
黄土高原中部沙棘与秋胡颓子光合参数之异同
2011-08-04 03:26:20何正祥刘广全1王鸿喆
水资源开发与管理 2011年2期
何正祥,刘广全1,,3,王鸿喆
(1.中国水利水电科学研究院,北京 100048;2.西北农林科技大学,陕西杨凌 712100;3.国际泥沙研究培训中心,北京 100048)
沙棘 (Hippophae rhamnoides L.)和秋胡颓子 (Elaeagnus umbellata Thunb.)同为胡颓子科典型植物种,是生态环境脆弱地带水土保持植被恢复的主要生态经济型树种,具有良好的保持水土、防风固沙、改良土壤功能,抗干旱瘠薄、耐盐碱、抗严寒、耐酷暑和适应性强等特点,而且具有较高的经济价值,为了改善生态环境、应对全球气候变化、振兴区域经济和实现经济社会可持续发展,大力发展、开发、利用和保护沙棘及秋胡颓子资源势在必行。有关沙棘的研究文献很多,光合作用研究的文献也较多[1~7],但是有关秋胡颓子的研究文献较少,所见文献主要集中在分类、栽培和利用上[8~10],全面系统开展沙棘叶片和秋胡颓子叶片光强度响应曲线、CO2浓度响应曲线、光合速率日变化曲线、荧光动力学曲线、淬灭分析、荧光响应曲线、荧光CO2响应曲线及其参数等比较研究未见报道,深入开展此方面研究意义重大。
1 自然概况
研究地点位于陕西省延安市西北农林科技大学延安树木园内,该地属暖温带大陆性季风气候森林草原过渡地带黄土丘陵沟壑区,年平均气温9.4℃,极端最高气温39.9℃,极端最低气温-28.5℃,≥10℃有效积温3268.4℃,1月平均气温-6.7℃,7月平均气温22.9℃。年均降水量550mm,年蒸发量1585mm,年均相对湿度62%,无霜期180d,年日照时数2445h,最大冻土深76cm。土壤以黄绵土为主,母质为黄土,pH值为8.4,土层达100cm以上。天然植被已经不覆存在,多数人工植被发育良好。
2 研究方法
2004年和2005年6~10月在黄土高原中部延安市宝塔区西北农林科技大学延安树木园,选择具有代表性20年生的沙棘和秋胡颓子林等为测定对象,用LICOR-6400光合仪对其光合响应曲线、ACI曲线、光合速率、蒸腾速率、气孔导度和叶绿素荧光动力学曲线、淬灭分析、荧光光曲线以及荧光ACI曲线等光合生理指标进行测定,测定光源分别为红蓝光源和叶绿素荧光叶室光源,每个指标作3个重复,每个重复分别读数5次。
测定光合响应曲线的光照强度取值分别为:0、20、50、100、200、500、1000、1500 和2000μ.molm-2.s-1;测定ACI曲线时光合有效辐射控制在 1500μ mol◦m-2◦s-1,CO2浓度取值为:50、100、200、300、400和600μ mol◦mol-1;光合作用日进程测定时间为:8:00、10:00、12:00、14:00、16:00和18:00;叶绿素荧光生理指标测定前要确定每个叶片的Fo、Fm、Fv/Fm值,多选在上午6:00~7:00之间测定,测定前采用暗适应,用遮光夹进行20min左右的遮光处理,以保证叶片完全的暗适应。
3 结果与分析
3.1 光强度响应曲线
光强度响应曲线反映植物光合速率随光照强度增减的变化规律。从沙棘和秋胡颓子光合速率与光辐射强度的关系曲线可以看出,开始光合速率为负值,即光合产物用于植物本身呼吸消耗,随着光照强度的增加,光合积累与呼吸消耗速率相当,光照强度达到补偿点;之后光照强度增加,所测定树种的光合速率明显增大,当光照强度达到一定值以后,光合速率基本上稳定在一定水平,即达到光饱和 (图1和图2)。

图1 沙棘叶片光照强度响应曲线

图2 秋胡颓子叶片光照强度响应曲线
黄土高原中部沙棘叶片和秋胡颓子叶片的暗呼吸速率、表观量子效率、最大光合速率、光补偿点和光饱和点等参数平均值分别为:-3.6770、-1.7150μ mol◦CO2◦m-2◦s-1,0.05169、0.05487mol◦mol-1,7.367、8.084μ mol◦CO2◦m-2◦s-1、71.15、31.26μ mol◦ m-2◦ s-1和 213.7、178.6μ mol◦m-2◦s-1。可见不同树种光强度响应曲线参数值差别较大,沙棘叶片的暗呼吸速率、表观量子效率和最大光合速率皆小于秋胡颓子叶片的,光补偿点和光饱和点相反。
3.2 CO2浓度响应曲线
从图3和图4可以看出,沙棘叶片和秋胡颓子叶片光合作用速率均随CO2浓度的增加而逐渐增大,当植物光合作用吸收的CO2量与呼吸作用和光呼吸释放的CO2量达到动态平衡时,环境中CO2的浓度为CO2补偿点;之后随着CO2进一步提高,植物光合作用速率迅速上升,几乎呈线形变化,直线的斜率被称为羧化效率,其值大小反映相同CO2浓度的光合作用效率的高低。当CO2的浓度升高达到一定值时,光合速率值达到最大,此时的CO2浓度为CO2饱和点,之后随CO2浓度增加,其光合速率值相对稳定或略有下降,原因可能是CO2浓度过高时引起细胞中pH值的改变,叶片保卫细胞膨压下降,气孔开度受到影响,进一步影响净光合速率。

图3 沙棘叶片CO2浓度响应曲线

图4 秋胡颓子叶片CO2浓度响应曲线
测定结果表明,沙棘叶片和秋胡颓子叶片CO2浓度响应曲线参数值分别为:CO2补偿点150.00μ mol◦mol-1、99 ◦00μ mol◦mol-1,CO2饱和点506.83370μ mol◦mol-1、490.0662μ mol◦mol-1,暗呼吸速率 -1.100μ mol◦CO2◦m-2◦s-1、-2.349μ mol◦ CO2.m-2◦ s-1, 羧化效率0.00630mol◦mol-1、0.02446mol◦mol-1,最大光合速率 3.01μ mol◦ CO2◦m-2.s-1、14.80μ mol◦CO2◦m-2◦s-1。秋胡颓子叶片的羧化效率和最大光合速率远高于沙棘叶片的,暗呼吸速率和CO2补偿点却低于沙棘叶片的,CO2饱和点二者差异不大,可见秋胡颓子对CO2补偿反应敏感,且利用率更高。
3.3 光合速率日进程
沙棘叶片和秋胡颓子叶片光合速率 (Pn)日变化都呈现为单峰曲线,峰值出现在10:00~12:00时左右,光合速率日变化从早晨日出起逐渐升高,出现峰值后光合速率逐渐降低。沙棘叶片和秋胡颓子叶片最大净光合速率平均值分别为16.3547 ±0.7128 、 17.7742 ±0.7245μ mol◦CO2◦m-2◦s-1,二者差别不大;光合速率逐渐降低,之后随着夜间的来临其光合速率逐渐降低,沙棘叶片和秋胡颓子叶片最低净光合速率平均值分别为 3.4806 ±0.7377、9.0465 ±0.5145μ mol◦CO2◦m-2◦s-1,后者是前者的2.6倍 (表1)。
植物的光合速率除了主要受叶片气孔的调节外,还受叶片所吸收的净辐射、风速、大气温度、空气湿度、土壤水分等多种因子的影响。叶片气孔导度 (Gs)的日变化与光合速率的日变化呈现出很好的相关性。在中午 12:00~14:00时,大气温度高达最高,从而限制了叶片的光合速率。光合有效辐射 (PAR)是影响植物光合速率大小的主要因素之一,光合有效辐射日变化曲线走势呈明显的单峰曲线,即早晚低、中午高,而且峰值出现在中午10:00~12:00之间。植物叶片气孔限制值大小与其光合速率呈现明显的负相关关系,但是在中午气孔限制值达最低时,部分植物光合速率也到达最低值,按照Farquhar和Sharke的观点,部分植物发生 “午休”现象主要原因是非气孔因素造成,即叶肉因素对光合速率的限制起主要作用。

表1 沙棘叶片和秋胡颓子叶片光合速率日变化特征
3.4 荧光动力学曲线
植物荧光动力学曲线是研究植物叶片从黑暗条件下转入光下的活化过程中各个参数的变化。一般而言,暗适应的植物叶片突然照光后,荧光上升,但是光合速率等指标并没有明显变化,没有随着高光照而表现出高的光合速率,这是由于两方面的条件限制的:酶活性的激活和气孔的诱导开放,这时往往气孔导度很低。电子传递速率(ET R-Electron Transport Rate)与植物光合速率有很强的线性关系,是又一个表征植物光合能力高低的变量。从图5和图6中可以看出,随着光照时间的加大,所测定的树种的ETR逐渐上升并在一定时间后达到稳定,与光合作用速率同步,只是不同树种达到稳定所需要的时间不同,变化的斜率也有较大的差异。PhiPSⅡ指PSⅡ所吸收的光量子中用于光化学反应的比例,在低光照条件下,PSⅡ的量子产量通常较高,因为叶片所吸收的光能中有较大的比例被用于光化学反应中;而在高光照强度条件下,因为叶片所吸收能量中的很大比例通过非化学过程而散失,所以经过高光照强度光适应的叶片的PhiPSⅡ较低。结果表明,沙棘叶片和秋胡颓子叶片的PhiPSⅡ随着光照时间逐渐上升,稳定时间与光合同步。
当叶片从黑暗条件转入光下,PSⅡ反应中心逐渐关闭,这造成叶绿素荧光产量 (1秒钟之内)的上升,在此之后,荧光产量开始下降,持续大约几分钟或几十分钟,这就是荧光淬灭。首先,电子被从PSⅡ传递走的速率开始上升,这是由于光诱导对C代谢酶的活化和气孔开放的活化,这种淬灭称为光化学淬灭 (qP-Proportion of Open PSII);同时,能量转化为热能的效率也提高了,这种过程称为非光化学淬灭(NPQ-Non Photochemical Quenching)。图 5和图6的结果表明,在黄土高原中部测定的沙棘叶片和秋胡颓子叶片从黑暗条件转入光下,qP和NPQ随着光照时间逐渐上升,达到一定的时间后突然减低,然后逐渐上升,在经过一段时间之后完成淬灭并达到相对稳定的状态。
3.5 淬灭分析 (三相分离)

图5 沙棘叶片荧光动力学曲线参数变化

图6 秋胡颓子叶片荧光动力学曲线参数变化
非光化学淬灭NPQ和热耗散线性相关,在一个典型的植物中,在饱和光强下大致在0.5~3.5之间。这取决于物种和植物以前的经历过程。qN是非光化学淬灭的比较传统的老的术语,有的时候也会被用到。它的范围在0~1.0之间,因此在淬灭较高时不很敏感。同样的淬灭在参考点淬灭较高时可能会表现为很小的上升,因此直接的比较不很明确。通常,如果暗适应的Fv/Fm明显不同,那么NPQ也不能直接进行比较。一般而言,非光化学淬灭的增长可能是由于叶片为免受光破坏的保护机制。研究此过程的一种方法是照光后弛豫的速率。不同的过程有不同的弛豫速率。其时间范围从几分钟到几小时。
图7和图8显示了沙棘叶片和秋胡颓子叶片在照光后弛豫的速率。就是对光适应的叶片转入黑暗下之后一个小时之内各种荧光参数的变化动态。从图中可看出最大荧光产量Fm'(maximal fluorescence during a saturating light flash)在叶片转入黑暗后逐渐上升,在17min左右趋于稳定。qP(Photochemical quenching)变化不明显,原因可能是关闭了活化光。qN变化曲线不很明显,原因可能是在淬灭较高时不很敏感。当光适应的叶片转入黑暗下之后,NPQ开始下降,在25min左右达到稳定。正常条件下,在利用瞬时光能时,低光照强度适应叶片的光能捕获效率更高。在低光照强度条件下,没有多余的光合有效辐射,因此允许大多数植物具有更高的光能捕获效率。

图7 沙棘叶片荧光淬灭动力学曲线参数变化
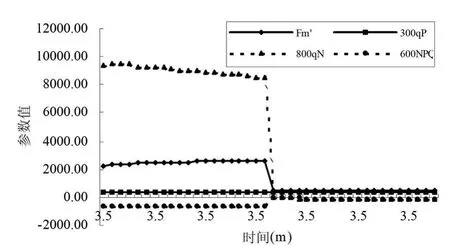
图8 秋胡颓子叶片荧光淬灭动力学曲线参数变化
3.6 荧光光强响应曲线
植物荧光光照强度响应曲线与植物普通光强度响应曲线的变化规律类似(图9和图10),荧光光照强度为零时,植物光合速率为负值,光合产物用于植物本身呼吸消耗,随着光照强度的增加,光合积累与呼吸消耗速率相当,光照强度达到补偿点 (LCP);之后光照强度增加,植物光合速率明显增大,当光照强度达到一定值以后,光合速率基本上稳定在一定水平,即达到光饱和(LSP)。
沙棘叶片和秋胡颓子叶片荧光响应曲线参数暗呼吸速率、光的补偿点、光的饱和点、最大光合速率和表观量子效率其值分别为-0.5106μ mol◦CO2◦m-2◦s-1、-1.3670μ mol◦CO2◦m-2◦s-1,14.72μ mol◦m-2◦s-1、 28.94μ mol ◦ m-2◦ s-1,259.6μ mol◦ m-2◦ s-1、321.3μ mol ◦ m-2◦ s-1,8.494μ mol◦CO2◦m-2◦s-1、 13.810μ mol◦CO2◦m-2◦s-1和 0.03469mol◦mol-1、0.04723mol◦mol-1,植物叶的荧光光响应参数与光合作用参数一样,主要是植物的遗传特性所决定,不同植物种类存在一定的差异。

图9 沙棘叶片荧光光强响应曲线

图10 秋胡颓子叶片荧光光强响应曲线
在植物荧光光照强度响应曲线的实验中,将从一个光适应的植物开始,通过减少瞬时光照强度,目的是为了逐渐得到更高的量子效率和量子产量。PhiPsⅡ指通过荧光计算的PSⅡ量子产量,从图中看出,随着光照强度的减弱PhiPsⅡ逐渐增高,说明叶片吸收的光能中用于光化学反应的比例逐渐增大。在一定的程度上,随着光强度的逐渐降低,PhiPsⅡ值几乎呈直线上升,光强度拐点值一般在 200~500μ mol◦m-2◦s-1之间,不同树种存在一定的差异;达到该值之后,光强度逐渐降低,PhiPsⅡ缓慢上升。PhiCO2是通过CO2同化作用计算的光量子产量,光强度在2000~25μ mol◦m-2◦s-1之间,树木叶的PhiCO2呈缓慢上升趋势,当光强度降到25μ mol◦m-2◦s-1左右时,PhiCO2值突然下降或上升。树种叶子光强度在 2000~25μ mol◦m-2◦s-1之间降低时,qP 值与光强呈显著的线形负相关关系;而qN、NPQ和ET R值与光强呈的线形正相关关系 (图11和图12)。
3.7 荧光C O2响应 (ACI)曲线
木本植物荧光CO2强度响应曲线与普通光源CO2强度响应曲线变化规律相似,即植物光合作用速率均随CO2浓度的增加而逐渐增大,当植物光合作用吸收的CO2量与呼吸作用和光呼吸释放的CO2量达到动态平衡时;之后随着CO2进一步提高,植物光合作用速率迅速上升,几乎呈线形变化,其值大小反映相同CO2浓度的光合作用效率的高低。当CO2的浓度升高达到一定值时,光合速率值达到最大,之后随CO2浓度增加,其光合速率值相对稳定或略有下降(图13和图14)。

图11 沙棘叶片荧光光强响应曲线参数变化

图12 秋胡颓子叶片荧光光强响应曲线参数变化
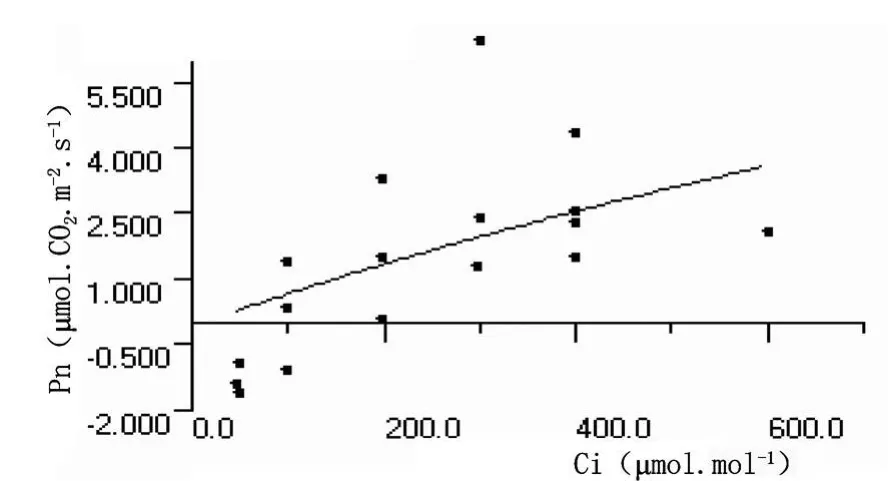
图13 沙棘叶片荧光CO2响应曲线
沙棘叶片和秋胡颓子叶片的荧光CO2响应曲线参数分别为:暗呼吸速率、CO2补偿点、CO2饱和点、最大光合速率以及羧化效率数值分别为 -0.0914、-1.0180μ mol◦CO2◦m-2◦s-1,87.12、 117.80μ mol ◦ mol-1, 745.2123、832.7138μ mol ◦ mol-1, 17.90、22.40μ mol◦CO2◦m-2◦s-1和 0.00769、0.01292mol◦mol-1。

图14 秋胡颓子叶片荧光CO2响应曲线
沙棘叶片和秋胡颓子叶片荧光ACI曲线参数PhiPsⅡ、PhiCO2、qP、qN、NPQ和 ET R变化中,随着CO2浓度的降低qN的值保持一个相对稳定的数值,NPQ呈现逐渐上升的趋势;其它参数PhiPsⅡ、PhiCO2、qP和ETR值随着CO2浓度的降低而降低,并且彼此呈显较好线形正相关关系 (图15和图16)。

图15 沙棘叶荧光ACI曲线参数关系

图16 秋胡颓子叶荧光ACI曲线参数关系
4 小结
(1)沙棘叶片和秋胡颓子叶片开始光合速率为负值,即光合产物用于植物本身的呼吸,随着光强的增加,光合速率与呼吸速率相当,之后光强增加光合速率明显增大,当光照强度达到一定值以后,光合速率基本上稳定在一定水平;二者光强响应曲线参数暗呼吸速率、表观量子效率、最大光合速率、光补偿点和光饱和点平均值分别为:-3.6770μ mol ◦ CO2◦ m-2◦ s-1、-1.7150μ mol◦CO2◦m-2◦s-1,0.05169mol◦ mol-1、0.05487mol◦mol-1,7.367μ mol◦CO2◦m-2◦s-1、8.084μ mol◦CO2◦m-2◦s-1、71.15μ mol◦m-2◦s-1、31.26μ mol◦m-2◦ s-1和 213.7μ mol◦m-2◦ s-1、178.6μ mol◦m-2◦s-1。
(2)沙棘叶片和秋胡颓子叶片光合速率随CO2浓度的增加而逐渐增大,当植物光合吸收的CO2量与呼吸和光呼吸释放的CO2量达到动态平衡时,之后随着CO2浓度提高,植物光合速率迅速上升,几乎呈线形变化,延续到光合速率值达到最大,再后光合速率相对稳定或略有下降。测定二者叶片CO2浓度响应曲线参数CO2补偿点,CO2饱和点,暗呼吸速率,羧化效率及最大光合速率分别为 CO2补偿点150.00μ mol◦mol-1、 99.00μ mol ◦ mol-1,506.83370μ mol◦mol-1、 490.0662μ mol◦mol-1,-1.100μ mol ◦ CO2◦ m-2◦ s-1、-2.349μ mol◦CO2◦m-2◦s-1,0.00630mol ◦ mol-1、0.02446mol◦mol-1,3.01μ mol◦CO2◦m-2◦s-1、14.80μ mol◦CO2◦m-2◦s-1。
(3)沙棘叶片和秋胡颓子叶片光合速率日变化为单峰曲线,其峰值出现在10:00~12:00时左右,最大值分别为 16.3547±0.7128、17.7742±0.7245 μ mol◦CO2◦m-2◦s-1, 最低值分别为 3.4806±0.7377、9.0465±0.5145μ mol◦CO2◦m-2◦s-1;实际上,植物的光合速率除了主要受叶片气孔的调节外,还受叶片所吸收的净辐射、风速、大气温度、空气湿度、土壤水分等多种因子的影响。
(4)随着光照时间的加大,沙棘叶片和秋胡颓子叶片的ET R逐渐上升并在一定时间后达到稳定,与光合作用速率同步。沙棘叶片从黑暗条件转入光时,qP和NPQ随着光照时间逐渐上升,达到一定的时间后突然减低,然后逐渐上升,在经过一段时间之后完成淬灭并达到相对稳定的状态。
(5)在照光后弛豫的速率,就是对光适应的叶片转入黑暗下之后一个小时之内各种荧光参数的变化动态。沙棘叶片和秋胡颓子叶片最大荧光产量Fm'在叶片转入黑暗后逐渐上升,在17min左右趋于稳定;qP和qN变化不明显;当光适应的叶片转入黑暗下之后,NPQ开始下降,25min左右达到稳定。
(6)沙棘叶片和秋胡颓子叶片荧光强度响应曲线与其普通光强响应曲线的变化规律类似,荧光响应曲线参数暗呼吸速率、光的补偿点、光的饱和点、最大光合速率和表观量子效率其值分别为 -0.5106μ mol◦ CO2◦ m-2◦ s-1、-1.3670μ mol◦CO2◦m-2◦s-1,14.72μ mol◦m-2◦s-1、28.94μ mol◦m-2◦s-1,259.6μ mol◦m-2◦s-1、321.3μ mol◦m-2◦s-1,8.494μ mol◦CO2◦ m-2◦s-1、13.810μ mol◦CO2◦m-2◦s-1和 0.03469mol◦ mol-1、0.04723mol◦mol-1。
(7)沙棘叶片和秋胡颓子叶片荧光CO2响应曲线与普通光源CO2响应曲线变化规律相似,荧光CO2响应曲线参数暗呼吸速率、CO2补偿点、CO2饱和点、最大光合速率以及羧化效率分别为 -0.0914μ mol◦CO2◦m-2◦ s-1、-1.0180μ mol◦CO2◦m-2◦s-1,87.12μ mol ◦ mol-1、117.80μ mol ◦ mol-1,745.2123μ mol ◦ mol-1、832.7138μ mol◦mol-1,17.90μ mol◦CO2◦m-2◦s-1、22.40μ mol◦CO2◦m-2◦s-1和0.00769mol◦mol-1、0.01292mol◦mol-1。
[1] 阮成江,李代琼.半干旱黄土丘陵区沙棘的光合特性及其影响因子[J].植物资源与环境学报,2000,9(1):16-21.
[2] 刘瑞香,杨吉力,高丽.不同土壤水分条件对中国沙棘和俄罗斯沙棘的光合和蒸腾作用的影响[J].内蒙古大学学报(自然科学版),2006,37(2):163-180.
[3] 蒋高明,林光辉.几种荒漠植物与热带雨林植物在不同CO2浓度下光合作用对光照强度的反应[J].植物学报,1996,39(12):972-991.
[4] 孟函宁,刘明国,刘青柏,等.阜新地区不同沙棘品种光合及蒸腾特性的研究 [J].沈阳农业大学学报,2007,38(3):345-348.
[5] 李文华,朱清科,赖亚飞,等.陕北黄土高原丘陵沟壑区沙棘的光合特性研究 [J].干旱地区农业研究,2007,25(1):135-139.
[6] 张玮,李晓燕,李连国,等.内蒙古西部地区沙棘光合与蒸腾特性研究[J].华北农学报,2007,22(3):97-100.
[7] 唐道锋,贺康宁,朱艳艳,等.白榆沙棘光合生理参数与土壤含水量关系研究 [J].水土保持研究,2007,14(1):230-233.
[8] 柴发熹.牛奶子营养成分及开发利用途径[J].林业科技开发.1995(1):50.
[9] 徐晓丹,郑伟,钟晓红.胡颓子属种质资源开发利用研究进展 [J].湖南农业科学,2007,(5):44-46.
[10] 朱笃,徐曲.胡颓子果实营养成分的测定[J].江西师范大学学报 (自然科学版),2000,24(1):90-91.
[11] 武丽君,毕润成.不同生境伞花胡颓子光合生理特征的比较[J].山西师范大学学报 (自然科学版),2009,23(2):78-82.
猜你喜欢
今日农业(2022年13期)2022-09-15 01:21:40
中国水土保持(2022年3期)2022-03-24 12:15:40
昆明医科大学学报(2021年8期)2021-08-13 08:59:28
中学生数理化·高一版(2020年6期)2020-07-25 01:47:24
西藏农业科技(2019年1期)2019-07-25 00:37:10
知识经济·中国直销(2018年11期)2018-11-26 01:18:48
中央民族大学学报(自然科学版)(2018年3期)2018-11-09 01:16:32
中学生数理化·八年级物理人教版(2017年3期)2017-11-09 03:05:33
上海金属(2014年5期)2014-12-20 07:58:39
食品工业科技(2014年23期)2014-03-11 18:19:01
