protein yolk platelets inSchistosomamansoni[J].Molecular Reproduction and Development,1995,41(3):325-330.
[2]Lightowlers M W,Flisser A,Gauci C G,et al.Vaccination against cysticercosis and hydatid disease[J].Parasitology Today,2000,16(5):191-196.
[3]Dietzal J,Hirzmann J,Preis D,et al.Ferritins ofSchistosoma mansoni;sequence comparison and expression in female and male worms[J].Mol Biochem Parasitol,1992,50:245-254.
[4]王敏,易新元,曾宪芳,等.日本血吸虫铁蛋白基因的克隆,表达及其免疫诊断的研究[J].中国免疫学杂志,2004,20:696-699.
[5]舒新华,易新元,张顺科,等.日本血吸虫重组铁蛋白表达和疗效考核价值[J].中国寄生虫病防治杂志,2000,2(13):134-138.
[6]Benitex L,Harrison Leslie J S,Parkhouse R,et al.Sequence and immunogenicity of Taenia saginate ferritin[J].Mol Biochem Parasitol,1996,82:113-116.
[7]Eresfeld K,Craig PS.Cloning and immunological characterization ofEchinococcusgranulosusferritin[J].Parasite Res,1995,81:382-387.
[8]王娅娜,赵嘉庆,丁淑琴,等.细粒棘球蚴中国大陆株铁蛋白基因的分子克隆和序列分析[J].宁夏医学院学报,2005,28:253-257.
[9]王娅娜,丁淑琴,王健,等.细粒棘球蚴铁蛋白基因的克隆重组高效表达及免疫学初步研究[J].中国人兽共患病学报,2006,22(5):399-402.
[10]Dempster RP,Harrison GB,Berridge MV.Maternal transfer of protection fromEchinococcusgranulosusinfection in sheep[J].Research in Veterinary Science,1995,58,197-202.
[11]Barriga OO,Al-Khalidi NW.Humoral immunity in the prepatent primary infection of dogs withEchinococcusgranulosus[J].Veterinary Immunology and Immunopathology,1986,11(4):375-389.
[12]Haralabidis S,Karagouni E,Frydas S,et al.Immunoglobulin and cytokine profile in murine secondary hydatidosis[J].Parasite Immunology,1995,17(12):625-630.
[13]Pater C,Muller V,Harraga S,et al.Intestinal and systemic humoral immunological events in the susceptible Balb/c mouse strain after oral administration ofEchinococcusmultiloculariseggs[J].Parasite Immunology,1998,20(12):623-629.
[14]Bell RG.IgE,allergies and helminth parasites:a new perspective on an old conundrum[J].Immunology and Cell Biology,1996,74(4):337-345.
[15]Capron A,Dessaint J P.Immunologic aspects of schistosomiasis[J].Annual Review of Medicine,1992,43:209-218.
[16]Liu D,Lightowlers MW,Rickard MD.Examination of murine antibody response to secondary hydatidosis using E LISA and immunoelectrophoresis[J].Parasite Immunol,1992 ,14 :239-248.
[17]Riley EM,Dixon JB.ExperimentalEchinococcusgranulosusinfection in mice:immunocytochemical analysisi of lymphocyte populations in local lymphokd infections during early infection[J].Parasitol,1987,94(3):523-532.
[18]Sakamoto T,Cabrera PA.immunohistochemical observations on cellular response in unilocular hydatid lesions and lymph modes of cattle[J].Acta Trop,2003,85(2):271-279.
An initial investigation on effects of rEgferritin on immune response of mice infected with hydatid cysts ofEchinococcusgranulosus
WANG Ya-na,WANG Jie,WANG Shu-jing,ZHANG Yan,ZHAO Wei
(CenterofScientificTechnologyofNingxiaMedicalUniversity,Yinchuan750004,China)
In order to initially investigate the effects of rEgferritin on the immune response of mice infected with hydatid cysts ofEchinococcusgranulosus,ICR mice were immunized with rEgferritin by thrice and challenged by protoscoleces ofE.granulosusat 12week.Mice were killed at 20weeks after infection,and blood of mice were obtain at 0week,12week and 32 week prepare serum,The splenocytes were isolated and the percentages of the splenic CD+4and CD+8T lymphocytes were measured by FCM .In every groups splenocytes were cultivated with stimulation of rEgferritin native antigen,ConA and normal nutrient.The levels of the supernatant of cultures were determined by ELISA.The adjuvant group and blank group were set up for comparison.It was found that the OD values of IgG,IgG2aand IgG2bof immunized group were all higher than that of the control group(P<0.01),and the OD value of IgG3was no significant difference among the three groups.Mice vaccinated with rEgferritin and challenged intraperitoneally withE.granulosusprotoscoleces revealed significant protective efficacy up to 85.6%.The percentage of the splenic CD+4and CD+8T lymphocytes were all increased in the immunized group of mice,but CD+4T lymphocytes of immunized group were obviously higher than adjuvant group and control group(P<0.01),splenic lymphocytes cell of immunized rEgferritin group of mice were growthed obviously by induced with rEgferritin protein and native antigen ofEchinococcusgranulosus.These data indicated that rEgferritin protein immunized could can induce humoral immunity of mice against the hydatid cyst ofE.granulosus.The rEgferritin might be a valuable candidate vaccine for against hydatid disease.
Echinococcusgranulosus;rEgferritin;immune response
R383.3
A
1002-2694(2011)10-0882-05
*国家自然科学基金(30660167),宁夏回族自治区自然科学基金(NZ0993)
赵巍,Email:zw-6915@163.com
1.宁夏医科大学医学科学技术研究中心,银川 750004;
2011-05-04;
2011-07-30
猜你喜欢
有色金属(矿山部分)(2021年4期)2021-08-30
昆明医科大学学报(2021年3期)2021-07-22
中国海上油气(2020年5期)2020-10-20
中国乳品工业(2019年10期)2019-12-09
中成药(2018年10期)2018-10-26
中成药(2018年6期)2018-07-11
中国乳业(2018年8期)2018-01-25
中成药(2017年8期)2017-11-22
家庭用药(2016年7期)2016-05-14
中国资源综合利用(2016年9期)2016-01-22
