
不同饵料搭配对凡纳滨对虾幼体与仔虾的影响
2011-06-08 13:05:40王家伟戴习林臧维玲丁福江
湖南农业科学 2011年15期
王家伟,戴习林,谢 剑,臧维玲,丁福江
(1.上海海洋大学水产与生命学院,上海 201306;2.上海申漕特种水产开发公司,上海 201376)
1 材料与方法
1.1 试验材料
试验于2008年3~5月在上海申漕特种水产开发公司进行。试验用幼体为购自厦门的凡纳滨对虾无节幼体(子一代,F1),在长方形 51×61×38 cm3塑料箱(118L)内培养。试验用水采用浓缩海水与天然河口水调配成盐度为28.6的育苗海水,过滤、消毒后使用。
1.2 试验饵料
试验用饵料为牟氏角毛藻(Chaetoceros mulleri,CM)、绿色巴夫藻(Pavlova vilids,PV)、湛江叉鞭金藻(Dircaraleria galbana,DG),云微藻(Chlorophyta sp,Cs)、褶皱臂尾轮虫(Brachionus plicatilis,BP)、蒙古裸腹溞(MM),以及购自市场的卤虫(Aitemia salina,AS)卵、双色虾片、螺旋藻粉、食母生、B.P粉(一种微囊饲料)和对虾人工配合饲料(FD)。
单胞藻用f/2配方配制的营养液培养;采用酵母(YE)为主、单胞藻为辅,交替饲养BP和MM,分别在投喂幼体前8 h和12 h用单胞藻营养强化。
1.3 试验设计
试验设计6种饵料搭配,每种2个重复,每组对虾无节幼体(N)密度为4~5万尾/m3。以全FD为对照组,试验用饵料种类和搭配列于表1。
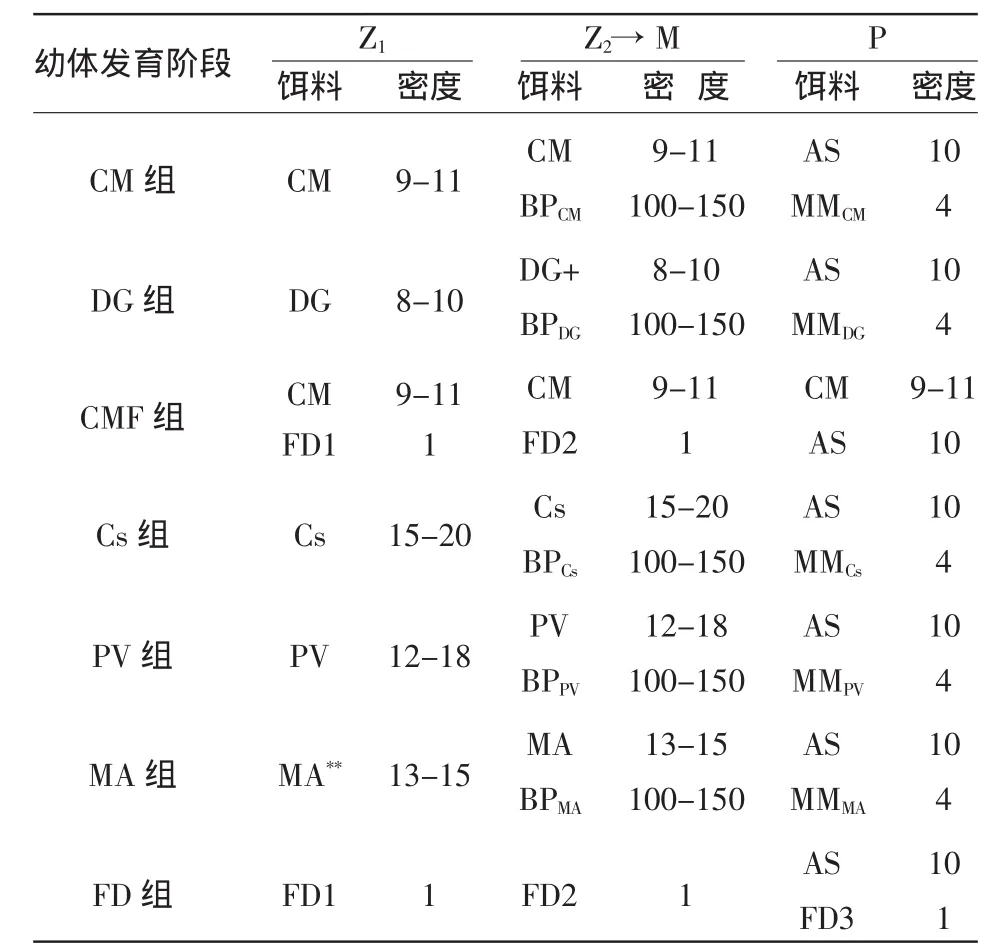
表1 各试验组饵料投喂种类和密度
1.4 试验管理
试验期间,连续充气,水温30±0.2℃,光照强度3 000~10 000 Lx。每日测定水化学指标和饵料密度,自M1开始每日换水20%,上午每个水槽随机采样5点估算幼体数,记录幼体发育状况。
1.5 脂肪酸和水质指标的测定
总脂抽提参照Folch法[7],采用14%的三氟化硼-甲醇(v/v)进行甲脂化,仪器为Agilent-6890气相色谱。水质测定方法:CODMn采用碱性高锰酸钾法,NO2--N采用重氮偶氮比色法,NO3--N采用锌镉还原-重氮偶氮比色法[8],pHB-4型 pH计测定pH值。
2 结果与分析
2.1 饵料脂肪酸含量与组成
2.1.1 单胞藻脂肪酸含量 单胞藻脂肪酸相对含量测定结果列于表2。由表2知,PV的n-3HUFA和∑EFA相对含量最高分别为51.7%和31.2%,CM、DG次之,Cs最低,分别仅为PV的1/6和1/2。C18∶3n-3仅在PV和CM中检出。EPA在PV和CM中相对含量高,分别为27.9%和23.8%,Cs中未检出。DG、PV的DHA相对含量都很高,分别为19.8%和12.6%,Cs较低为8.8%。
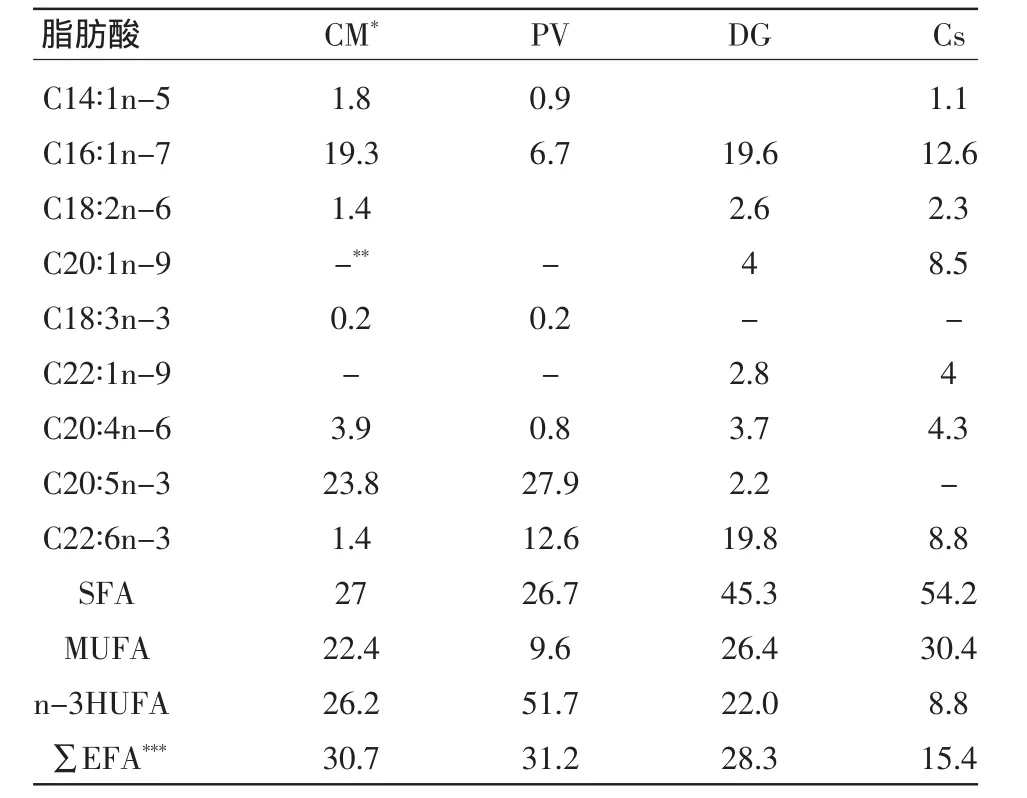
表2 单胞藻脂肪酸相对含量
2.1.2 营养强化轮虫的脂肪酸含量与组成 营养强化后的BP体内脂肪酸相对含量列于表3。由表3可知,BPCs体内∑EFA、n-3HUFA的相对含量最高,分别为17.4%与26%,次之为BPCM,分别是23.3%与17%。C18∶3n-3在5种藻液强化BP体内均未检出,DHA仅在BPDG体内未检出,其余3种必需脂肪酸均被检出。5种藻液强化BP体内EPA和DHA相对含量都存在较大差异,BPCs体内DHA和EPA含量最大分别为17.4%和8.6%,分别是BPDG的3倍和17倍。
供给情况:氮肥方面,尿素企业复产停产交替,总体以复产增产居多,企业整体开工率小幅上升至54%。磷肥方面,个别前期短期检修的企业陆续复产,二铵企业整体开工率升至66%。钾肥方面,盐湖基准产品60%粉晶到站价2200-2250元/吨,每日发运量200-300车;港口钾供应持续紧俏;边贸钾供应仍偏紧。复合肥方面,经销商拿货缓慢,部分企业因库存较多降低产量,各地环保压力不减,企业整体开工率下降至40%。
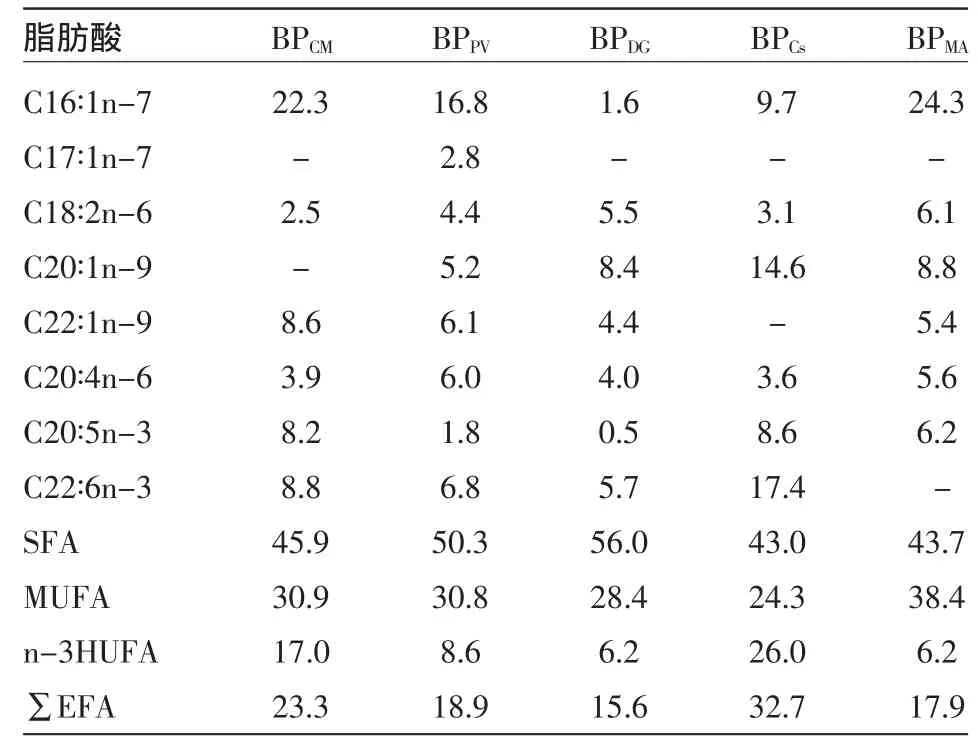
表3 单胞藻营养强化轮虫的肪酸相对含量
2.1.3 营养强化后蒙古裸腹溞的脂肪酸含量 单胞藻营养强化的MM的脂肪酸相对含量测定结果列于表4。由表4可见,MMDG和MMPV体内∑EFA的相对含量最高,分别为44.3%和42.4%,约为MMYE的2倍,而MMCs和MMMA体内∑EFA相对含量低于MMYE。MMPV体内EPA的相对含量最高为13.5%,是MMMA的4倍,MMDG虽未检到EPA,但DHA含量最高,为31.6%,分别是MMPV、MMCM、MMYE的 1.5、2、4 倍,而 MMCs和 MMMA未检出。所有组MM体内未检出C18∶3n-3,∑n-3HUFA含量呈现与∑EFA一样的规律。
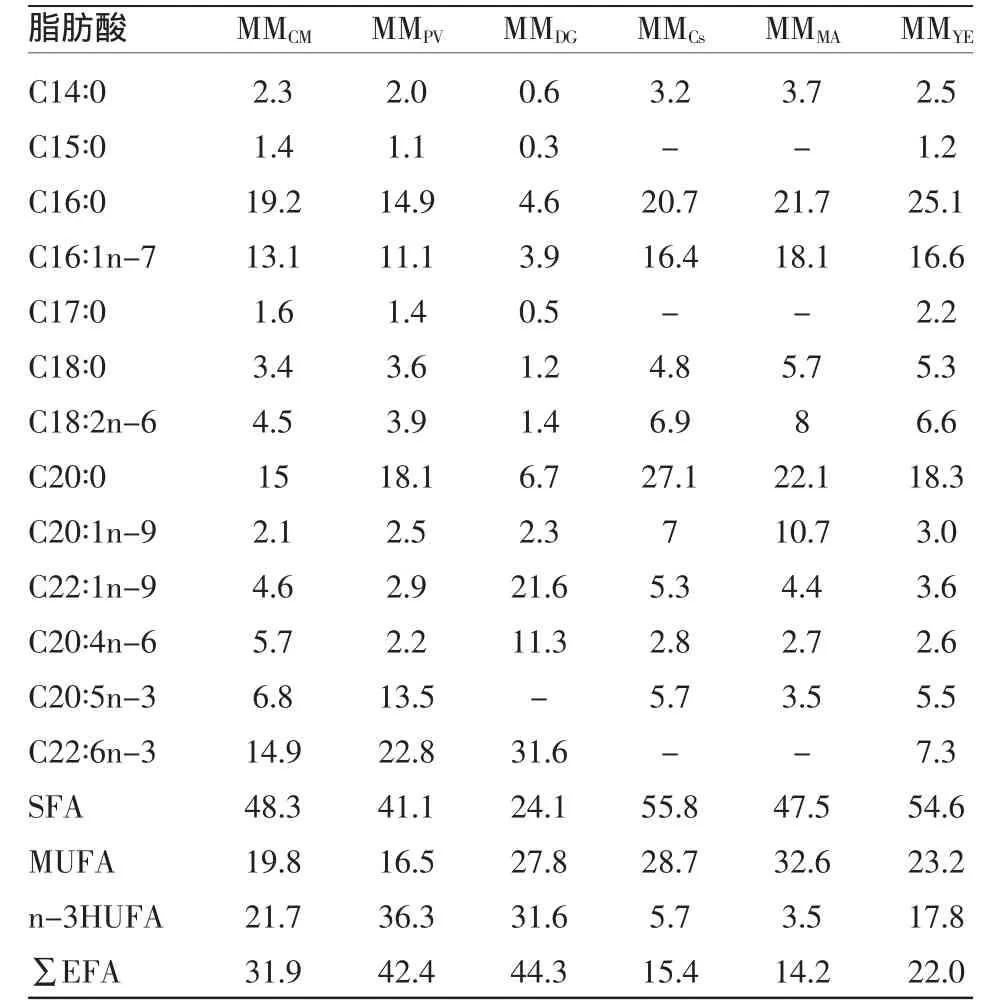
表4 单胞藻营养强化蒙古裸腹溞脂肪酸相对含量
2.2 凡纳滨对虾幼体培育效果
凡纳滨对虾仔虾和幼体各阶段存活率及发育情况分别列于表5和图1、图2。表5表明,N变态为Z1阶段(N→Z1),投喂Cs和混合藻的幼体存活率r最低,分别只有78.4%和65.8%,其它组r均在80%以上。Z1→Z2,CM、DG、CMF、PV 组是高 r组,Cs组最低为45.6%。
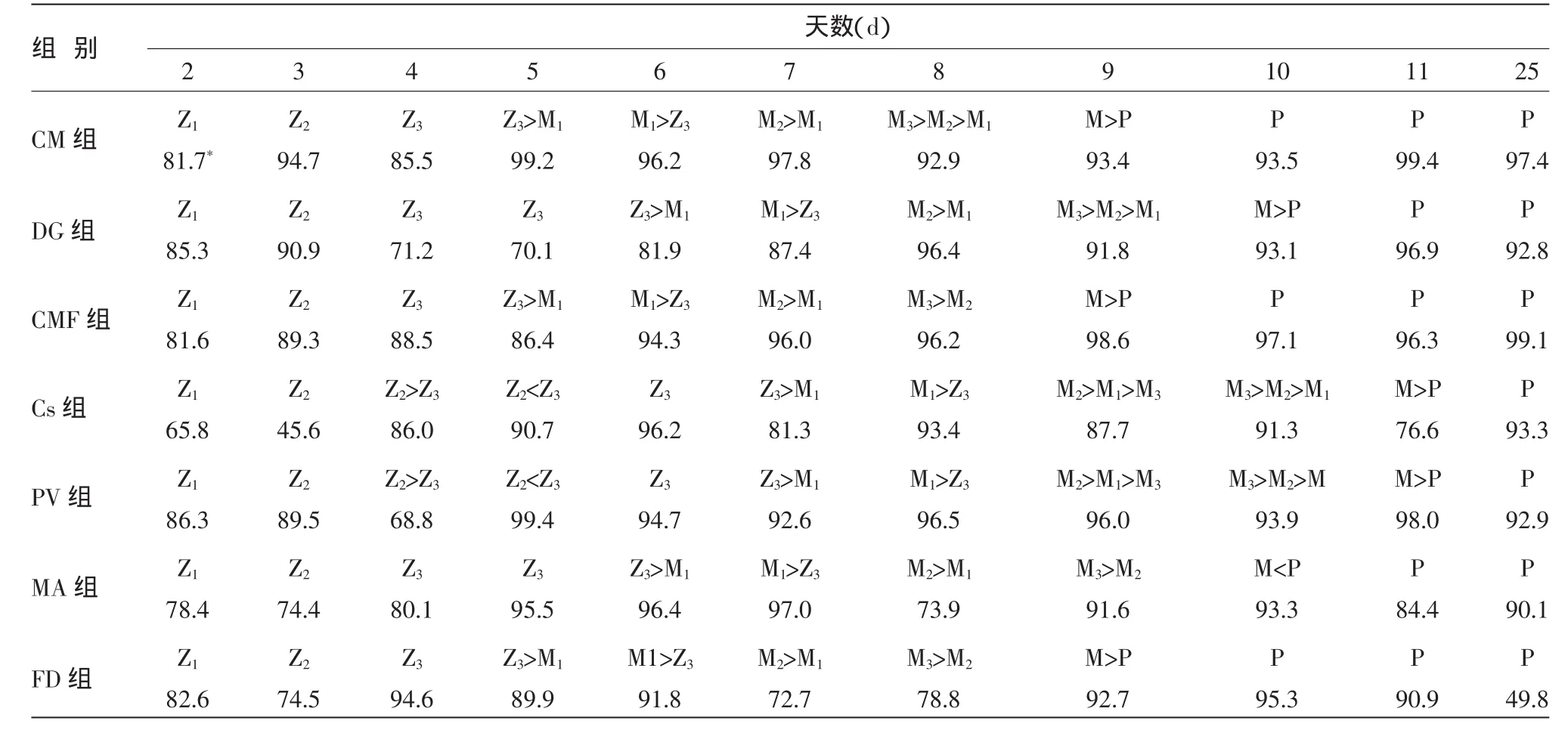
表5 凡纳滨对虾幼体、仔虾发育状况与存活率
由图 1可知,Z2→Z3,FD 组 r最高,达到94.6%,CM与CMF组次之,且三者间无明显差异,Cs、DG和PV组r最低,分别只有75.0%、71.2%和64.8%,且相互间无差异。Z3→M1,CM组r最高为93.4%,显著高于PV、CMF组的80.8%与78.1%,DG组最低为48.3%。
M1→P,幼体变态情况分别是r为90%组(CMF、PV 组)、80%组 (DG、CM 组)、70%组(MA、Cs、FD组)。P阶段,FD组r最低为49.8%,其余组均在90%以上。
由表5可发现,各实验组N变态为Z2与M变态为P速度一致,PV和Cs组Z2,2 d才发育至Z3,其余组仅1 d;CM组、CMF组与FD组Z3,3 d内全部变态为M1,而PV藻组和Cs组需5 d。
由图2可知,出苗率最高发生在CM藻与CMF组,PV组次之为26.2%,Cs与FD组最低,低于10%。
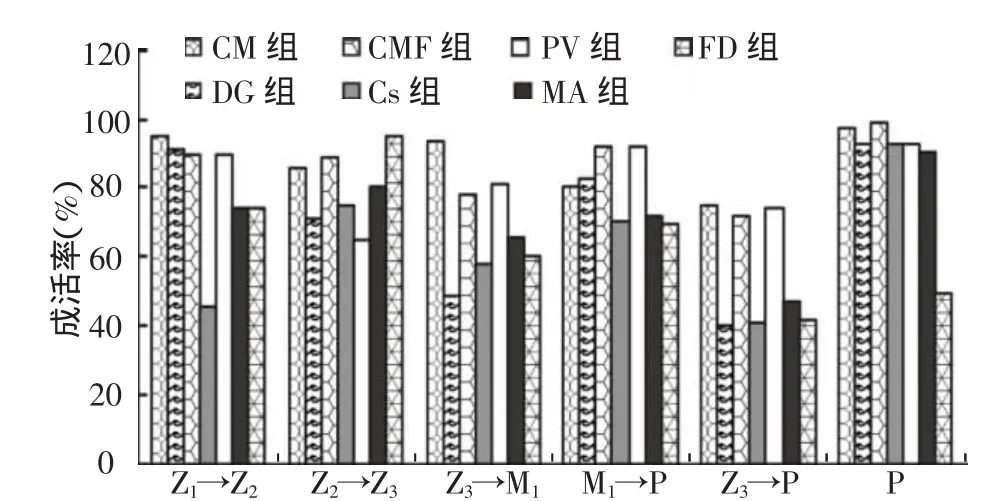
图1 凡纳滨对虾幼体各阶段的存活率
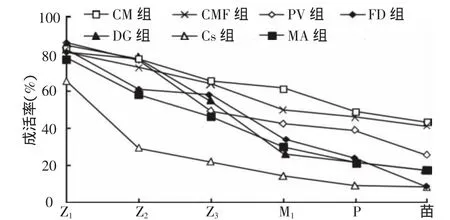
图2 凡纳滨对虾幼体各阶段的存活率
3 讨 论
3.1 轮虫和蒙古裸腹溞的营养强化效果
BP和MM是人工育苗中的重要生物饵料,但是酵母培养的BP和MM体内EPA和DHA含量很低,有资料报道酵母轮虫n-3HUFA、EPA和DHA的相对含量分别只有0.9%、0.7%和0.2%[4]。除MMDG 的 EPA 未检出外,MMCM、MMPV、MMCs体内∑EFA、n-3HUFA、EPA和 DHA含量都高于MMYE,营养强化的MM和相对应的藻类体内脂肪酸的组成和相对含量趋于一致,此与何志辉等[11]的研究结果一致,表明蒙古裸腹溞的生化组成明显受食物条件的控制。
同时试验所用4种藻类除BPDG的EPA含量没有提高外,BPCM、BPPV、BPCs体内 n-3HUFA、EPA 和DHA含量都明显高于BPYE;Cs中n-3HUFA、EPA含量最低,但BPCs中n-3HUFA、EPA含量最高,PV体内n-3HUFA、EPA含量最高,但其BPPV体内n-3HUFA、EPA含量却较低,营养强化的BP和相对应的藻类体内脂肪酸的组成和相对含量并不趋于一致,与陈炜[12]的研究结果相似,表明轮虫的脂肪酸组成明显受饵料影响,但又不完全取决于饵料。因此,采用合适的单胞藻强化BP和MM,可显著提高它们体内的HUFA,而且MM受食物的影响要大于褶皱臂尾轮虫,生产过程中,可选择HUFA含量高的单胞藻强化MM,而对于BP并非如此。
3.2 HUFA对凡纳滨对虾幼体存活率的影响
虾蟹类中,机体吸收的部分n-3HUFA尤其是EPA和DHA能用于合成磷脂,构成磷脂双层膜,磷脂能促进胆固醇 (合成蜕皮激素的前体物质)的吸收和利用,n-3HUFA间接影响了虾蟹类幼体的蜕皮与提高幼体变态和存活率[13]。但大部分海水虾蟹类却不能合成EPA和DHA,只能从食物中获得[14]。Z1→Z2,r最低组是n-3HUFA含量最低的Cs组,甚至低于FD组;BPDG体内n-3HUFA含量最低,尤其是EPA含量仅占总脂含量的0.5%,对应的Z2→M阶段,投喂BPDG组r最低,幼体发育速度较慢。可见饵料中n-3HUFA的含量显著影响幼体发育,BPCs、BPDG不能完全满足幼体发育的需要。
比较藻类脂肪酸含量发现,CM含有全部的对虾5种必需不饱和脂肪酸,而其它藻类体内最多检到4种,这可能是CM组和CMF组的各阶段幼体与仔虾r高,变态速率快的原因之一。梁华芳等[15]也发现CM投喂凡纳滨对虾效果要好于PV、球等鞭金藻、中肋骨条藻。
对于Cs组,又发现即使在Z2→M阶段投喂HUFA最高的BPCs,幼体的存活率仍较低,且Z2、Z3生长缓慢,2 d才全部变态为Z3,5 d全部变态为M1。此可能是Z1阶段幼体饵料HUFA含量低,进而影响Z2→M阶段幼体发育,这与Legger等[16]发现蓝对虾早期幼体摄取的HUFA对后期幼体的营养状况有明显影响的结论一致。
3.3 饵料搭配对凡纳滨对虾幼体发育的影响
BPMA与MMMA体内HUFA均最低,且均不含DHA,MA组的Z1→M的幼体变态率均较低,Z3发育缓慢,并没有体现出MA营养互补,增加强化效果,促进幼体发育的优势。这可能由于几种藻混合使用产生“毒性”作用,曹淑莉等[17]也作了类似的报道。
有资料表明卤虫HUFA含量低,本次试验的仔虾培育阶段,AS与CM或强化MM搭配饲养的仔虾r是AS搭配FD的2倍,这表明AS与CM或强化MM都能形成营养互补,提高仔虾存活率。
投喂合适的单胞藻、BP、MM等活饵料的虾苗出苗率优于FD组,分析两者育苗效果不同的原因,首先是活饵料均匀散布于水体中,对虾幼体可一直摄取,而配合饲料易沉底,过量投喂也难以提高摄食率;其次是配合饲料易败坏水质,试验中投喂过FD的CMF组与FD组水体中弧菌含量和CODMn 分别为 440、400、5.54、5.90 mg/L,明显高于未投FD组,而活饵料不仅对水质污染小,单胞藻还可净化水体,做动物性活饵料的营养强化饲料。
综上所述,CM或Cs可用来强化BP,CM或PV可用来强化MM,CM、PV和DG可作为凡纳滨对虾Z1的开口饵料,后阶段幼体培育阶段可用来调节水质,并作辅助饵料,但不宜采用两种或两种以上的MA强化或投喂。可用强化BP培育凡纳滨对虾Z2→M,可交替使用强化蒙古裸腹溞与卤虫无节幼体培育仔虾。在生产中可采用BPCM或BPCs、AS、MMPV的饵料搭配培育凡纳滨对虾虾苗。
[1] 朱丽岩,郑家声,王梅林.不同饵料及添加剂对中国对虾幼体的影响[J].海洋科学,2000,24(11):41-44.
[2] 张 煜,臧维玲,刘永士,等.淡水养殖凡纳滨对虾的瞬时耗氧速率与体长、溶氧水平关系研究[J].湖南农业科学,2010,(5):136-139.
[3] 蓝汉冰,曹俊明,许丹丹,等.饲料中添加核苷酸粗提物对凡纳滨对虾生长性能的影响[J].广东农业科学,2009,(10):143-145.
[4] 陈 炜,王 鑫,童圣英.海洋酵母培养褶皱臂尾轮虫的脂肪酸组成研究[J].水产科学,2001,20(3):3-6.
[5] 童圣英,林成辉,王雪涛.蒙古裸腹溞营养成分分析与评价[J].大连水产学院学报,1988,(Z1):29-33.
[6] 曾庆华,周洪琪,黄旭雄,等.我国六个产地卤虫初孵无节幼体的营养价值[J].上海水产大学学报,2001,10(3):213-217.
[7] Folch J M L,Sloane G H S.A simple method for the isolation and purification of total lipids from animal tissues[J].J Bio Chem,1957,226:497-509.
[8] 国家质量监督局.海水分析[S]//海洋监测规范.北京:中国标准出版社,1998.142-142,150-162.
[9] 李荷芳,周汉秋.海洋微藻脂肪酸组成的比较研究[J].海洋与湖沼,1999,34(1):31-40.
[10] 季文娟,徐学良.中国对虾(Penaeus orientalis Kishinouye)的几种必需脂肪酸营养价值的比较研究 [J].中国水产科学,1994,(1):1-9.
[11] 何志辉,秦建光,阎立光,等.一种盐水裸腹溞的生物学和培养的研究[C].全国鱼虾饲料学术讨论会文集,1988.188-192.
[12] 陈 炜,李晓东,王增立,等.鸡粪浆及5种微藻对轮虫脂肪酸组成的影响[J].水产科学,2006,25(3):113-116.
[13] 陈立侨,江洪波,周忠良,等.ψ-3HUFA对中华绒螯蟹幼体存活率及体脂肪酸组成的影响[J].水产学报,2000,24(5):448-452.
[14] 成永旭,严生良,王 武,等.饲料中磷脂和多不饱和脂肪酸对中华绒螯蟹大眼幼体育成仔蟹的存活率和生长的影响 [J].水产学报,1998,22(1):9-15.
[15] 梁华芳,龙建艺,林小彬,等.5种单胞藻培育南美白对虾溞状幼体的饵料效果[J].海洋科学,2001,25(7):9-10.
[16] Leger P,Beiber G F,Soregloos P.International study on ArtemiaⅩⅩⅩⅢ.Promising results in larval rearing of Penaeus stylirostris using a prepared diet as algal substitute and for Artemia enrichment[J].J World Aquac Soc,1985,16:354-367.
[17] 曹淑莉,向葆卿,房 慧,等.几种海洋微藻喂养中国对虾溞状幼体饵料效果的试验研究[J].海洋通报,1990,9(3):56-62.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05 04:33:14
生物学通报(2021年5期)2021-03-16 06:42:26
科学养鱼(2020年10期)2020-11-23 09:04:10
海洋通报(2020年3期)2020-10-20 09:26:40
课外语文·中(2020年2期)2020-03-24 22:18:09
课外语文(2020年5期)2020-03-14 08:16:40
当代水产(2019年1期)2019-05-16 02:42:00
知识窗(2019年4期)2019-04-26 03:16:02
广东海洋大学学报(2015年4期)2016-01-13 08:39:32
应用海洋学学报(2014年1期)2014-11-22 07:17:44
