
滩地人工杨树林土壤呼吸变化规律与环境因子的关系研究
2011-06-21 10:14周金星
水土保持研究 2011年6期
马 涛,周金星
(1.甘肃省水土保持科学研究所,兰州 730020;2.中国林业科学研究院 荒漠化研究所,北京 100091)
滩地人工杨树林土壤呼吸变化规律与环境因子的关系研究
马 涛1,周金星2
(1.甘肃省水土保持科学研究所,兰州 730020;2.中国林业科学研究院 荒漠化研究所,北京 100091)
为揭示滩地人工杨树林土壤呼吸特征及其与环境因子的关系,采用红外气体分析(IRGA)法对滩地人工杨树林土壤CO2释放量进行测定,分析温度、水分、林下植被结构等因素对林地土壤呼吸日变化动态的影响。结果表明:滩地人工杨树林生态系统土壤呼吸日动态变化呈单峰曲线,在11:00-13:00达到最大值。土壤呼吸速率大小的排列顺序为:2006年6月>2005年10月>2006年3月>2006年10月。日均土壤呼吸速率为2.29±0.75μmol/(m2·s)(P<0.05,n=13),变异系数为54.40%。土壤呼吸对地表温度的敏感性大于对地下5cm处温度的敏感性。土壤呼吸温度敏感性指标Q10值为1.28±0.79(P<0.05,n=13),低于不包括湿地在内的全球尺度水平。土壤水分与土壤呼吸相关性差,不是滩地林地土壤CO2释放量的限制因子。林下植被亦是影响土壤呼吸速率的一个重要因素。
人工杨树林;土壤呼吸;环境因子;Q10值;滩地
全球土壤有机碳库的总量为1 200~1 400Pg,占整个陆地生态系统有机碳库的2/3,约为大气碳库的两倍[1]。土壤呼吸是陆地生态系统第二大碳通量,陆地生态系统的总呼吸量中有50%~75%来源于土壤呼吸[2]。森林生态系统是陆地生态系统中最重要的组成部分,具有较大的CO2负通量,是一个重要的C库[3]。而森林土壤碳亦是全球碳库的重要组成部分,占全球土壤碳的73%,在全球碳循环方面发挥重要作用[4]。森林地表释放CO2是森林生态系统碳循环的一个重要环节,为森林碳支出的主要方式[5],也是大气CO2重要的来源[6]。在目前已建立的长期监测CO2通量网站中,森林土壤呼吸已成为重要的监测项目之一[7]。因此对森林地表CO2通量进行长期连续观测和对其影响因子的研究是认识整个森林生态系统碳平衡的重要组成部分。
自20世纪70年代,国外就有关于森林土壤呼吸研究的报道,而国内在这方面研究起步相对较晚,到目前为止,国内对森林土壤呼吸的研究主要分布在北京山地温带、暖温带林区[8-10]、长白山阔叶林区[11-13]、鼎湖山丘陵阔叶混交林区[14-15]、西双版纳及海南岛尖峰岭热带雨林区[16-18],同时在六盘山天然次生林区[19]以及在亚热带喀斯特森林[20]也有土壤呼吸的相关报道。在湿地生态系统仅对草甸群系土壤呼吸进行过研究[21-23]。
本研究以湖南、安徽两处滩地人工杨树林为研究对象,采用红外气体分析法,测定土壤呼吸昼夜变化规律,揭示影响土壤呼吸作用的环境因子及其内在机制,为深入研究滩地杨树人工林土壤碳动态以及估算该地区的碳收支提供参考。
1 材料与方法
1.1 试验地概况
滩地是平原河床季节性淹水的微地形。长江中下游滩地分布于亚热带季风气候区,生物资源、气候和土壤资源丰富,是我国经济相对发达地区。长江中下游沿江约有60万hm2滩地[24]。20世纪80年代初开始试栽杨树,开发并利用滩地土地资源,为当地经济发展和生态环境的改善起到巨大作用。选择湖南、安徽长江外滩人工杨树林地进行试验研究。试验地基本情况见表1。

表1 试验地概况
1.2 研究方法
1.2.1 试验设计 2005年9月在湖南、安徽两滩地杨树林地内分别随机布置2个3m×3m的样方,在每个样方内随机布置6个聚乙烯圆柱体测量室,其中长期测量室1个,短期测量室5个,样点分布为正五边形,短期测量室置于正五边形的顶点,长期测量室置于内切圆圆心。每个试验点共布置12个测量室。长期测量室的配套基座为直径230mm,高114.3mm;短期测量室的基座直径100mm,高44mm,嵌入土壤深度为25~35mm,用于定期测定土壤呼吸速率。
选择2005年10月、2006年3月、2006年6月和2006年10月中每月上旬和下旬晴朗天气各1d,用长、短期测量室同步进行土壤呼吸速率日变化测定,取其平均值说明各月份滩地人工杨树林土壤呼吸速率日变化特征。
1.2.2 土壤呼吸速率的测定 土壤呼吸日动态变化采用红外气体分析仪(IRGA)测定,仪器型号为LI-8100(由美国LI-COR公司生产,基因有限公司提供)。该仪器的测量原理、测量过程及主要特点见文献[25]。测定前,将基座内的草本植物齐地剪除,并将枯枝落叶清除,但尽量做到不破坏土壤。砸实基座外土壤,以防漏气。为避免由于安置气室对土壤扰动造成的短期内呼吸速率的波动,经过24h平衡后进行测定。土壤呼吸速率进行24h连续测定,9:00-19:00每2h测定1次,19:00至次日7:00每3h测定1次,共计10次。土壤CO2排放通量的计算公式为:
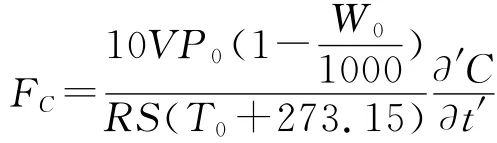
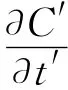
1.2.3 环境因子的测定 在土壤呼吸速率测定的同时,用连接在LI-8100仪器辅助传感器热电偶上的温度传感器和水分传感器同步进行地表温度、土壤温度(5cm)及土壤含水量(5cm)的测定。
1.2.4 林下植被调查 2006年5月,在湖南、安徽试验地随机建立48个1m×1m的样方,记录植物的株数、高度、盖度等因子,分析两地优势植物群落。
2 结果与分析
2.1 滩地人工杨树林土壤呼吸速率变化
土壤呼吸是森林生态系统呼吸最重要的组成部分,受环境因素的影响,包括温度、土壤湿度、根系呼吸量以及枯枝落叶等。这些环境因素的变化同时也反映了植物物候期的变化[26]。由于全球土壤碳库所储存的碳量是大气储碳量的两倍[1],因此土壤呼吸的微小变化都能对大气碳库产生强烈影响。
考虑到温度日变化较小,则土壤呼吸速率日变化幅度不大[18]的特点。为了更好地进行土壤呼吸与环境因子的关系研究,并能更好地分析土壤呼吸速率日变化动态规律,选择在晴朗的天气进行土壤呼吸速率测定,以期更好地反映土壤呼吸速率的日变化规律。土壤呼吸日变化特征如图1所示。
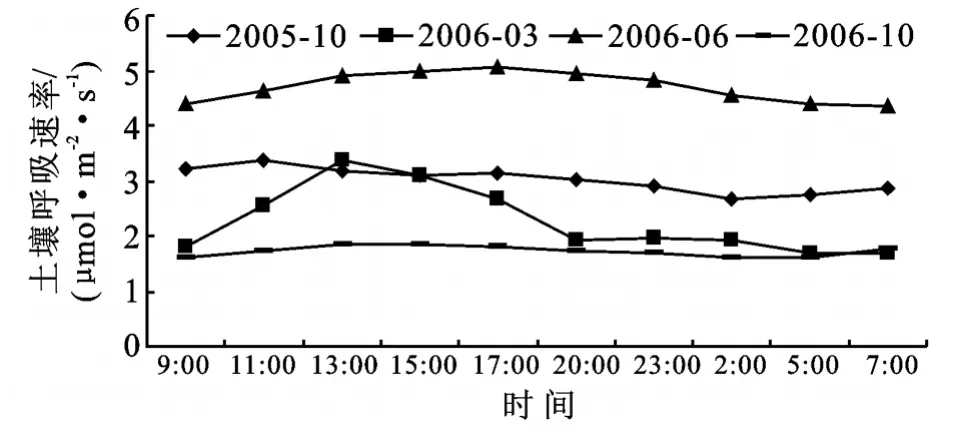
图1 滩地人工杨树林土壤呼吸速率日变化特征
从趋势线的变化规律看,滩地人工杨树林土壤呼吸速率日变化动态表现为单峰曲线。11:00-13:00土壤呼吸作用最强,凌晨2:00-5:00土壤呼吸作用最弱,7:00土壤呼吸速率出现缓慢上升趋势。2006年6月土壤呼吸速率日变化最大值出现时间略晚(17:00),原因为夏季日辐射能量高,日照时数长,植物进行光合作用的时间相对较长,因此,土壤呼吸速率日变化最大值出现的时间相对滞后。从图1可知,2006年6月土壤呼吸速率最大,2006年10月土壤呼吸速率最小,究其原因,在一个生长季节内,环境因子的变化,比如温度的变化必将引起土壤水分、植物第一性生产力、土壤碳储量等因子的同步变化,而这些因子引起的土壤呼吸速率的变化则是随着季节的变化而变化的。夏季土壤温度高,土壤含水量很可能成为土壤呼吸的限制因子[27];冬季大气温度达到一年中最低值,土壤温度反而有可能成为土壤呼吸的限制因子[28]。本研究选择滩地生态系统,冬陆夏水的自然特征,使土壤水分不会成为土壤呼吸的限制因子,因此在植物生长旺季的夏季,土壤呼吸速率显著高于其它各季节;同样,在秋冬季节,植物处于生长末期或休眠期,土壤温度较低,土壤呼吸速率处于一年中的最低值;春季温度回升,植物及土壤生物开始活动,土壤呼吸速率亦呈现上升趋势。从图1看出,2005年10月土壤呼吸速率明显大于2006年10月,原因为2005年7-9月滩地处于淹水期,而2006年滩地未上水,土壤含水量等因子在2005年较2006年优越,因此,土壤呼吸速率值较大。
2.2 林下植被对土壤呼吸速率的影响
植物群落不同,所引起的温度、湿度、土壤有机质含量和pH值等生态环境因子各不相同,因而土壤呼吸的强度也不同[29]。分别对湖南林地和安徽林地5个和8个优势植物群落进行土壤CO2呼吸速率的测定。每个优势种随机设置3个平行试验,取平均值研究植物群落对土壤呼吸速率的影响。优势植物群落情况见表2。

表2 滩地人工杨树林林地优势植物群落名录
不同植物群落土壤呼吸作用的强弱不同(图2),在湖南人工杨树林地,按土壤呼吸速率的大小排列:狗牙根+牛鞭草>益母草>水芹>狗牙根+野胡萝卜>牛鞭草。其日均土壤呼吸速率分别为:5.22,4.33,2.43,1.98,1.21μmol/(m2·s)。取平均值为3.03μmol/(m2·s),变异系数为55.30%。在狗牙根+牛鞭草群落的土壤呼吸日变化中,呼吸速率最大值5.74μmol/(m2·s),最小值4.79μmol/(m2·s),差值0.95μmol/(m2·s)。益母草群落呼吸速率最大值4.79μmol/(m2·s),最小值3.84μmol/(m2·s),差值0.95μmol/(m2·s)。水芹群落呼吸速率最大值3.10μmol/(m2·s),最小值1.88μmol/(m2·s),差值1.22μmol/(m2·s)。狗牙根+野胡萝卜群落呼吸速率最大值2.28μmol/(m2·s),最小值1.71μmol/(m2·s),差值0.57μmol/(m2·s)。牛鞭草群落呼吸速率最大值1.36μmol/(m2·s),最小值0.88μmol/(m2·s),差值0.48μmol/(m2·s)。可知在此林地,土壤呼吸速率日变幅为0.48~1.23μmol/(m2·s)。
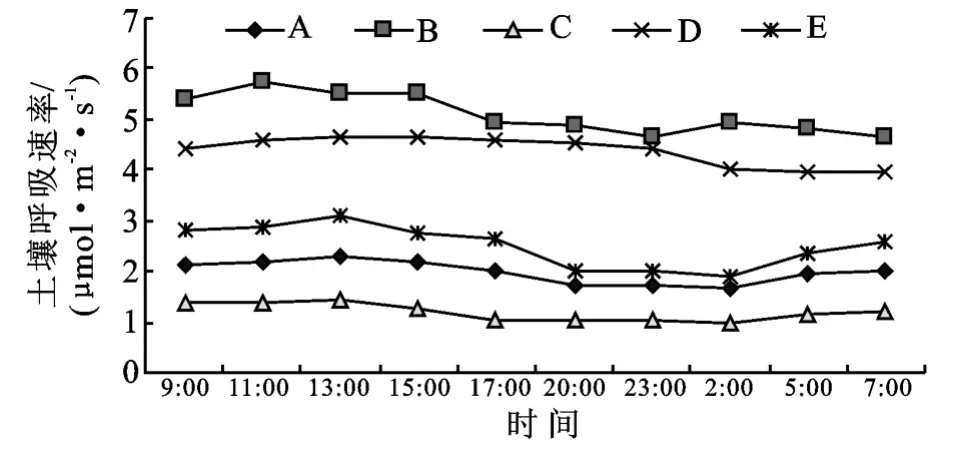
图2 湖南林地不同植被结构土壤呼吸速率日变化
安徽林地不同植物群落土壤呼吸速率日变化曲线变化幅度较湖南林地大(图3)。这是因为安徽造林时间比湖南造林时间早11a,造林密度比湖南林地小。因此由次生演替及人为作用所造成的林下植被结构较湖南林地复杂,因此变化幅度更大一些。从图3中可知,早熟禾+南荻群落、水蓼+狗牙根群落土壤呼吸速率最大,以下依次为扬子毛茛+益母草群落、南荻+狗牙根群落。牛鞭草群落、狗牙根+早熟禾群落以及土牛膝+早熟禾群落土壤呼吸速率日变化趋势基本一致。且均大于苔草+鸡矢藤群落的土壤呼吸速率。按从大到小的顺序,各植物群落日均土壤呼吸速率分别为:2.72,2.59,2.19,1.84,1.46,1.44,1.30,0.99μmol/(m2·s)。取平均值为1.82μmol/(m2·s),变异系数为34.66%。呼吸速率的日变化幅度为0.64~1.68μmol/(m2·s)。
综合以上分析,植被结构的不同使得土壤呼吸速率日变化存在明显差异。将两林地综合分析,滩地人工杨树林土壤呼吸日变幅为:0.48~1.68μmol/(m2·s)。日均土壤呼吸速率0.99~5.22μmol/(m2·s)。取平均值为(2.29±0.75)μmol/(m2·s)(P<0.05,n=13),变异系数为54.40%。说明日均土壤呼吸速率在空间上存在很大的差异性,这也证明了植被结构是影响土壤呼吸速率的一个因子。与其它地域森林生态系统土壤呼吸速率相比,该生态系统日均土壤呼吸速率低于北京山区辽东栎林地[(5.92±1.32)μmol/(m2·s)1][9]及海南岛尖峰岭热带山地雨林10.69μmol/(m2·s)[17],高于长白山林地205.85~395.69μmol/(m2·s)[11],与鼎湖山针阔混交林地488.99~700.57μmol/(m2·s)[15]相当。
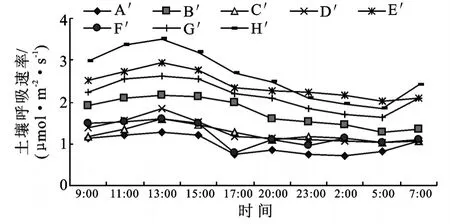
图3 安徽林地不同植被结构土壤呼吸速率日变化
2.3 土壤呼吸速率与温度的关系
湖南林地土壤呼吸速率与地表温度的相关性(R2=0.6773)比其与地下5cm处温度的相关性(R2=0.1523)高(图4)。指数方程可以较好地说明这一点。当温度较低时,土壤呼吸散点聚集在拟合曲线附近,随着温度升高,散点偏离加大。究其原因,该林地有机物质的来源主要是以凋落物和放牧牲畜粪肥的形式集中于地表,土体中的有机质含量相对较低。有机物质在微生物的作用下分解最终产生CO2,且微生物的活性和一系列生化反应的速率都与温度呈明显的正相关[30],而此时土壤水分含量的日变化很小或者说基本稳定,从而使温度成为影响CO2排放的关键因子。同时5cm地温受外部环境影响较小,温度变化较为和缓,CO2排放通量对其响应不甚明显。故CO2排放日变化与地表温度的相关性较与土壤温度的相关性好。

图4 湖南林地土壤呼吸速率与地表温度和地下5cm温度之间的关系
安徽林地土壤呼吸速率与地下5cm处温度的相关性比地表温度的相关性高(图5)。和湖南林地一样,当温度较低时,所有群落土壤呼吸的散点聚集在拟合曲线附近,温度升高,则散点偏离加大。土壤呼吸速率与地下5cm处温度的相关性较差,主要原因亦为5cm地温受外部环境影响较小,温度变化幅度小。从图5中可知,5cm处地温的日变化幅度小(均在5℃之内),从而致使CO2排放通量对其响应不明显;且乔、灌、草3层植被的合理垂直分布,有效地减小了因林下光斑和风等因子引起的地表温度的变异,能够较准确地说明随时间的推移,林内温度的变化情况。故CO2排放日变化与地表温度的相关性较与土壤温度的相关性好。
土壤呼吸的Q10值是衡量土壤呼吸对温度变化响应敏感程度的一个方便指数,即温度每升高10℃,土壤呼吸速率增加的倍数。按统计分析中小样本抽样分析的方法算得两林地土壤呼吸Q10值的置信区间为:1.28±0.79(P<0.05,n=13)。该值大于北京山区辽东栎林土壤呼吸Q10值(1.51)[9],与西双版纳热带季节雨林土壤呼吸Q10值(2.03~2.36)[18]相当,比长白山林地土壤呼吸Q10值(2.44)[11]和鼎湖山森林土壤呼吸Q10值(1.86~3.24)[5]低。其Q10值偏低的原因主要是由于长江中下游滩地冬陆夏水的特殊自然特征,使滩地生态系统大气相对湿度RH值偏高(70%~90%),同时土壤水分含量也保持在较高水平(35%左右),致使林地土壤结构性较差,即土壤固、气、液比例失调,从而制约了土壤呼吸强度的温度敏感性。
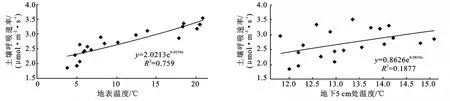
图5 安徽林地土壤呼吸速率与地表温度和地下5cm温度之间的关系
2.4 土壤呼吸速率与土壤水分含量的关系
当土壤湿度降低到永久萎蔫点以下或超过田间持水量的情况时,土壤表面CO2排放量会降低。土壤水分对土壤呼吸的影响主要是通过对植物和微生物的生理活动、微生物的能量供应和体内再分配、土壤的通透性及气体扩散等的调节与控制实现[31]。本研究采用线性方程分析,得出结果是本试验区滩地人工杨树林土壤呼吸速率与地下5cm处水分含量相关性差(图6)。这与Bridge B J.和 Mathes K.的研究结果[32-33]相同。究其原因:(1)可能是由于对土壤含水量的测定数据比较少,无法精确模拟出其与土壤CO2通量的关系;(2)土壤水分含量的高低对土壤孔隙通透性有很大影响,过高的土壤含水量使土壤处于嫌气状态,植物根系和好氧微生物的活动受到抑制,土壤有机质的分解速率降低,土壤中产生的CO2减少。由图6可知,两林地5cm处土壤含水量高(30%~40%),土壤孔隙通透性差,制约了土壤CO2气体的排放,使得土壤呼吸速率对含水量的变化极不敏感,同时也是本地区土壤呼吸速率较热带、亚热带森林地区低很多的原因。
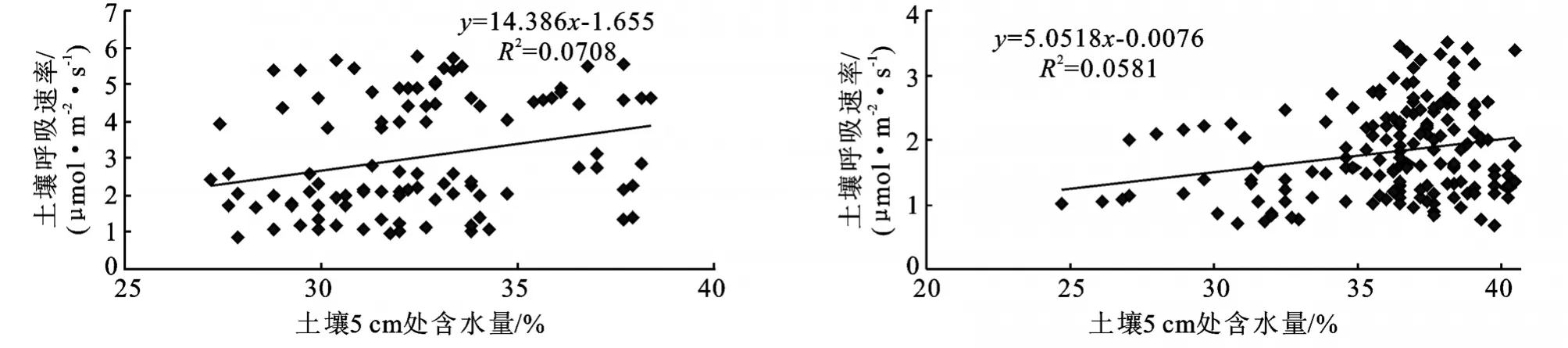
图6 林地土壤呼吸速率与土壤5cm处含水量之间的关系
3 讨论
植物群落在短期或周期性的气温或水分变动的影响下,出现逐年或年际的变化。随季节的变化,水、肥、气、热等环境因子也随之变化,使得植物群落的物候相也相应发生改变。土壤呼吸速率随植物物候相及环境因子的改变而变化。本试验林地土壤呼吸速率随植物物候期的变化而变化的特征就能较好地说明这一点。当然,影响土壤呼吸速率的因子很多,本文仅就温度和水分因子进行了重点分析。在今后对土壤呼吸速率变化特征进行研究时,应从大气-土壤-植被系统着手,综合考虑各生态因子对土壤呼吸速率的影响,以期能够很好地研究土壤呼吸速率变化的特征。
对滩地人工杨树林土壤呼吸速率的研究发现,一次指数模型可以较好地表示土壤呼吸对温度变化的响应,相比较而言,土壤呼吸速率与地表温度的相关性比其与土壤5cm处温度的相关性要高。同时,植被结构不同,土壤呼吸日变化动态亦不同,说明林地土壤呼吸存在空间异质性。研究者们用Q10值来说明土壤呼吸随温度的变化情况。目前所报道的众多Q10值存在着一定的差异,Raich和Schlesinger[34]经过综合研究发现其中值为2.4。全球尺度上(不包括湿地)基于气温产生的Q10值为1.5[35]。本研究中滩地人工杨树林生态系统的土壤呼吸Q10值介于1.11~1.46之间,低于不包括湿地在内的全球尺度水平。说明在长江中下游滩地森林生态系统土壤呼吸Q10值会因植被覆盖的不同而变化。
对土壤呼吸来说,最优的土壤水分状况通常接近最大田间持水量,因为此时土壤中的大孔隙多数被空气所填充,有利于O2的扩散,而且小孔隙的空间大多被水分所占据,便于可溶性基质进行扩散。当土壤处于过干或过湿状态时,土壤呼吸会受到抑制。回归分析表明,本试验地土壤5cm处水分含量与林地土壤CO2释放量的相关性不显著。原因是本研究区属于典型的亚热带湿润气候区,年降水量为1 200~1 500 mm,土壤含水量高,对土壤呼吸起制约作用。因此可初步确定该滩地人工杨树林地土壤水分不会成为林地土壤CO2释放量的限制因子,其土壤CO2释放量的变化主要受到林内气温的影响,但因水分含量高,土壤呼吸总量将减少。
另外,在进行研究期间当地气候温暖,在分析土壤呼吸速率与温度关系的散点图中,在低温区没有曲线分布,即缺少低温时土壤呼吸的特征研究。同时,在土壤呼吸的研究中,除温度和水分外,土壤肥力、大气CO2浓度以及人类活动等因子对土壤呼吸强度的影响都是不容忽视的。总之,在进行某一生态类型区土壤呼吸研究时,必须进行系统的、整体的规划和长期的定位监测工作。
[1]向元彬,胡庭兴,张健,等.华西雨屏区不同密度巨桉人工林土壤呼吸特征[J].自然资源学报,2011,26(1):79-88.
[2]Hanson P J,Edwards N T,Garten C T,et al.Separating root and soil microbial contributions to soil respiration:A review of methods and observations[J].Biogeochemistry,2000,48:115-146.
[3]张宪权,王文杰,祖元刚,等.东北地区几种不同林分土壤呼吸组分的差异性[J].东北林业大学学报,2005,33(2):46-47,73.
[4]易志刚,蚁伟民.森林生态系统中土壤呼吸研究进展[J].生态环境,2003,12(3):361-365.
[5]周存宇,周国逸,张德强,等.鼎湖山森林地表CO2通量及其影响因子的研究[J].中国科学:地球科学,2004,34(增刊Ⅱ):175-182.
[6]易志刚,蚁伟民,周国逸,等.鼎湖山三种主要植被类型土壤碳释放研究[J].生态学报,2003,23(8):1673-1678.
[7]杨玉盛,陈光水,王小国,等.中国亚热带森林转换对土壤呼吸动态及通量的影响[J].生态学报,2005,25(7):1684-1690.
[8]刘绍辉,方精云,清田信.北京山地温带森林的土壤呼吸[J].植物生态学报,1998,22(2):119-126.
[9]蒋高明,黄银晓.北京山区辽东栎林土壤释放CO2的模拟实验研究[J].生态学报,1997,17(5):477-482.
[10]杜睿,黄建辉,万小伟,等.北京地区暖温带森林土壤温室气体排放规律[J].环境科学,2004,25(2):12-16.
[11]林丽莎,韩士杰,王跃思,等.长白山四种林分土壤CO2释放通量的研究[J].生态学杂志,2004,23(5):42-45.
[12]林丽莎,韩士杰,王跃思.长白山阔叶红松林土壤CO2释放通量[J].东北林业大学学报,2005,33(1):11-13.
[13]蒋延玲,周广胜,赵敏,等.长白山阔叶红松林生态系统土壤呼吸作用研究[J].植物生态学报,2005,29(3):411-414.
[14]周存宇,周国逸,王迎红,等.鼎湖山针阔叶混交林土壤呼吸的研究[J].北京林业大学学报,2005,27(4):23-27.
[15]周存宇,张德强,王跃思,等.鼎湖山针阔叶混交林地表温室气体排放的日变化[J].生态学报,2004,24(8):1741-1745.
[16]吴仲民,曾庆波,李意德,等.尖峰岭热带森林土壤C储量和CO2排放量的初步研究[J].植物生态学报,1997,21(5):416-423.
[17]骆土寿,陈步峰,李意德,等.海南岛尖峰岭热带山地雨林土壤和凋落物呼吸研究[J].生态学报,2001,21(12):2013-2017.
[18]沙丽清,郑征,唐建维,等.西双版纳热带季节雨林的土壤呼吸研究[J].中国科学:地球科学,2004,34(增刊Ⅱ):167-174.
[19]吴建国,张小全,徐德应.六盘山林区几种土地利用方式土壤呼吸时间格局[J].环境科学,2003,24(6):23-32.
[20]冉景丞,何师意,曹建华,等.亚热带喀斯特森林土壤CO2排放量动态研究[J].贵州科学,2002,20(2):42-47.
[21]杨青,吕宪国.三江平原湿地生态系统土壤呼吸动态变化的初探[J].土壤通报,1999,30(6):254-256.
[22]郝庆菊,王跃思,宋长春,等.三江平原湿地土壤CO2和CH4排放的初步研究[J].农业环境科学学报,2004,23(5):846-851.
[23]宋长春,阎百兴,王跃思,等.三江平原沼泽湿地CO2和CH4通量及影响因子[J].科学通报,2003,48(23):2473-2477.
[24]项艳,孙启祥,程从新.长江滩地杨树无性系苗期选择研究[J].安徽农业大学学报,2002,29(3):289-292.
[25]赵广东,王兵,杨晶,等.LI-8100开路式土壤碳通量测量系统及其应用[J].气象科技,2005,33(4):363-366.
[26]Jared L D,Aslko M,Steve G M,et al.Phenophases alter the soil respiration-temperature relationship in an oak-dominated forest[J].Int.J.Biometeorol,2006,51:135-144.
[27]Ma S,Chen J,Butnor J R,et al.Biophysical controls on soil respiration in dominant patch types of an oldgrowth mixed conifer forests[J].For.Sci.,2005,51:221-232.
[28]Curiel-Yuste J C,Janssens A,Carrara A,et al.AnnualQ10of soil respiration reflects plant phenological patterns as well as temperature sensitivity[J].Global Change Biol.,2004,10:161-169.
[29]张东秋,石培礼,张宪洲.土壤呼吸主要影响因素的研究进展[J].地球科学进展,2005,20(7):778-785.
[30]李明峰,董云社,齐玉春,等.锡林河流域羊草群落春季CO2排放日变化特征分析[J].中国草地,2003,25(3):9-14.
[31]陈全胜,李凌浩,韩兴国,等.水热条件对锡林河流域典型草原退化群落土壤呼吸的影响[J].植物生态学报,2003,27(2):202-209.
[32]Bridge B J.The formation of degraded areas in the dry savanna woodlands of northern Australia[J].Aust.J.Soil Res.,1983,21:91-104.
[33]Mathes K.The soil respiration during secondary succession:influence of temperature and moisture[J].Soil Biol.Biochem,1985,17(2):205-211.
[34]Raich J W,Schlesinger W H.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J].Tellus,1992,44B:81-89.
[35]Raich J W,Potter C S.Global patterns of carbon dioxide emissions from soils[J].Global Biogeochemical Cycles,1995,9:23-36.
Studies on Soil Respiration Change and Its Relation with Environmental Factors in Poplar Plantation on Beach Land
MA Tao1,ZHOU Jin-xing2
(1.Gansu Institute of Soil and Water Conservation,Lanzhou730020,China;2.Institute of Desertification Studies,Chinese Academy of Forestry,Beijing100091,China)
In order to reveal the soil respiration change and its relation with environmental factors,soil CO2emission rates in artificial poplar plantation on Beach Land were investigated by using Infra Red Gas Analysis(IRGA)technique,and the soil respiration changes in relation to environmental factors such as temperature,soil water content and vegetation structure underforest were analyzed.The results showed that the diurnal dynamics of soil respiration presented a single peak curve with the maximum occurring around 11:00-13:00.The order of soil CO2emission rates is Jun-2006>Oct-2005>Mar-2006>Oct-2006.The mean value of efflux is 2.29±0.75μmol/(m2·s)(P<0.05,n=13)and a coefficient of variation is 54.40%.The sensitivity of the soil respiration to the temperature above ground is superior to the soil temperature at 5cm.The Temperature susceptibility indexesQ10value is 1.28±0.79(P<0.05,n=13),which is less than the global level except for beach land.The relationship between soil moisture and soil respiration is not good,so the soil moisture is not the limitational factor to the efflux of CO2.The vegetation structure is also the one influence factor controlling soil respiration.
poplar plantation;soil respiration;environmental factors;Q10value;beach land
S715.71;S154.4
A
1005-3409(2011)06-0031-06
2011-04-10
2011-06-17
“十一五”国家林业科技支撑项目(2006BAD03A15)
马涛(1983-),男,甘肃天水人,硕士,工程师,主要从事流域治理与技术服务研究。E-mail:mataosbsgsu@126.com
猜你喜欢
热带作物学报(2022年8期)2022-09-16
鸭绿江(2020年15期)2020-11-12
黑龙江粮食(2019年4期)2019-05-10
自然保护地(2019年1期)2019-03-18
中国农村水利水电(2018年12期)2018-12-28
农民致富之友(2018年2期)2018-03-25
农民致富之友(2018年1期)2018-02-26
前卫文学(2017年5期)2017-11-09
中国水土保持(2017年6期)2017-01-21
中国水土保持科学(2016年4期)2016-09-21
