
广谱抗真菌转基因水稻植株化学成分与结构动态变化
2011-07-05 03:43:44姚清华陈丽华宋永康黄敏敏李玥仁1福建省农业科学院中心实验室福建省精密仪器农业测试重点实验室福建福州350003
生物安全学报 2011年3期
黄 薇,姚清华,陈丽华,宋永康,黄敏敏,李玥仁1福建省农业科学院中心实验室;福建省精密仪器农业测试重点实验室,福建福州 350003
真菌病害是影响农作物产量和品质的重要因素。近年来,随着分子植物病理学和植物基因工程的迅速发展,利用DNA重组技术和转基因技术,将外源抗真菌基因导入水稻,培育出抗病水稻新品种,成为解决这一问题的有效途径(张志忠等,2005;宋冬明等,2010)。在众多外源抗真菌基因中,具有非特异性的广谱抗真菌蛋白基因因其具有抗性谱宽、可在多种植物中表达等优势,而倍受研究者的重视。
广谱抗真菌蛋白基因在植物组织中的组成型高水平表达,使得转基因水稻获得了较高的抗病水平和较宽的抗性谱,但同时也可能造成潜在的生态风险。此外,真菌在植株木质素的降解中起着主要的作用,组成型高水平表达的广谱抗真菌蛋白有可能抑制真菌对水稻秸秆木质素的降解(郁红艳等,2003)。根据转基因食品安全评估中的实质等同原则(刘晓云和何小维,2005),本文在前期研究的基础上,对广谱抗真菌转基因水稻转品1和转品8的整个生长发育过程中化学成分的变化进行分析,同时利用电子显微镜观察水稻品种的组织显微结构,以期为进一步研究广谱抗真菌转基因水稻秸秆的降解效率以及环境安全性评估提供参考。
1 材料与方法
1.1 供试材料
供试水稻品种为广谱抗真菌转基因水稻转品1、转品8及其非转基因对照七丝软粘。转品1和转品8由中山大学和广东海洋大学联合选育,是利用基因枪法将2~3个广谱抗真菌蛋白基因以及标记基因hpt导入籼稻品种“七丝软粘”中,经过选育获得的对稻瘟病Magnaporthe grisea(Hebert)Barr和纹枯病菌Rhizoctonia solani Kühn抗性显著提高的转基因水稻(冯道荣等,1999)。其中,转品1为三价抗真菌转基因水稻,转入水稻碱性几丁质酶基因(RC24、RCH10)和水稻酸性几丁质酶基因(RAC22);转品8为双价抗真菌转基因水稻,转入目的基因为水稻碱性几丁质酶基因(RC24)和苜蓿葡聚糖酶基因(β-1,3-Glu);启动子为在水稻中高效表达的Act1。
1.2 方法
1.2.1 供试水稻的栽培方法 于2008年3月17日在福建省福州市闽侯县良种场种植供试水稻。供试地貌为闽江两岸丘陵河谷区,属南亚热带季风气候。选择水肥条件一致的隔离试验水田,试验水田为梯田形式,面积约为500 m2,分为9个试验小区。水田土壤为黄红壤,总碳含量 16.44 g·kg-1,总氮 1.15 g·kg-1,有效磷 33.31 mg·kg-1,速效钾68.97mg·kg-1,pH 5.56。田间水肥、栽培技术管理按当地栽培水平实施,每个品种种植3个重复小区。水稻测试样品均以多点随机的方法分别于分蘖期、孕穗期、抽穗期、灌浆期、成熟期进行采样。将采回的样品叶片、叶鞘和茎分开,在105℃下杀青15 min,然后在80℃烘24 h,粉碎后过60目筛,装入样品袋存放于干燥器内,以备分析待用。
1.2.2 化学成分测定 秸秆化学成分含量以干物质为比较基础,依据van Soest et al.(1991)的方法,采用FOSS纤维浸提器测定秸秆各植物学部位(叶片、叶鞘、茎)半纤维素、纤维素和木质素的含量。依据GB/T 6432-1994,采用FOSS全自动凯氏定氮仪测定秸秆叶片、叶鞘、茎的粗蛋白质含量。每个样品设3个重复。
1.2.3 组织显微结构观察 取成熟期水稻,经液氮干燥后,将叶片上表皮和茎秆横断面固定在样品铜台上,真空离子溅射仪喷金包埋,于日本电子JSM-6380LU扫描电镜下进行观察、拍照。
1.2.4 数据处理 采用 Excel 2003软件进行数据的基本处理,对不同生育期各水稻品种的化学组分采用SPSS软件包中的方差分析过程(ANOVA)进行比较。
2 结果与分析
2.1 广谱抗真菌转基因水稻纤维素含量分析
广谱抗真菌转基因水稻转品1和转品8与相应非转基因水稻七丝软粘在不同生育期叶片、叶鞘和茎的纤维素含量动态变化趋势见图1。从图中可知,七丝软粘叶片、叶鞘和茎的纤维素含量在不同生育期内均呈现明显的变化,且各部位的变化趋势大致相似,即从分蘖期至孕穗期逐渐下降,至抽穗期逐渐增加,至灌浆期逐渐下降,种子灌浆后纤维化程度增加。转品1和转品8的变化趋势与七丝软粘一致,且含量差异均未达到显著水平(P>0.05)。
2.2 广谱抗真菌转基因水稻半纤维素含量分析
图2显示为广谱抗真菌转基因水稻转品1、转品8以及相应非转基因水稻七丝软粘在不同生育期叶片、叶鞘和茎半纤维素含量的变化趋势。从图中可以看出,供试水稻品种的叶片、叶鞘和茎的半纤维素含量变化趋势基本一致;且除个别情况外,不同生育期品种间各个部位半纤维素含量的差异不显著(P>0.05)。此外,我们还发现,供试品种半纤维素的动态变化与纤维素成负相关,当纤维素含量增加时,半纤维素含量下降。由此可见,水稻植株生理上具有一定程度的自主调节能力,纤维素与半纤维素的协同作用,能够维持对植物体的机械支撑。此现象同 Wang et al.(2006)和沈恒胜等(2004)的研究结果基本吻合。
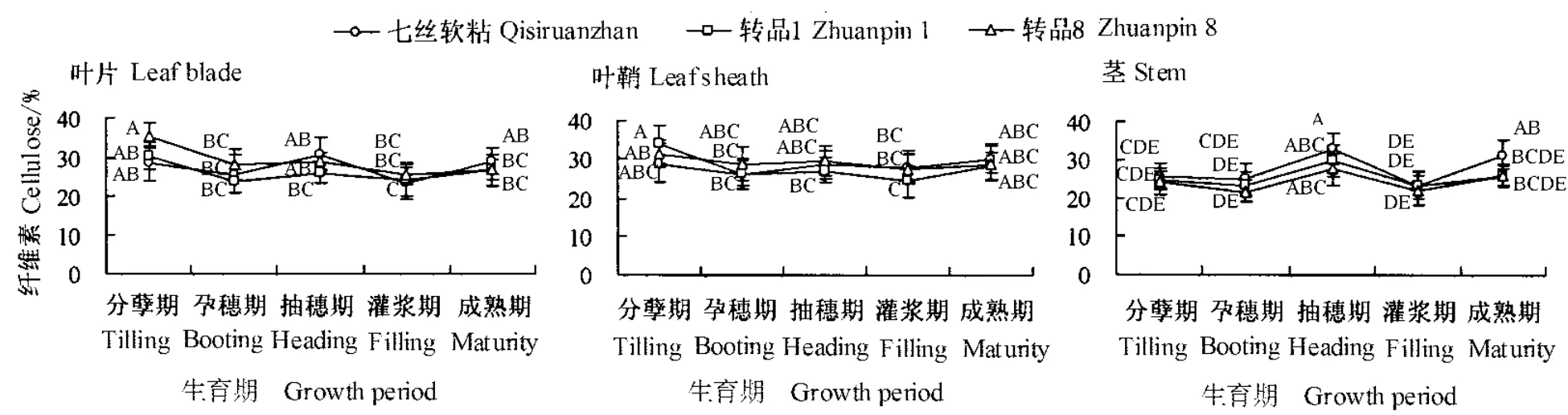
图1 广谱抗真菌转基因水稻生长发育过程中纤维素含量动态变化Fig.1 Dynamic changes in cellulose content in transgenic rice plants expressing broad-spectrum antifungal proteins at their different growth and developmental stages
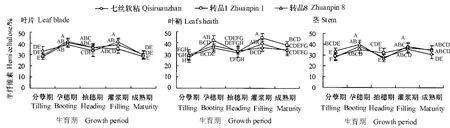
图2 广谱抗真菌转基因水稻生长发育过程中半纤维素含量动态变化Fig.2 Dynamic changes in hemi-cellulose content in transgenic rice plants expressing broad-spectrum antifungal proteins at their different growth and developmental stages
2.3 广谱抗真菌转基因水稻木质素含量分析
广谱抗真菌转基因水稻转品1、转品8及其相应非转基因水稻七丝软粘在进入抽穗期后,其叶片、叶鞘和茎的木质素含量均有不同程度的上升趋势,且各个生育期木质素含量都表现为叶片、叶鞘>茎(图3)。沈恒胜等(2001)认为,稻草中木质素含量同硅化物的含量具有显著相关性,而水稻硅化物分布又具有随生长发育向外、向上传输的特性,故相对于叶片和叶鞘,茎的木质素含量较低。
木质素的降解是真菌、细菌及相应微生物群落共同作用的结果,其中真菌起主要作用。分析结果表明,除孕穗期的叶鞘及成熟期的茎外,供试广谱抗真菌转基因水稻各部位的木质素含量与相应非转基因水稻相比均不存在显著差异(P>0.05)。由此可推测,广谱抗真菌蛋白基因的导入对秸秆木质素的含量和分布没有较大的影响。
2.4 广谱抗真菌转基因水稻粗蛋白质含量分析
从图4可知,七丝软粘叶片、叶鞘和茎的粗蛋白质含量均在分蘖期最高,分别为23.4%、10.4%和17.1%;之后逐步下降,成熟期达到最低,分别为11.8%、5.8%和 3.0%。转品 1、转品 8 与七丝软粘的叶片、叶鞘和茎粗蛋白质含量变化趋势一致。水稻在营养生长和生殖生长过程中都需要大量的蛋白质来满足细胞组织的更新和修补,这可能是导致粗蛋白质含量持续降低的原因。对试验数据进行差异显著性检验表明,转品1和转品8与七丝软粘的蛋白质含量差异不显著(P>0.05)。这在一定程度上反映了广谱抗真菌转基因水稻的营养品质与其相应非转基因品种具有相似性。
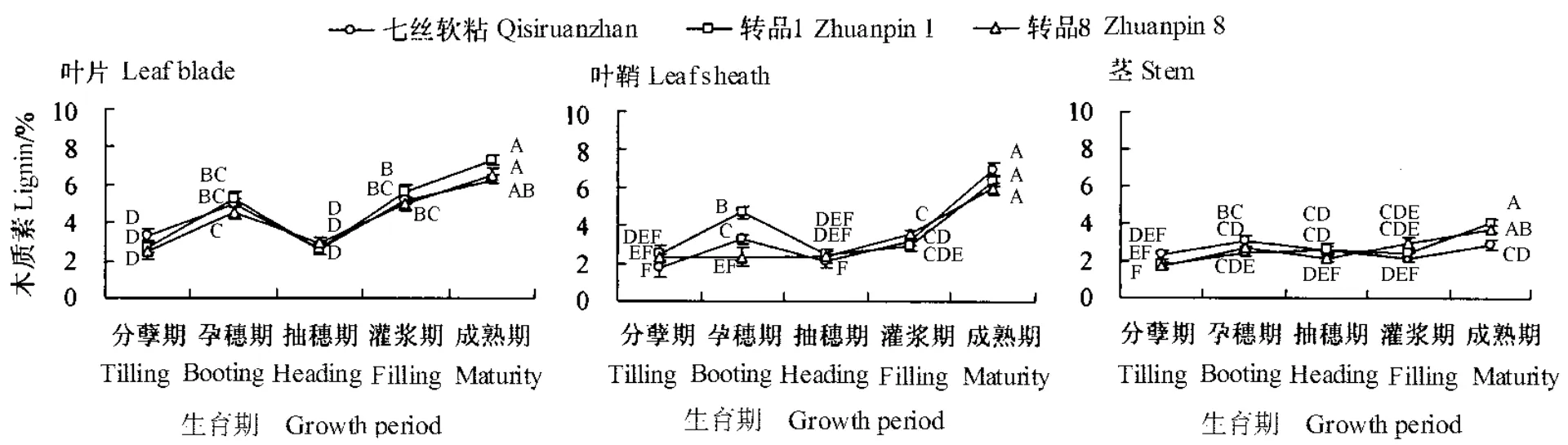
图3 广谱抗真菌转基因水稻生长发育过程中木质素含量动态变化Fig.3 Dynamic changes in lignin content in transgenic rice plants expressing broad-spectrum antifungal proteins at their different growth and developmental stages
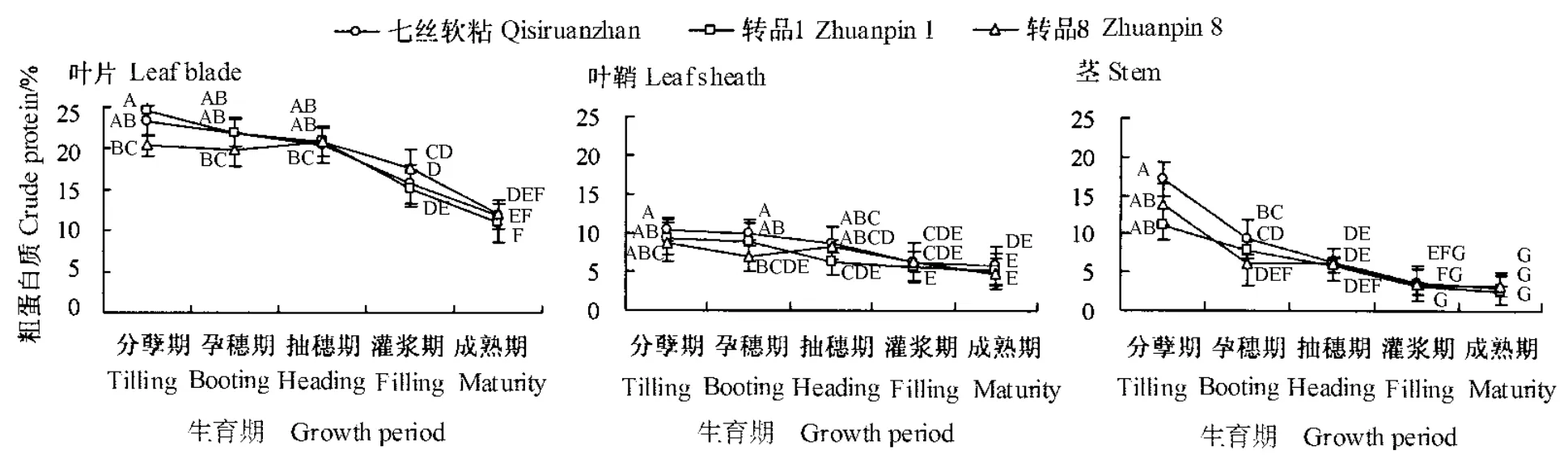
图4 广谱抗真菌转基因水稻生长发育过程中粗蛋白质含量动态变化Fig.4 Dynamic changes of crude protein content during the growth and development in transgenic rice plants expressing broad-spectrum antifungal proteins plants at different growth stages
2.5 广谱抗真菌转基因水稻组织显微结构观察
供试水稻品种的叶片表面均含有许多大小不一的硅质瘤状结构,这些硅质瘤状结构分布在叶片不同的部位,包括气孔的保卫细胞上(图5A~C)。分析发现,转品1、转品8及七丝软粘的硅质瘤状结构排列形状变化趋向一致,致密程度无明显差异。此外,各品种间叶表皮的气孔排列致密程度接近,每个气孔周围均分布了4~8个呈簇状排列、尖端向心性的硅质瘤状结构。
用扫描电镜观察茎秆组织(图5D~F)显示,广谱抗真菌转基因水稻与相应非转基因水稻茎秆的形态结构基本相似,茎壁、厚壁组织、薄壁组织以及维管束等结构的形态和所占面积比例基本一致,大、小维管束的数目、分布密度及面积大小等也无明显差异,且各水稻品种茎秆的薄壁组织细胞中均贮藏着丰富的木质素以及淀粉等多糖颗粒。由此可见,广谱抗真菌基因的转入对水稻的组织形态没有影响。
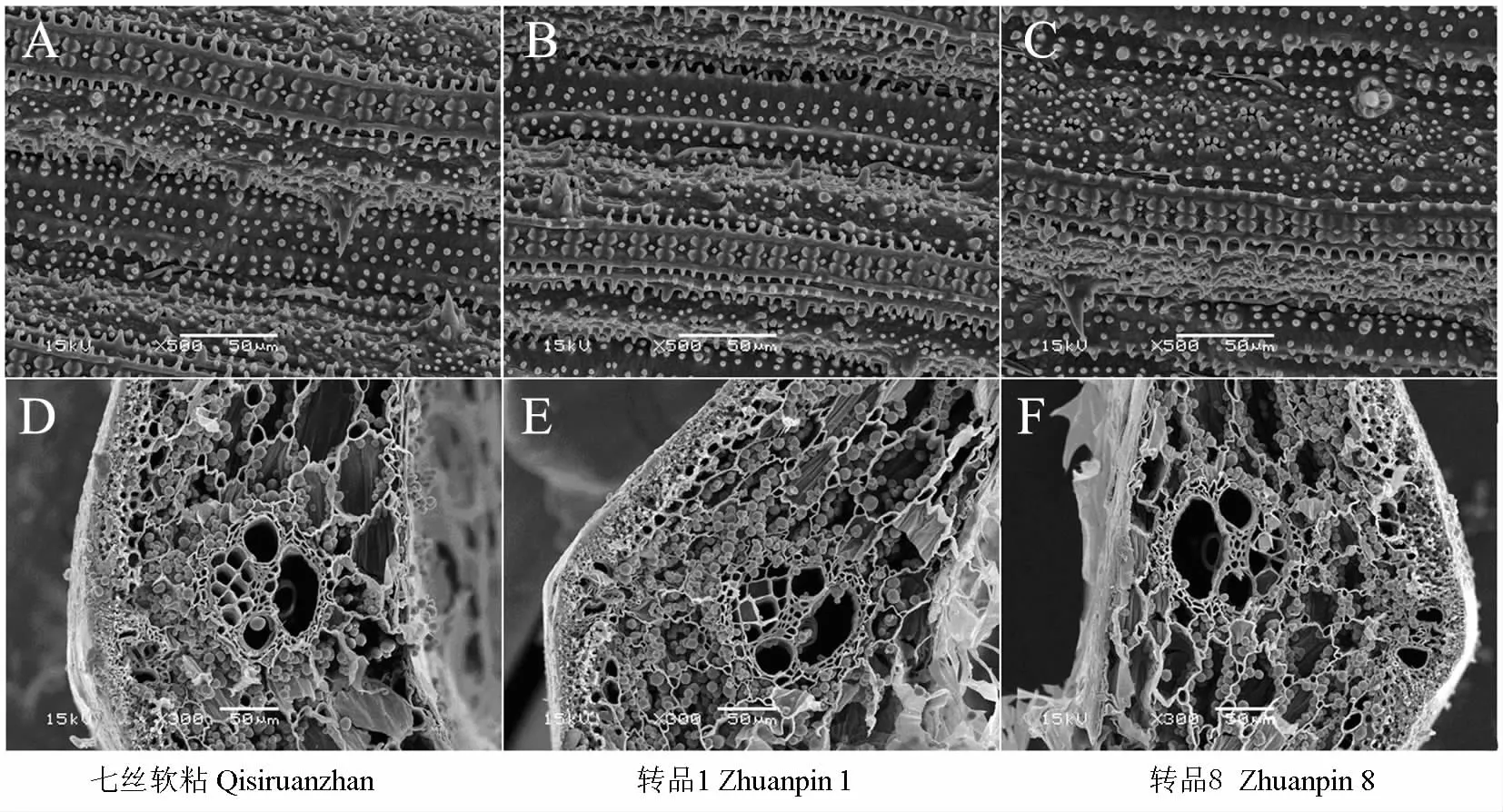
图5 广谱抗真菌转基因水稻与非转基因对照的组织显微结构Fig.5 The microstructure of leaf and stem tissue of transgenic rice plants expressing broad-spectrum antifungal proteins plants and its nontransgenic counterpart under scanning electron microscope(SEM)
3 讨论
广谱抗真菌转基因水稻尽管对真菌病害具有明显抗性,可以大幅度提高水稻产量,但人们对转基因产品的食用安全性及其对生态环境的影响存在疑虑。将转基因作物及来源的食品与非转基因同源对照物在表型、农艺性状、组成成分等方面进行详细的比较,从而得出转基因作物与非转基因同源对照物是否具有实质等同性结论的工作被认为是转基因作物安全性评价的重要起始点(贺晓云等,2008)。李英华等(2004)对转入抗水稻白叶枯病Xanthomonas oryzae pv.oryzae Dye Xa21基因的转基因大米进行营养成分分析时,发现当转基因水稻与非转基因水稻种植条件完全相同时,营养成分无差异。Han et al.(2005)对转入豇豆胰蛋白酶抑制剂基因的水稻及其亲本的主要营养成分(水分、蛋白质、氨基酸、脂肪、碳水化合物、纤维、灰分、各类脂肪酸、维生素和矿物质等)进行了详细的分析和比较,结果显示除脂肪和脂肪酸有细微的区别外,其他成分均无差异。Li et al.(2007)同样对转sck和cry1AC基因的抗虫水稻与非转基因对照进行了细致的营养成分等同性分析,结果认为转基因水稻与对照在主要营养成分上具有实质等同性。但是,这些分析采用的都是稻米的数据,仅强调了食用安全的评价,而忽视了转基因水稻对环境的安全性评价。
本研究对广谱抗真菌转基因水稻转品1和转品8及其相应非转基因水稻七丝软粘在整个生长发育过程中各植物学部位(叶片、叶鞘、茎)的主要化学成分,包括半纤维素、纤维素、木质素以及粗蛋白质的含量进行了测定和分析。结果显示,在整个生长发育过程中,转品1和转品8各植物学部位的化学成分含量变化趋势与对照七丝软粘基本一致。水稻叶面的“硅质化”特殊结构以及茎秆的粗细,大、小维管束数目和茎壁厚度等物理特性都已被证明与水稻抵抗病菌和害虫侵害的能力有着密切的关系(杨秉耀等,2006;Belanger et al.,1995;Fidanza& Dernoeden,1996;张上都等,2010)。故本研究利用扫描电镜观察了成熟期转品1、转品8及对照七丝软粘的组织显微结构,结果发现广谱抗真菌转基因水稻的组织形态结构未发生明显变化,与其相应非转基因水稻基本一致。因此,认为广谱抗真菌几丁质酶基因和葡聚糖酶基因的导入未引起水稻植株化学成分组成和组织显微结构的明显变化。据报道,秸秆的化学成分组成与秸秆的降解消化有密切的相关性(Keys&DeBarthe,1974)。
转基因水稻安全性评价工作是复杂的系统性工程。本研究仅是转基因水稻环境安全性评价的起步,有关其食用安全性以及对生态环境的影响等还有待于进一步探索试验。
贺晓云,黄昆仑,秦伟,罗云波.2008.转基因水稻食用安全性评价国内外概况.食品科学,29(12):760-765.
李英华,朴建华,陈小萍,卓勤,毛德倩,杨丽琛,杨晓光.2004.Xa21转基因大米的营养学评价.卫生研究,33(3):303-306.
冯道荣,许新萍,卫剑文,李宝健,杨祁云,朱小源.1999.使用双抗真菌蛋白基因提高水稻抗病性的研究.植物学报,1999,41(11):1187 -1191.
刘晓云,何小维.2005.转基因食品安全评估.食品工业科技,(3):20-22.
沈恒胜,陈君琛,曾大力,涂杰峰,汤葆莎,滕胜.2004.低纤维基因突变水稻稻草细胞壁组分动态发育分析.中国农业科学,37(7):943-947.
沈恒胜,陈君琛,倪德斌.2001.稻草硅化和溶解特性对稻草纤维降解率及其利用的影响.中国农业科学,34(6):672-678.
宋冬明,贺梅,孟昭河,李春光,张景龙,程芳艳.2010.转基因水稻的研究进展.北方水稻,(3):67-80.
杨秉耀,陈新芳,刘向东,郭海斌.2006.水稻不同品种叶表面硅质细胞的扫描电镜观察.电子显微学报,25(2):146-149.
郁红艳,曾光明,胡天觉,陈耀宁.2003.真菌降解木质素研究进展及在好氧堆肥中的研究展望.中国生物工程杂志,23(10):57-61.
张上都,余显权,赵福胜,马天进.2010.水稻脆秆重组体茎秆的形态结构观察与理化测定.西南农业学报,23(1):1-4.
张志忠,吴菁华,吕柳新,林义章.2005.植物几丁质酶及其应用研究进展.福建农林大学学报:自然科学版,34(4):494-499.
Belanger R R,Bowen P A,Ehret D L and Menzies J G.1995.Soluble silicon its role in crop and disease management of greenhouse crops.Plant Disease,79:329 -335.
Fidanza M A and Dernoeden P H.1996.Interaction of nitrogen source application timing and fungicide on Rhizoctonia blight in ryegrass.Hort Science,31:389 -392.
Han J,Yang Y,Chen S,Wang Z,Yang X,Wang G and Men J.2005.Comparison of nutrient composition of parental rice and rice genetically modified with cowpea trypsin inhibitor in China.Journal of Food Composition and Analysis,18:297 -302.
Keys J E and DeBarthe J V.1974.Cellulose and hemicellulose digestibility in the stomach,small intestine and large intestine of swine.Journal of Animal Science,39:53 -56.
Li X,Huang K,He X,Liang Z,Li H and Lou Y.2007.Comparison of nutritional quality between Chinese indica rice with sck and cry1AC genes and its nontransgenic counterpart.Journal of Food Science,72:420 -424.
van Soest P J,Robertson J B and Lewis B A.1991.Method for dietary fiber,neutral detergent fiber,and nonstarch polysaccharides in relation to animal nutrition.Journal of Dairy Sciences,74:3583 -3597.
Wang H F,Wu Y M,Liu J X and Qian Q.2006.Morphological fractions,chemical compositions and in vitro gas production of rice straw from wild and brittle culm1 variety harvested at different growth stages.Animal Feed Science and Technology,129:159 -171.
猜你喜欢
作物杂志(2023年3期)2023-08-07 01:04:08
三农资讯半月报(2020年6期)2020-04-10 11:01:12
上海包装(2019年8期)2019-11-11 12:16:14
天津造纸(2016年1期)2017-01-15 14:03:29
植物保护(2016年1期)2016-09-14 06:14:52
中国造纸学报(2015年1期)2015-12-16 19:35:25
中国塑料(2014年4期)2014-10-17 03:00:45
湖南农业(2014年9期)2014-04-02 07:03:37
中国信息化·学术版(2013年5期)2013-10-09 06:02:58
华南农业大学学报(2013年4期)2013-08-29 09:30:28

- 生物安全学报的其它文章
- 用超微细胞化学定位技术揭示ATP酶在紫茎泽兰高温适应性中的作用