
泰索帝对甲状腺未分化癌细胞系作用的蛋白质组学研究
2011-08-20 10:39付言涛郑海波赵吉生
中国实验诊断学 2011年9期
付言涛,孙 辉,郑海波,赵吉生
(1.吉林大学中日联谊医院甲状腺外科,吉林长春 130033;2.吉林大学第二临床学院麻醉科)
应用蛋白质组学技术,可从整体上定性和定量检测分析甲状腺肿瘤细胞经化疗药物作用前后细胞内蛋白质分子的变化,在蛋白水平进一步探讨药物作用的机制及寻找新药靶点有重要的意义。泰索帝是一种紫杉类抗肿瘤药物,临床研究提示它能够改善甲状腺未分化癌的预后,但尚无从整体蛋白质水平上对其作用的机制进行相关研究。
本实验拟从蛋白质组学水平观察泰索帝作用甲状腺未分化癌细胞前后蛋白表达谱的变化,探讨泰索帝对人甲状腺未分化癌细胞的作用机制,从蛋白质组学角度进一步深化对泰索帝抗肿瘤作用机制的认识。
1 材料与方法
1.1 材料
1.1.1 细胞及试剂 人甲状腺未分化癌细胞系(TA-K)由日本东北大学医学部第二外科惠赠;变性剂UREA(尿素),Thiourea(硫脲),表面活性剂CHAPS,载体两性电解质ampholyte,还原剂TBP(三丁基膦),还原剂DTT(二硫苏糖醇),Iodoacetamide(碘乙酰胺),SDS(十二烷基磺酸钠),Arc(丙烯酰胺),PDA(哌嗪双丙烯酰胺),TEMED(催化剂),APS(过硫酸胺),矿物油,低熔点琼脂糖,ReadyStripTM IPG胶条(17 cm)均购自美国Sigma-Aldrich公司;Bio-Rad AC DC蛋白定量试剂盒购自BIO-RAD公司;IPG胶条购自BIO-RAD公司;泰索帝购自法国安万特公司。
1.1.2 主要仪器有PROTEAN IEF系统组件(BIORAD公司);PROTEANⅡxi系统组件(BIO-RAD公司);4700TOF-TOF质谱仪(ABI公司);GS-800校准型光密度扫描成像系统(BIO-RAD公司)等。
1.1.3 主要溶液有细胞培养液(WE,10%胎牛牛血清);细胞样品裂解液(7 M UREA,2 M thiourea,4%w/v CHAPS,40 mM Tris碱,与1%v/v蛋白酶抑制剂的混合液);水化液(8 M urea,2%w/v CHAPS,20 mM DTT,0.2%v/v Bio-lyte 3-10,0.002%痕量溴酚蓝)。平衡缓冲母液(50 mM Tris-HCl,pH 8.8,6 M urea,30%甘油,2%SDS);12%SDS-PAGE凝胶缓冲液(12%Arc,0.38 mmol/L Tris(pH 8.8),0.1%SDS,0.08%APS,0.04%TEMED)。
1.2 蛋白质组学研究
1.2.1 样品制备 当贴壁生长于WE培养液中的TA-K细胞处于生长对数期时,将泰索帝以0.01 μ g/mL的浓度加入细胞培养液,共同培养72 h.对溶媒组加入等体积的0.002 6%乙醇浓度溶液。胰蛋白酶消化细胞,收集,磷酸缓冲液(PBS)洗涤、离心(1 500×g,10 min)3次。加入裂解缓冲液(1.5×106个细胞大约加入 100 μ L裂解液),吹打混匀.液氮中反复冻融3次,在室温下融化。加 50 μ g/ml RNase及200 μ g/ml Dnase,4℃放置 15 min。经高速离心 15 000转,15 min,取上清,采用Bio-Rad RC DC蛋白定量试剂盒进行蛋白定量。分装至 Eppendof管里保存在-80℃备用
1.2.2 胶条溶胀与水化 17 cm水化盘中加入含有100 μ g蛋白的 400 μ L水化液,将IPG 胶条覆盖到液面上,水化盘置于PROTEAN IEF电泳仪中,被动水化,20℃过夜(>12 h).
1.2.3 第一向等电聚焦 将充分溶胀的 IPG胶条取出,置于聚焦盘的相应聚焦槽中,再将聚焦盘置于PROTEAN IEF电泳仪中,20℃、60 kVh进行等电聚焦.
1.2.4 第二向SDS-PAGE取出聚焦好的IPG胶条,经含有1%DDT和5%碘乙酰胺的平衡缓冲液平衡30分钟,以还原蛋白巯基,移至预先灌注好的12%SDS-PAGE凝胶顶端,与胶面完全接触,将凝胶移至垂直电泳槽内.起始电流为10 mA/gel,30分钟后,加大电流至24 mA/gel,待指示剂达到底部边缘时停止电泳.取出凝胶,切角做记号.
1.2.5 银染 程序设置如下
1.2.6 图像扫描与分析 凝胶经胶体银染色后,使用PowerLook 2100XL型光密度扫描仪和PHOTOSHOP CS软件对凝胶进行扫描成像。在PDQuest双向电泳分析软件的辅助下,对图像进行背景消减、斑点检测、匹配、获取斑点位置坐标等分析。蛋白质点的量表示为该点的所有像素强度值的总和,并将各点含量表示为相对含量,即单个蛋白质点的量占该块胶内所有蛋白质点总量的百分数。
1.2.7 差异蛋白质点的选择 实验共重复3次,每次均采用相同实验条件,以保证较好的可重复性。实验组与对照组各选定一块“平均胶”作为基准胶。所谓“平均胶”即通过对组内各块凝胶进行匹配分析,蛋白质点显示比较均匀,杂质斑点少,最能体现该组蛋白质点分布状态的一块凝胶。差异蛋白质点的选择借助图像分析软件,以平均胶为基准,参考其它胶,选择组间表达差异明显,组内表达含量稳定、位置恒定的点作为目标点。确定差异点后在凝胶上切取相应的点,去离子水洗涤,甲醇固定,-80℃冻干保存。
1.2.8 蛋白质分子的检测 采用MALDI-TOF-S进行蛋白质肽质量指纹谱的分析,蛋白质的鉴定、质谱打分结果和肽段匹配信息等以MASCOT为搜索引擎在NCBInr数据库检索(www.matrixscience.com)。
1.3 相关分子的免疫组化研究
根据质谱分析、蛋白鉴定结果,选取两种与细胞凋亡或药物作用相关的蛋白质,购买抗体,进行免疫组化染色。每种蛋白质染色分三组,正常细胞,乙醇对照组细胞,加泰索帝组细胞。细胞培养72 h,胰蛋白酶消化,PBS冲洗3次,离心收集细胞铺片,用冷丙酮固定10 min,Triton-X 100洗10 min,PBS洗5min×3次;用3%H2O2孵育25 min,阻断内源性氧化酶,PBS洗 5 min×3;正常羊血清50 μ l/片滴于组织片上,室温下孵育20 min,封闭非特异性位点;倒去多余血清,分别滴加选定的蛋白抗体,50 μ l/片,室温 1小时,PBS洗5 min×3;滴加生物素化羊抗兔IgG与玻片上 ,50 μ l/片 ,37℃30min,PBS 洗5 min×3;滴加辣根过氧化物酶(HRP)标记的亲和素,50 μ l/片,37℃30分钟,PBS洗5分钟×3;DAB显色,蒸馏水终止反应,苏木素复染核,弱氨水中分化/盐酸返蓝,酒精逐级脱水,二甲苯透明,树胶封片。显微镜下观察其表达情况。
1.4 Western blot检测蛋白表达
各取50 μ g蛋白 ,上样于12%SDS-PAGE,再转移至硝酸纤维素膜,5%脱脂奶粉封闭1小时,加入蛋白抗体,4℃过夜,次日用TBS洗涤3次(每次15分钟),加入ECL显色液,X光胶片曝光冲洗,以β-actin做内参照。
2 结果
2.1 双向电泳凝胶图
实验共重复3次,泰索帝0.01 μ g/mL作用TA-K细胞72 h后的实验组与对照组分别得到3幅双向电泳凝胶图(图1),以平均胶为参考标准,凝胶图像匹配率分别为80.5%(泰索帝组)和78.5%(溶媒组)。目前认为,凝胶图像匹配率在75%以上即可判定图像重复性较好。由PHOTOSHOP CS软件和PDQuest双向电泳分析软件2.2质谱分析,质谱分析结果见表1。

图1 泰索帝作用前后2-DE凝胶图

表1 差异蛋白质谱分析结果
实验组与对照组相比较,经泰索帝处理后,表达明显上调的蛋白有:多聚(ADP-核糖)聚合酶[poly(ADP-ribose)polymerase,PARP],ADP-核糖焦磷酸酶(ADP-ribose pyrophosphatase),matrin 3,细胞凋亡易感蛋白(cellular apoptosis susceptibility protein,CAS)。表达下降蛋白有:热休克蛋白27(heat shock proteins 27,HSP27),未命名蛋白质(unnamed protein product)。
2.2 PARP与HSP27免疫组化染色
选取PARP抗体TBX2与HSP27抗体,对正常组,乙醇对照组,泰索帝组细胞进行免疫组化染色比较。PARP染色显示,正常组和乙醇对照组PARP变化不大,泰索帝组较乙醇对照组棕色颗粒明显明显增多(见图2、图3)。HSP27染色显示应用泰索帝作用后,HSP27棕色颗粒较正常组和乙醇组减少(见图4,图5),与2-DE灰度变化相一致。
2.3 PARP与HSP27蛋白水平分析
应用PARP抗体与HSP27抗体,对乙醇对照组、泰索帝组蛋白质进行Western blot检测(重复2次),检测结果显示,泰索帝能够增加PARP的表达,降低HSP27的表达(图6)。

图2 溶媒组PARP染色(×40)

图3 泰索帝组PARP染色图片(×40)

图4 溶媒组 HSP染色(×40)
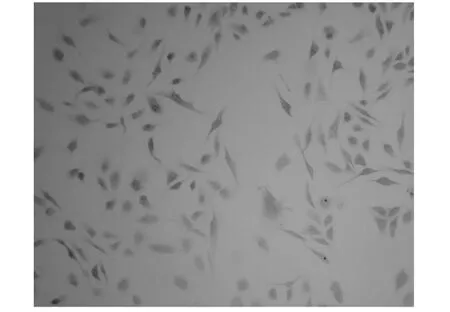
图5 泰索帝组HSP染色(×40)

图6 PARP与HSP27免疫印迹检测
3 讨论
甲状腺未分化癌(ATC)是高度恶性的肿瘤,其化疗在肿瘤局部控制与延缓远处转移上具有重要的意义。但目前的化疗药物与化疗方案,效果欠佳,并不能明显的改善ATC的预后。泰索帝是最应用临近床的一种紫杉类化疗药物,在体外对多种细胞系显示细胞毒性作用,可以直接诱导细胞凋亡,其对非小细胞肺癌、晚期乳腺癌、卵巢癌、胃癌等临床效果显著,化疗效果明显优于不含泰索帝药物的方案。曾有临床试验证实,紫杉醇较其它化疗药物能够延长ATC的生存期[2-4]。且有研究证实,ATC之所以耐药,是因为他能够合成P-gp和MRP[5],这两种蛋白能够把化疗药物排出细胞外,泰索帝细胞内浓度是紫杉醇的3倍,应当是一个比较有希望的化疗药物。既往研究证实对ATC有明显的抑制作用[6-7]。
本实验通选择最佳药物作用浓度0.01μ g/mL,作用时间72小时,与对照组进行蛋白质组学研究。2-DE显示有9个蛋白差异点,应用质谱分析,6个蛋白点质谱显示阳性。灰度分析结合质谱结果显示,上调的蛋白质有PARP,ADP-核糖焦磷酸酶,matrin 3,CAS;下调的蛋白质有HSP27,unnamed protein product。免疫组化与Western blot证实PARP和HSP27的变化与2-DE、质谱分析蛋白鉴定结果相一致。
3.1 多聚(ADP-核糖)聚合酶(PARP)
在细胞受到外界损伤因素作用时,DNA发生断裂,PARP结合到DNA断裂口,其催化活性被激活,以尼克酰胺腺嘌呤二核苷酸(nicotinamide-adenine dinucleotide,NAD+)为底物,催化其裂解为二磷酸腺苷核糖(ADP-ribose)和尼克酰胺两部分,并将ADP-ribose转移至受体蛋白谷氨酸残基上,对多种受体蛋白进行修饰,如染色质蛋白多聚(ADP-核糖)基化后,活性受抑制并与DNA分离、组蛋白多聚(ADP-核糖)基化后,可使染色质结构松散修饰后受体蛋白,进而发生一系列级联反应,使细胞对外界刺激作出应答。如果PARP本身发生多聚(ADP-核糖)基化后,因电荷排斥作用从DNA断口脱落,与DNA分离。PARP可作为细胞内有效监测DNA损伤的分子感受器(molecular sensor),修复损伤或诱导细胞凋亡。
Caspase指的是半胱天冬酶,被认为是凋亡的中心实施者,其执行凋亡的功能依赖于底物蛋白的酶解活性,在Caspase的级联反应中,最终均有Caspase-3的活化。PARP是活化了的Caspase-3的底物之一。在细胞凋亡过程中,Capase-3酶解PARP,使其裂解为2个片段(89 kDa和24 kDa)。24 kDa的PARP片段含有PARP的结合区域,结合到DNA链的断口,因24 kDa的片段缺乏自身修饰域,不能被残存的PARP多聚(ADP-核糖)基化,因此,24 kDa片段将不容易从DNA链的断口释放出来,冻结了DNA断口,使DNA修复酶不能接近,不能对破裂的DNA片段进行修复,促进凋亡的发生。裂解PARP形成的89 kDa片段,因不含结合域,断裂的DNA不能使其活化,防止了细胞内NAD和ATP的过度消耗,为细胞凋亡保存了能量。因此PARP裂解可作为凋亡早期的敏感指标。当大量DNA损伤得不到有效修复时,大量PARP进行自身修饰,集聚的PARP激发caspase对其进行剪切,即紧随PARP核糖化之后PARP便发生了蛋白水解作用,其机制为聚ADP-核糖可增加caspase与PARP的结合力,促进PARP蛋白酶的水解,从而推动细胞凋亡的发生,PARP的”细胞保护”功能便发生了变化[8-9]。这一作用形式,使细胞根据受损程度作出反应,损伤不严重时,诱导细胞进入生长休止状态,修复损伤,以利于细胞功能的恢复;损伤严重时则放弃修复,使细胞进入凋亡。
另有研究显示PARP可以抑制P53的降解诱发细胞凋亡。许多实验已证明在几种不同的生理条件下通过表达P53可启动细胞凋亡,但P53蛋白半衰期极短,在细胞内迅速降解,因而正常细胞内浓度极低。DNA的断裂激活PARP,细胞内爆发大量的核糖化反应,PARP诱发P53基因mRNA蛋白使之发生聚ADP核糖化反应,减少了 P53的降解。另外Malanga等[10]研究显示PARP与P53在胞内分布具有相关性,PARP可调控P53与DNA的结合力,调节P53的功能。
本实验研究显示在泰索帝的作用下,甲状腺未分化癌细胞系中PARP表达上升,这与Negri等研究VP16诱导Hela细胞凋亡过程中,胞内被自身修饰的PARP大量增加相一致,考虑可能与泰索帝的药物作用,使细胞DNA受损,促使PARP的大量自身修饰,促进凋亡的发生有关。
3.2 热休克蛋白27(HSP27)
HSP27分子量(27kDa),是小分子热休克蛋白(small HSPs)sHSPs亚家族中的重要一员,参与调节细胞的增殖、分化和细胞凋亡的信号转导等。研究证实HSP27与抗肿瘤凋亡和肿瘤耐药有关。
本组实验证实,泰索帝作用后HSP27蛋白下调,同时有研究显示,ATC的耐药机制和P-gp有关,HSP27还可降低细胞内P-gp的表达,降低细胞的耐药性,对HSP27的研究,有可能改善ATC对化疗药物的耐受性。
3.3 ADP-核糖焦磷酸酶
ADP-核糖焦磷酸水解酶亚家族可水解ADP-核糖,调控体内ADP-核糖的水平,维持细胞正常代谢。ADP-核糖是 NAD+、环形ADP-核糖和聚ADP-核糖的酶催化代谢物。细胞内高水平的ADP-核糖使体内蛋白质产生非催化型的ADP-核糖化,从而导致蛋白质功能的失活,此外高水平的ADP-核糖还会干扰正常ADP-核糖依赖型酶的生理功能。例如肌动蛋白的聚合可通过在半胱氨酸的ADP-核糖化而被抑制;ADP-核糖化的组蛋白H1的可作为聚ADP-核糖聚合酶的延长识别位点。此外,酶催化的ADP-核糖化也具有某些毒害作用,如在某些系统中的导致凋亡作用和介导细菌性毒素的细胞毒害作用。
本组研究中泰索帝药物作用后ADP-核糖焦磷酸酶的升高,可能是由于多聚药物作用后PARP明显升高,继发ADP酶的升高有关。
3.4 Matrin 3
matrin 3是一个高度保守的核基质DNA结合蛋白,具有正电荷的N末端和高度的带负电荷的C末端[14]。它在信使核糖的与其它核基质蛋白的相互作用中,形成细胞核内相互作用[15]。据报道,在细胞核内,matrin3与肌苷特异性RNA结合蛋白p54和剪接因子PSF形成了一个复合体,形成的复合体可以把超编辑RNA固定核基质上,同时有选择性地释放剪辑mRNAs。matrin3同其它基质蛋白相同,可能参与前体mRNA剪接。在生理情况下matrin 3上的丝氨酸-208,丝氨酸-596,丝氨酸-598,丝氨酸-604可被磷酸化[15],事实上在小鼠脑内N-甲基-D-天冬氨酸(N-methyl-D-aspartate,NMDA)受体激活后,蛋白激酶A将matrin3磷酸化,诱导其降解。蛋白激酶A抑制剂H-89可防止NMDA诱导matrin 3磷酸化和降解,以及神经元死亡。即蛋白激酶A介导的matrin 3磷酸化,作为一种快速的方式传递信息,在生理情况下从突触含有NMDA受体传递到细胞核,并在病理情况下诱导神经元死亡。最近应用蛋白质组学研究发现[16],Matrin-3是钙调素结合蛋白与caspase-3的下游底物,功能受与细胞分裂有关的钙调蛋白和caspase的调节。
本组研究中发现,TA-K细胞经泰索帝药物作用后matrin 3蛋白表达上调,matrin 3参与RNA的代谢,与细胞凋亡有关,具体药物作用后matrin 3是否有促进凋亡的作用或抑制凋亡的作用,还有待于进一步研究。另外matrin 3能够使神经元坏死,matrin 3的变化是否与泰索帝具有神经系统毒性作用有关,尚待进一步研究证实。
3.5 细胞凋亡易感蛋白(CAS)
CAS是由hCAS/CSE1L基因编码的一种酵母基因CSE1人源性同系物。在研究乳腺癌细胞对抗药物毒性引起的细胞凋亡中首次被克隆发现,和细胞增殖和凋亡的多种过程有关[17-18]。CAS在增殖细胞中高表达,在静止细胞和组织中处于低水平。研究显示CAS的表达能够促进细胞的凋亡,抑制细胞生长增殖,可作为肿瘤细胞的一种”保护机制”[19]。
Tanaka等[17]研究证实CAS(hCAS/CSE1L)与P53的一个靶启动子亚群有关,其中包括PIG3。CAS的降解使P53靶启动子的转录减少,启动子所引起的凋亡相关的激活减少,对细胞凋亡具有抑制作用。另外hCAS/CSE1L活性抑制增加PIG3基因的的组蛋白H3赖氨酸27的甲基化。CAS是一种核质的转运因子,与染色质相关,是异染色质延伸的阻遏蛋白。因此CAS能够选择性的激活P53凋亡相关基因的转录,促进凋亡的发生。
综合分析以上结果,泰索帝在分子水平上可能在以下方面对甲状腺未分化癌细胞系TA-K细胞系产生作用:①泰索帝可能导致细胞DNA损伤,大量PARP积聚,产生细胞凋亡或死亡。同时PARP的集聚导致ADP-核糖增多,ADP-焦磷酸酶继发性升高。②通过PARP,CAS,HSP调节P53的表达与代谢,促进细胞凋亡。③下调HSP27使DAXX的活性升高,促进细胞凋亡。④泰索帝影响细胞核内外物质转运,以及细胞核基质的蛋白的调节,影响细胞核内核糖核酸的代谢。⑤HSP27与抗肿瘤耐药有关,泰索帝能够下调HSP27,改善ATC抗肿瘤药物的敏感性。另外泰索帝作用后Matrin-3升高,Matrin-3与NMDA信号途径有关,是否与化疗药物的神经毒性有关,尚需进一步研究。
[1]Asakawa,T Kobayashi,Y Komoike,et al.Establishment of anaplastic thyroid carcinoma cell lines useful for analysis of chemosensitivity and carcinogenesis[J].Journal of Clinical Endocrinology&Metabolism,1996,81(10):3547.
[2]Seiner S,Anderson NL.Pharmacentical p roteomics[J].Ann N Y Acad Sci,2000,919:48.
[3]Sai-Ching JY,Guangpu X,Jingxuan P,et al.Manumycin enhances the cytotoxic effect of paclitaxel onanaplastic thyroid carcinoma cells[J].Cancer Res,2000,60(4):650.
[4]Guangpu X,Jingxuan P,Martin C,et al.Angiogenesis inhibition in the in vivo antineoplastic effect of manumycinand paclitaxel against anaplastic thyroid carcinoma[J].J Clin Endocrinol Metab,2001,86(4):1769.
[5]Jingxuan P,Guangpu X,Sai-ching JY.Cytochrome c release is upstream to activation of caspase-9,caspase-8 and caspase-3 in the enhancedapoptosis of anaplastic thyroid cancer cells induced by manumycin and paclitaxel[J].J Clin Endocrinol Metab,2001,86(10):4731.
[6]付言涛,赵吉生,周 乐,等.泰索帝对人甲状腺未分化癌细胞系(TA-K)增殖抑制作用[J].中国实验诊断学,2008,12(1):16.
[7]付言涛,赵吉生,郑海波,等.泰索帝与阿霉素对甲状腺未分化癌细胞(TA-K)作用比较及对细胞周期的影响[J].中国实验诊断学,2010,14(7):1051.
[8]Sekiguchi M,Shiroko Y,Arai T,et al.Biological characteristics and chemosensitivity profile of four human anaplastic thyroid carcinoma cell lines[J].Biomed Pharmacother,2001,55(8):466.
[9]Viatcheslav A,Soldatenkov MD,Mark Smulson PH.Poly(ADP-ribose)Polymerase in DNA damage-response pathway:implications for radiation oncology[J].Int J Cancer(Radiat Oncol Invest),2000,90(1):59.
[10]Smulson ME,Simbulan Rosenthal CM,Boulares AH,et al.Roles of poly(ADP-ribosyl)ation and PARP in apoptosis,DNA repair,genomic stability and functions of p53 and E2F-1[J].Adv Enzyme Regul,2000,40(2):183.
[11]Rane MJ,McLeish K R,Klein J B,et al.Heat shock protein 27 controls apoptosis by regulatingAkt activation[J].J Biol Chem,2003,278(30):27828.
[12]Schneider J,Marx D,Meden H,et al.Co-expression of the MDR1 gene and HSP27 in human ovarian cancer[J].Anticancer Res,1998,18(4C):2967.
[13]LudwigA,Dietel M,Lage H,et al.Identification of differentially expressed genes in classical and atypical multidrug-resistant gastric carcinoma cells[J].Anticancer Res,2002,22(6A):3213.
[14]Valencia CA,Ju W,Liu R.Matrin 3 is a Ca2+/calmodulin-binding protein cleaved by caspases[J].Biochem Biophys Res Commun,2007,361(2):281.
[15]Tanaka T,Tanaka T,Ohkubo S,et al.hCAS/CSE1L associates with chromatin and regulates expression of select p53 target genes[J].Cell,2007,130(4):638.
[16]Behrens P,Brinkmann U,Wellmann A.CSE1L/CAS:its role in proliferation and apoptosis[J].Apoptosis,2003,8(1):39.
[17]Praveen K,Leary JH 3rd,Evans DL,et al.Molecular cloning of cellular apoptosis susceptibility(CAS)gene in Oreochromis niloticus and its proposed role in regulation of non-specific cytotoxic cell(NCC)functions[J].Fish Shellfish Immunol,2006,20(4):647.
猜你喜欢
肝博士(2022年3期)2022-06-30
海外星云(2021年9期)2021-10-14
陶瓷学报(2021年1期)2021-04-13
军事文摘(2020年20期)2020-11-16
中学生数理化·八年级物理人教版(2020年12期)2020-01-01
中学生数理化·八年级物理人教版(2018年12期)2019-01-31
中国生化药物杂志(2016年5期)2016-07-24
中国科学院院刊(2013年6期)2013-02-24
中国生化药物杂志(2012年4期)2012-01-06
祝您健康(1986年4期)1986-12-30
