
濒危树种金钱松RAPD体系的建立和遗传多样性分析
2011-07-30 10:40:38高燕会樊民亮骆文坚黄华宏童再康
浙江农林大学学报 2011年5期
高燕会,樊民亮,骆文坚,黄华宏,童再康
(1.浙江农林大学 生物技术研究所,浙江 临安 311300;2.浙江省林业种苗管理总站,浙江 杭州 310020)
金钱松Pseudolarix amabilis又名金松,是中国特有的单种属植物。地质年代的白恶纪金钱松曾经在亚洲、欧洲、美洲都有分布,更新世的冰河时代各地金钱松都相继灭绝,唯有中国长江中下游残留少数,成为现今仅存于中国的单属单种特有的植物[1-4]。由于其特殊的分类地位,金钱松在裸子植物松科Pinaceae系统发育、古生态和古气候的研究方面具有重要意义。这一宝贵的植物遗产被中国定为国家二级保护植物,分布于江苏、安徽、浙江、江西、湖南等地,喜光爱肥,适宜酸性土壤。金钱松树干通直,材性优良,纹理直,耐水湿,可供建筑、桥梁、船舶及家具等用材,而且树形优美,秋季叶呈金黄色,是优良的绿化和庭院观赏树种。种子可榨油,树皮和根皮(中药名为土槿皮或土荆皮)有止痒杀虫的功能,外用于治疗手脚癣、神经性皮炎、湿疹[5-6]等。目前,国内外对金钱松的研究多集中在生物学和生理学特征方面[7],例如叶表皮的扫描电镜观察[8]、 种子品质[9]、 栽培繁殖技术[10-11]、 生长情况[11]、 化学成分研究[12]。也有研究金钱松细胞分类学的报道,认为根据其核型分析,应该单独建立一个金钱松亚科[13-14]。零星的还有组织培养[15]的报道。而对金钱松遗传多样性的研究报道较少。随机扩增多态性DNA(radom amplified polymorphic DNA,RAPD)分子标记技术操作简单易行,具有多态性高,无需活材料,能实现全基因组无偏取样和无组织特异性等,在濒危植物遗传变异方面有较为广泛的应用[16-18]。此外,还广泛地应用于松类植物遗传多样性和遗传结构的研究中,如多脂松Pinus resinosa,欧洲赤松Pinus sylvestris,辐射松 Pinusradiata,红松 Pinuskoraiensis,黄山松 Pinustaiwanensis,马尾松 Pinusmassoniana 等树种[19-24]。本研究采用RAPD分子标记技术对浙江省内不同来源的金钱松遗传多样性进行研究,旨在为合理地保护、利用与开发金钱松遗传资源提供可靠的科学依据。
1 材料与方法
1.1 试验材料
供试材料采自浙江省安吉县灵峰寺林场,共9个来自金钱松天然林的62个单株(表1),采集叶芽,于-70℃的低温冰箱中保存备用。
1.2 试验方法
1.2.1 金钱松基因组DNA的提取 金钱松基因组DNA的提取采用十六烷基三甲基溴化铵(CTAB)-硅珠法[25]。
1.2.2 金钱松随机扩增多态性DNA-聚合酶链式反应(RAPD-PCR)扩增体系的建立[25-27]针对Taq DNA聚合酶量,镁离子(Mg2+),模板DNA,三磷酸碱基脱氧核苷酸(dNTPs)和引物,采用5因子4水平的正交设计L16(45)设计方案详(表2)进行试验,共16个处理,重复2次·处理-1,在Gene Amp PCR System 9700扩增仪上进行PCR扩增。RAPD-PCR扩增反应程序为94℃预变性4 min,38个PCR循环(94℃变性30 s,38℃退火30 s,72℃延伸2 min),72℃延伸7 min。扩增的反应产物用10.0 g·kg-1的琼脂糖凝胶电泳检测,于Gel Doc XR凝胶成像系统拍照分析。
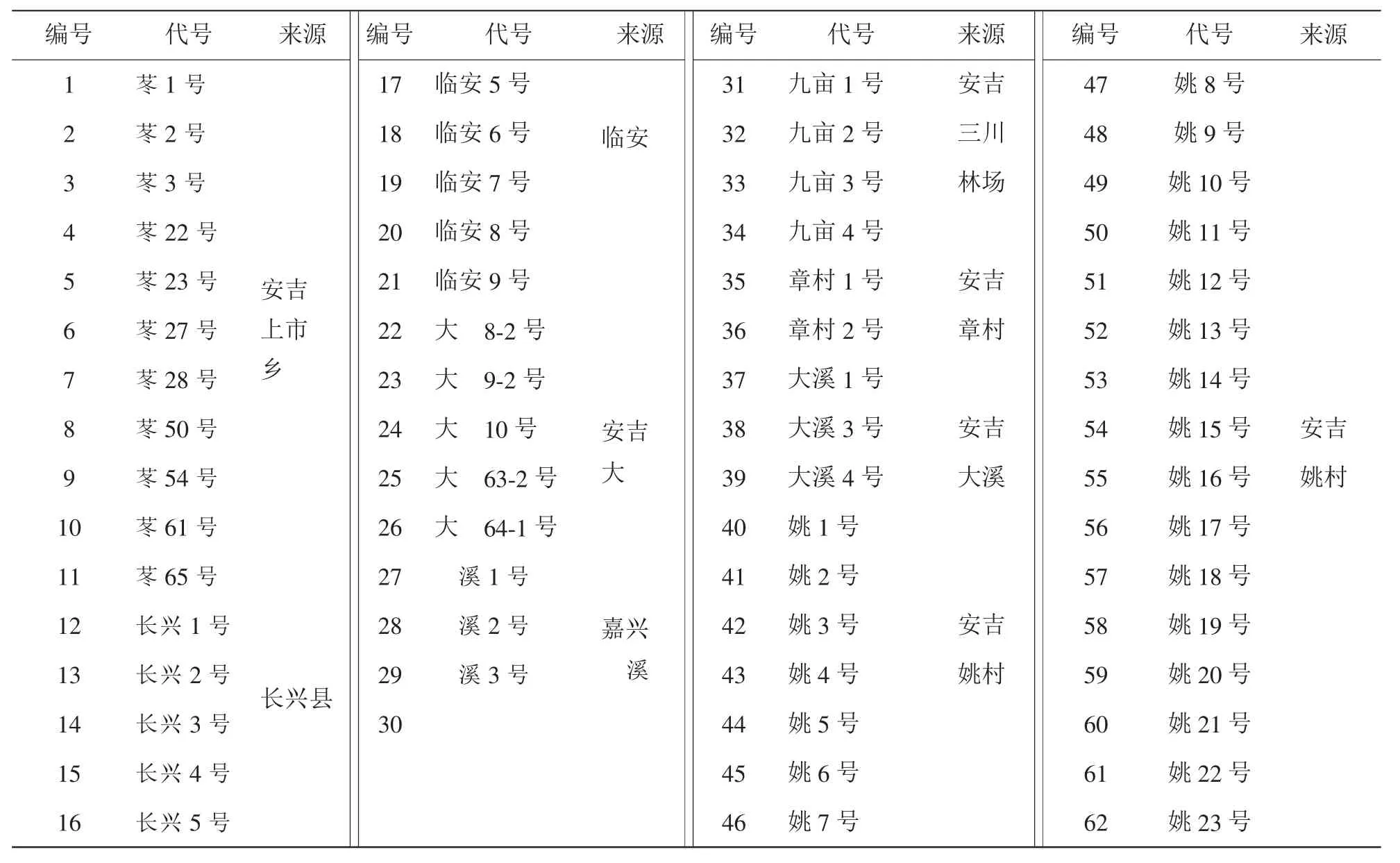
表1 供试材料编号及其来源Table 1 Code and source of the materials investigated

表2 PCR扩增体系各成分因素-水平正交设计表L16(45)Table 2 Orthoronal design for PCR
1.2.3 不同退火温度的梯度试验 用以上处理条件所得到的最佳反应体系进行退火温度梯度试验,设置退火温度。在PCR Express(HyBAID)扩增仪上自动生成12个温度梯度:30.0,30.3,30.9,31.8,32.9,34.7,35.5,36.9,38.4,39.3,39.8,40.1℃,比较退火温度对电泳谱带的清晰度和数量的影响,从而确定最佳退火温度。
1.2.4 引物筛选 引物筛选包括初筛和复筛。用1个样品对200个随机引物进行初筛,选择谱带清晰的引物用于复筛。复筛时用4个不同种源各1个样品进行扩增,选择出清晰、多态性较高的引物用于RAPD-PCR扩增。
1.2.5 数据统计分析 对金钱松62个单株进行RAPD-PCR扩增的电泳谱带总条数和多态性条带进行统计。根据分子标记在相同电泳迁移率的有无统计得到所有位点的二元数据;有DNA扩增有带记为1,无带记为0,强带和可分辨的弱带赋值为1。扩增条带与标准分子量的迁移率相比对照,相同引物扩增出来的同一长度(同一电泳迁移率)的带视为同一位点。将得到的二元数据应用SPSS 16.0软件进行分析,并计算其欧氏遗传距离,按Average Linkage对金钱松种质资源进行聚类分析,建立聚类树状图。
2 结果与分析
2.1 金钱松RAPD-PCR最佳反应体系的建立
2.1.1正交试验的直观分析 根据本试验的正交设计进行PCR的扩增结果和电泳检测,依据琼脂糖电泳条带的强弱和杂带的多少做直观分析。正交试验设PCR扩增结果是由镁离子(Mg2+),dNTPs,引物,Taq DNA聚合酶和模板DNA等因素综合作用的结果,因而扩增结果存在明显差异。第1,2,3,4组合谱带较弱,其原因可能是由于TaqDNA聚合酶的浓度太低而引起的;由于TaqDNA聚合酶的浓度一定时,镁离子(Mg2+)浓度的过低(第5,6,9,10组合扩增不出条带),而镁离子(Mg2+)浓度过高(第8组合扩增条带拖尾且模糊)会影响扩增的效果,第7组合条带清晰且多态性高;当Taq DNA聚合酶和镁离子(Mg2+)浓度浓度升高时,会扩增出非特异性条带(如第11,12组合)。通过多因素综合比较选择第7组合(1.0 × 16.67 nkat TaqDNA 聚合酶,2.5 mmol·L-1镁离子(Mg2+),50 ng 模板DNA,0.1 mmol·L-1dNTPs,0.5 μmol·L-1引物)扩增的谱带不但多态性好,重复性好,且谱带清晰,是金钱松RAPD-PCR扩增的最佳反应体系。
2.1.2 金钱松RAPD-PCR最佳退火温度的确定 退火温度是影响PCR的重要因素之一。退火温度不仅与引物序列有关,还与物种DNA的序列密切相关[28],因此,确定最佳退火温度对获得稳定可靠的ISSRPCR结果非常重要。金钱松RAPD-PCR温度梯度PCR的电泳结果见图1,在12个温度梯度下30.0,30.3,30.9,31.8,32.9和34.7℃扩增的条带较弱,不能正确地表达其引物的多态性,35.5,36.9和38.4℃引物扩增出清晰明亮的RAPD条带。39.3,39.8和40.1℃温度过高时,扩增出的条带有拖尾,所以退火温度过高或过低都会影响到PCR扩增条带的清晰度和多态性的正确表达,综合多方面的因素,最终选择38.0℃为金钱松RAPD-PCR扩增最适宜的退火温度。
2.2 扩增产物的多态性分析
通过初筛和复筛共得到的18条多态性引物(表3),18条RAPD引物对62个供试材料进行RAPD-PCR扩增,共扩增出172条带,其中170条带具有遗传多态性,多态性比率为99.2%。在62个供试材料中,不同RAPD引物的总位点差异较大,从3~20条不等,平均为9.56条,扩增出的DNA片段大小分布为200~3 000 bp;其中扩增条带数最多的为引物S53,共扩增出20条带,说明不同引物与供试材料总DNA部分区域的同源性有较大差异。可以说明:金钱松种子园群体内的DNA多态性是很丰富的,遗传变异幅度很大,可能与野生类群分布大范围较广,分布区内环境复杂多样有关,也可能同其复杂的遗传背景与起源有关,因而这些丰富的变异类型为选育观赏新品种奠定了物质基础。

图1 正交试验PCR产物电泳图Figure 1 Theelectrophoresis map of PCR products of the orthogonal tests
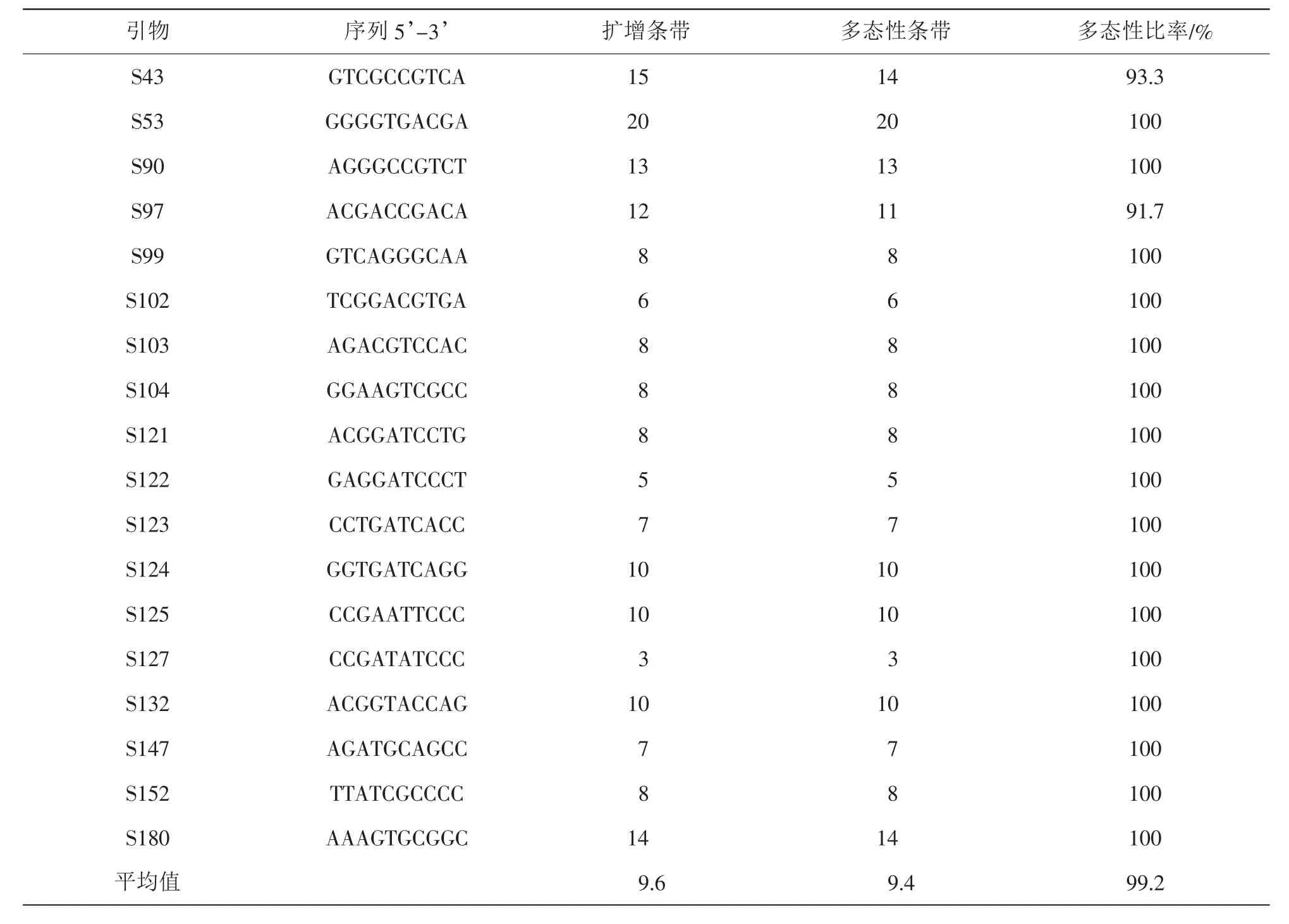
表3 18个引物扩增出DNA片段数Table 3 DNA fragments amplified with 18 primers
2.3 聚类分析
将18条引物扩增后得到的二元数据应用SPSS 16.0软件进行分析,对供试材料根据RAPD标记欧氏遗传距离,按Average Linkage对62份金钱松种质资源进行聚类分析,建立聚类树状图(图2)。由图2中以看出,利用RAPD分子标记的结果可以将62份金钱松种质资源聚为三大类群:第一大类群是包括由采集于安吉大、安吉三川林场、安吉章村、安吉大溪以及安吉姚村的不同材料,第二大类包括采自长兴、临安以及嘉兴的材料,第三大类包括采自安吉上市乡的材料,其中,同一来源的材料几乎都聚在一起,说明金钱松天然群体RAPD的多态性分类和金钱松的地理分布有一定的关系。但也有例外,如来自临安野生种群的17号和来自长兴野生种群的16号就分别和来自安吉大的野生种群聚在一起,可能是遗传变异的结果;来自安吉大溪的39号较为特殊,与其他个体的遗传距离为0.280 0~1.255 7,表明金钱松天然种群内存在较高的的遗传多样性。
3 讨论
3.1 金钱松遗传多样性分析
遗传多样性是指种内基因的变化,包括种内不同的种群间和同一种群内个体的遗传变异,种内多样性是物种及以上各水平多样性的最重要来源。遗传变异、生活史特点、种群动态及其遗传结构等决定或影响着一个物种与其他物种及其环境相互作用的方式,而且种内的多样性是一个物种对人类影响进行反应的决定因素。种内的遗传多样性多发生在分子水平,并且都与核酸的理化性质紧密相关,新的变异是突变的结果[29]。
濒危植物的遗传多样性水平一般较低。本研究通过RAPD技术研究了浙江省内的金钱松的多态性位点的比率是99.2%,这一结果表明:分布于浙江省的金钱松居群具有较为丰富的种内遗传多样性,这与刘建锋等[16-17]对濒危植物崖柏Thuja sutchuenensis的遗传多样性的研究结论相一致:濒危植物遗传多样性具有一定的变化范围。说明并非所有的珍稀濒危植物都存在低水平的遗传多样性。
本研究通过RAPD分子标记得到的金钱松的聚类图中,同一来源的材料几乎都聚在一起,但也有个别的材料和其地理分布并不完全相同,如来自临安的17号、来自长兴的16号以及来自安吉大溪的39号,这可能是由于不同地理条件的环境因素或者是由于种群内的基因突变所引起的,这些突变经自然选择一些中性突变通过随机过程整合到基因组中,即可形成丰富的分子水平丰富的遗传多样性。
3.2 金钱松种质资源的保护策略
造成金钱松濒危的原因主要有:一是残存的个体稀少,分布零星,加上人为破坏,使其生存环境受到严重威胁,资源日渐减少;二是金钱松结实有大小年之分,一般3~5 a丰产1次,对其大量繁殖有明显地影响。
本研究中,金钱松的遗传多样性非常丰富,能为植物遗传改良提供理论指导。通过对金钱松种子园的遗传结构分析可以获得群体遗传多样性、遗传变异分布式样等方面的重要数据,对实现物种的有效管理和制定合理的保护策略有重要参考价值[29-30]。因此,结合金钱松的群体遗传多样性,对现存的金钱松种质资源采取就地保存策略时,重视大范围群体内不同类型个体的保存,尽可能防止人为破坏带来的遗传资源的流失。另一方面,由于浙江省内金钱松的遗传多样性较丰富,因此,在对金钱松种质资源实施保存时,不能仅考虑到群体内不同个体的保存,同时也应对不同地理区域的群体进行保存,可以进行迁地保护[32],尽可能多地收集其他地区的金钱松种质材料,建立种质资源库。本研究即是通过对建立浙江省安吉县灵峰寺林场的金钱松的种子园,可以为此加大种群间的交换,为基因交流和重组创造条件,以保存孑遗金钱松的遗传多样性和金钱松的种质资源。

图2 金钱松种质资源聚类图Figure 2 Dendrogram of Pseudolarix amabilis germ plasm resource
[1]李楠.世纪论松科植物的地理分布、起源和扩散[J].植物分类学报,1995,33(2):105-130.LI Nan.Studies on the geographic distribution,origin and dispersal of the family Pinaceae Lindl.[J].Acta Phytotaxon Sin,1995,33(2):105-130.
[2]应俊生.中国裸子植物分布区的研究(1)松科植物的地理分布[J].植物分类学报,1989,27(1):27-38.YING Junsheng.Areography of the gymnosprerms of China(1)distribution of the Pinaceae of China [J].Acta Phytotaxon Sin,1989,27(1):27-38.
[3]王冬,梁鹏,刘晓菊.金钱松高效栽培技术[J].现代农业科技,2009(3):57.WANG Dong,LIANG Peng,LIU xiaoju,The high effect cultivation techniques of Pseudolarix amabilis(Nelson)Rehd.[J].Mod Agric Technol,2009(3):57.
[4]姚志刚,陈玉清.榉树和金钱松在江苏的资源及保护[J].林业科技开发,2001,15(6):16-18.YAO Zhigang,CHEN Yuqing.Resources and protection for Zelkova and Pseudolarix amabilis(Nelson) Rehd.[J].China For Sci Technol,2001,15(6):16-18.
[5]王章荣,陈天华,黎章矩,等.金钱松遗传资源调查收集与研究利用[J].南京林学院学报,1985(2):47-52.WANG Zhangrong,CHEN Tianhua,LI Zhangju,et al.Genetic resources exploration and research utilization of Pseudolarix amabilis(Nelson) Rehd.[J].J Nanjing For Coll,1985(2):47-52.
[6]吴征镒.新华本草纲要:第1册[M].上海:上海科学技术出版社,1988.
[7]魏学智,胡玉熹,林金星,等.中国特有植物金钱松的生物学特性及其保护[J].武汉植物学研究,1999,17(增刊):73-77.WEI Xuezhi,HU Yuxi,LIN Jingxing,et al.The biology and concervation of pseudolarix amabilis(Nelson) Rehd.[J].J Wuhan Bot Res,1999,17(supp):73-77.
[8]邵邻相,张凤娟.6种松科植物叶表皮的扫描电镜观察[J].植物研究,2005,25(3):281-285.SHAO Lingxiang,ZHANG Fengjuan.Sem observation on leaf epidermis of 6 species in Pinaceae [J].Bull Bot Res,2005,25(3):281-285.
[9]喻方圆,钱锦.不同采种期金钱松种子品质的研究[J].中南林学院学报,1999,19(4):45-47.YU Fangyuan,QIAN Jin.a seed quality study of Pseudolarix amabilis in different collection periods [J].J Cent South For Univ,1999,19(4):45-47.
[10]钱洪涛,张纪林,宋建法,等.金钱松幼苗移植措施对生长的影响[J].林业科技开发,2005,19(3):59-60.QIAN Hongtao,ZHANG Jilin,SONG Jianfa,et al.The effect on growth of Pseudolarix amabilis by seedling transplant measures [J].China For Sci Technol,2005,19(3):59-60.
[12]刘洪亮,何承忠.金钱松化学成分及生物活性研究现状与展望[J].西南林学院学报,2008,28(1):52-56.LIU Hongliang,HE Chengzhong.Research status and prospect of the chemical constituents and bioactivities of Pseudolarix kameferi[J].J Southwest For Coll,2008,28(1):52-56.
[13] 李林初.金钱松(属)的细胞分类学研究[J].云南植物研究,1994,16(3):248-254.LI Linchu.A cytotaxonomical study on Pseudolarix kameferi[J].Acta Bot Yunnan,1994,16(3):248-254.
[14]吴文珊,张清其,刘剑秋.金钱松染色体核型的研究[J].福建师范大学学报,1996,12(1):81-83.WU Wenshan,ZHANG qingqi,LIU Jianqiu.Karyotype studies of Pseudolarix amabilis [J].J Fujian Teach Univ Nat Sci,1996,12(1):81-83.
[15]何子灿,柯善强,桂耀林,等.外源激素对金钱松胚外植体愈伤组织的诱导及其器官发生的调节作用[J].武汉植物学研究,1995,13(1):81-86.HE Zicang,KE Shanqiang,GUI Yaolin,et al.Influence of plant growth regulators on the in vitro organogenesis of Pseudolarix amabilis embryo explants [J].J Wuhan Bot Res,1995,13(1):81-86.
[16]刘建锋,肖文发.濒危植物崖柏遗传多样性的RAPD分析[J].江西农业大学学报,2008,30(1):68-72.LIU Jianfeng,XIAO Wenfa.RAPD analysis of the genetic diversity of a critically endangered plant,Thuja sutchuenensis(Cupressaceae)[J].Acta Agric Univ Jiangxi,2008,30(1):68-72.
[17]李建辉,金则新,李钧敏.濒危植物长叶榧群体遗传多样性的RAPD分析[J].应用生态学报,2007,18(12):2661-2667.LI Jianhui,JIN Zexin,LI Junmin,et al.Genetic diversity of endangered plant Torreya jackii:a study with RAPD markers [J].Chin J Appl Ecol,2007,18(12):2661-2667.
[18]曹福亮,花吉吉斌,汪贵斌,等.野生银杏资源群体遗传多样性的RAPD分析[J].浙江林学院学报,2008,25(1):22-27 CAO Fuliang,HUA Zhebin,WANG Guibin,et al.Genetic diversity in wild populations of Ginkgo biloba using random amplified polymorphic DNA(RAPD) analysis [J].J Zhejiang For Coll,2008,25(1):22-27.
[19] MOSSCLCR A,EGGER K N,HUGHCS G A.Low levels of genetic diversity in red pine confirmed by random amplitied polymorphic DNA markers [J].Can J For Res,1992,22:1332-1337.
[20] SZMIDT A E,WANG Xiaoru,LU Mengzhu.Empirical assessment of allozyme and RAPD variation in Pinus sylvestris(L.)using haploid tissue analysis [J].Heredity,1996,76:412-420.
[21] HONG Y P,HIPKINS V D,STRAUSS S H.Chloroplast DNA diversity among trees,populations and species in the California closed-cone pines(Pinus radiata,P.miuricata and P.attanuata) [J].Genetics,1993,135:1187-1196.
[22]夏铭,周晓峰,赵士洞.天然红松群体遗传多样性的RAPD分析[J].生态学报,2001,21(5):730-737.XIA Ming,ZHOU Xiaofeng,ZHAO Shidong.RAPD analysis on genetic diversity of natural populations of Pinus koraiensis [J].Acta Ecol Sin,2001,21(5):730-737.
[23]唐娟娟,范义荣,朱睦元.黄山松群体遗传多样性分析[J].浙江林学院学报,2003,20(1):23-26.TANG Juanjuan,FAN Yirong,ZHU Muyuan.Analysis of the genetic diversity of Pinus taiwanensis populations [J].J Zhejiang For Coll,2003,20(1):23-26.
[24]李丹,彭少麟.3个不同海拔梯度马尾松种群的遗传多样性及其与生态因子的相关性[J].生态学报,2001,21(3):415-421.LI Dan,PENG Shaolin.Genetic diversity in three Pinnus massoniana populations in different elevations and its relationship with ecological factors [J].Acta Ecol Sin,2001,21(3):415-421.
[25]高燕会,朱玉球,黄华宏,等.杨梅RAPD-PCR体系的正交优化研究 [J].生物技术,2006,16(3):55-58.GAO Yanhui,ZHU Yuqiu,HUANG Huahong,et al.Study on optimization of RAPD-PCR reaction system of Myrica rubra using orthogonal design [J].Biotechnology,2006,16(3):55-58.
[26]张雷凡,高燕会,朱玉球,等.石蒜属植物种质资源ISSR-PCR反应体系的建立[J].浙江林学院学报,2007,24(2):156-161 ZHANG Leifan,GAOyanhui,ZHU Yuqiu,et al.An inter simple sequence repeats(ISSR) reaction system for Lycoris(Amaryllidaceae) [J].J Zhejiang For Coll,2007,24(2):156-161.
[27]李慧慧,朱玉球,斯金平,等.栝楼ISSR-PCR体系的正交优化[J].生物技术,2009,19(6):38-41.LI Huihui,ZHU Yuqiu,SI Jinping,et al.Establishment of ISSR reaction system of Trichosanthes kirilowii Maxim[J].Biotechnology,2009,19(6):38-41.
[28]殷瑶,谷勇,蔡丽莎,等.浅述生物多样性的价值及其保护[J].生物学通报,2009,44(5):8-11.YIN Yao,GU Yong,CAI Lisha,et al.The value and protection of biodiversity [J].Bull Biol,2009,44(5):8-11.
[29]欧阳志勤,程勤,张兴,等.稀有植物云南金钱槭的现状及保护措施[J].林业调查规划,2007,32(2):143-145.OUYAN Zhiqin,CHENG Qin,ZHANG Xing,et al.Status of rare Dipteroina dyeriana and counter measure for its protection [J].For Invent Plan,2007,32(2):143-145.
[30]薛达元,蒋明康,李正方,等.苏浙皖珍稀濒危植物保护规划的研究[J].自然资源学报,1992,7(1):27-34.XUE Dayuan,JIANG Mingkang,LI Zhengfang,et al.A study of the conservation planning for the rare and risk plant species in Jiangsu,Zhejiang and Anhui Provinces [J].J Nat Resour,1992,7(1):27-34.
[31]陈泓,黄仕训.广西热带稀有濒危植物迁地保护地域探讨[J].广西植物,2006,26(6):670-674.CHEN Hong,HUANG Shixun.Discussion on regions of ex-situ conservation for tropical rare and endangered plants of Guangxi[J].Guihaia,2006,26(6):670-674.
猜你喜欢
今日农业(2022年13期)2022-09-15 01:18:00
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
中国麻业科学(2018年6期)2018-04-09 11:22:12
西南农业学报(2016年5期)2016-05-17 05:42:21
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
西南农业学报(2016年6期)2016-04-16 05:12:47
广西林业科学(2016年3期)2016-03-16 05:43:21
法医学杂志(2015年4期)2016-01-06 12:36:36
中国光学(2015年1期)2015-06-06 18:30:20
河南医学研究(2014年7期)2014-02-27 14:53:42
