
乌冈栎地理分布与水热环境因子的关系
2011-05-07 10:59:20谢春平方炎明
水土保持研究 2011年1期
谢春平,方 彦,方炎明
(1.南京森林警察学院 侦查系,南京210046;2.南京林业大学 森林资源与环境学院,南京210037)
植物在地球上已有上亿年的时间,从结构最简单的单细胞类群到今天纷繁复杂的被子植物,每一次植物类群的大更迭都与气候环境有着不可分割的密切关系。在长期的进化过程中,每一种植物存在于一个适于生长和发育的地理区域内,形成了与气候因素(例如温度、降水、光照等)相适应的格局[1],因此气候是在大尺度上决定物种分布的主要因素[2-3]。植物的分布与气候因子尤其是水热因子的关系已从定性描述发展到定量研究[4],目前常用的指标有Kira的温暖指数(WI)、寒冷指数(CI),徐文铎的湿润指数,Penman的可能蒸散和干燥度,Thomthwaite的可能蒸散和水分指数,Holdridge的生命地带分类系统指标生物温度和可能蒸散率等[5],这些指标均在植物分布区生态适应性方面的研究得到了广泛的应用[6-10]。近年来,随着新的统计技术和地理信息系统的发展,用于预测物种分布的模型技术也得到了迅速发展,并作为一种重要的工具被广泛地应用于生物地理分布、生物多样性和气候影响评估等方面[11]。
乌冈栎林(Quercus phillyraeoides forest)主要分布于陡坡峭壁或山脊部位,生境多表现为基岩裸露、土层浅薄。一般说来,乌冈栎林始终是一个较稳定的森林群落;但人为的干扰破坏若接踵而至,则森林有向逆向演替的可能,到那时情况就十分严重,先是林地沦为荒草、灌丛,继而出现光山秃岭,水土流失,所造成的恶果将不堪设想。因此,乌冈栎是山顶、山崖等贫瘠地段的重要水土保持树种,其森林的覆盖对当地起到十分重要的水土保持作用。由于它的分布区范围较广,气候、地形地貌、土壤环境等复杂多变,由此导致环境因子发生剧烈变化,这些变化必然会给乌冈栎的生长和发育带来各种复杂的影响,形成许多具有适合当地气候的地理生态型。通过对乌冈栎地理分布气候特点的研究,本文拟解决以下几个问题:(1)乌冈栎适生区有哪些地方,其分布的南北界大致在什么范围;(2)限制乌冈栎分布的主要气候因子有哪些;(3)建立分布区气候数学模型;(4)大致对乌冈栎的分布区进行气候区划。因此,本文旨在通过对乌冈栎不同分布区与气候因子变化的相关研究,为该物种的开发利用提供一定的科学参考依据。
1 研究方法
1.1 数据收集
1.2 生态适应性指标
温暖指数(Warmth index):
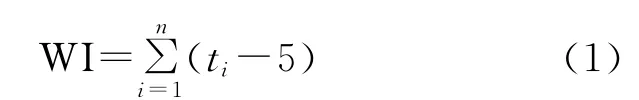
寒冷指数(Coldness index):
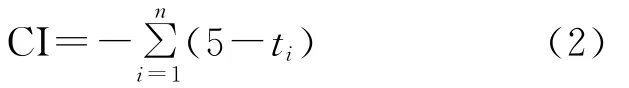
干燥度指数(Aridity index):
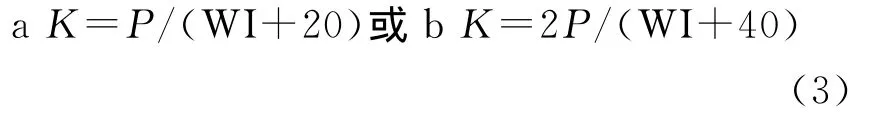
水热综合指数(Integrated index of precipitation and temperature):
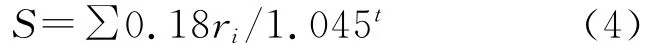
最适热量分布范围(Peak width at half height):
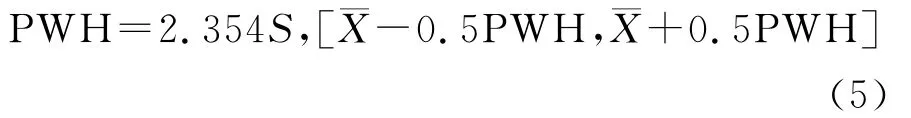
生物温度((Biotemperature):

可蒸散量(Potential evapotranspiration):
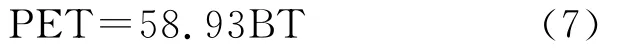
年均可能蒸发量率(Potential evapotranspiration ratio):
1)土地利用程度综合指数模型。土地利用程度综合指数是计算区域内的土地利用分级指数加权平均数,可用于综合评价该区域土地利用程度。土地利用程度综合指数值越大,人类活动对土地利用的程度就越大,反之,则说明该地区的土地利用有较大的发展潜力。
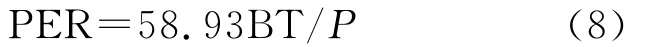
其中,ti在(1)式为大于5℃的月均温,在(2)式中位小于5℃的月均温;(3)式中,当WI≤100时采用a式,WI>100时采用b式,P为年平均降水量(mm);(4)式中ri为月降水量(mm),t为月平均温度;(5)式中¯X为温暖指数的平均数,S为标准差;(6)(8)式中t、P 同式(3)(4)。
1.3 统计与分析
文中所有绘图与数据统计分析,均由DIVAGIS 5.2、Excel 2003以及MVSP 3.1完成。
2 结果与分析
2.1 乌冈栎的地理分布
根据相应的文献,利用DIVA-GIS 5.2绘制出乌冈栎在东亚地区的水平分布格局(图1)。中国乌冈栎的分布范围大致在23°-34°N,100°-120°E,其分布范围涵盖了中国亚热带的大部分地区;分布多度格局表明,乌冈栎分布大致有:华东地区以武夷山脉、怀玉山脉和黄山为主要分布中心;华中地区的乌冈栎分布以秦岭南坡-伏牛山为界,主要分布在大巴山脉、武陵山脉、雪峰山等一带,并延伸至云贵高原的滇东南、黔东南、四川盆地周边等小部分地区,并且呈现出零星分布状,大多未形成典型的乌冈栎群落;华南地区分布主要集中在南岭山脉,并延伸至桂北与湘西南接壤的地方。由此从分布图的整体可以看出,乌冈栎在中国的分布大致以长江秦岭为界,其分布中心在华东的浙江西南部、福建大部以及安徽与江西交界处;其余地区大多呈星散状。日本乌冈栎分布的纬度要略高于中国,其范围大致在26°-37°N,128°-140°E,主要分布在琉球群岛、九州岛、四国岛和本州岛,多生长于海岸沿线或内陆山地,形成低矮的纯林[13-16]。
乌冈栎多是分布于陡峭的悬崖或山脊之上,其分布地以丹霞和石灰岩地貌为主;垂直分布在300~1 300m,不同海拔之间的生长及林相有较大的区别,随着海拔的升高其森林资源量逐渐减少,并形成灌木状与其他物种混生。
2.2 乌冈栎分布区气候环境指数
由于中国内地乌冈栎分布区环境较日本诸岛屿情况复杂,因此在统计分析时,将中日两国的气候环境指数分开,具体包括:一月均温(TM-Jan)、七月均温(TM-Jul)、最 高 温 (TMax)、最 低 温 (TMin)、年 均 温(TM-A)、年均降水量(PM)、温暖指数(WI)、寒冷指数(CI)、干燥度指数(K)、水热综合指数(S)、年均生物温度(BT)、可蒸散量(PET)和年均可能蒸发量率(PER)共计13项,详见表1。
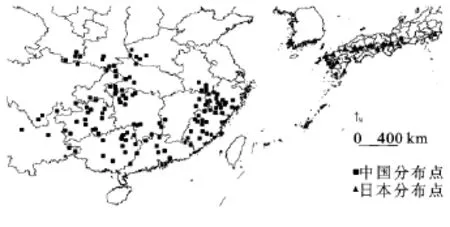
图1 乌冈栎东亚地区地理分布点
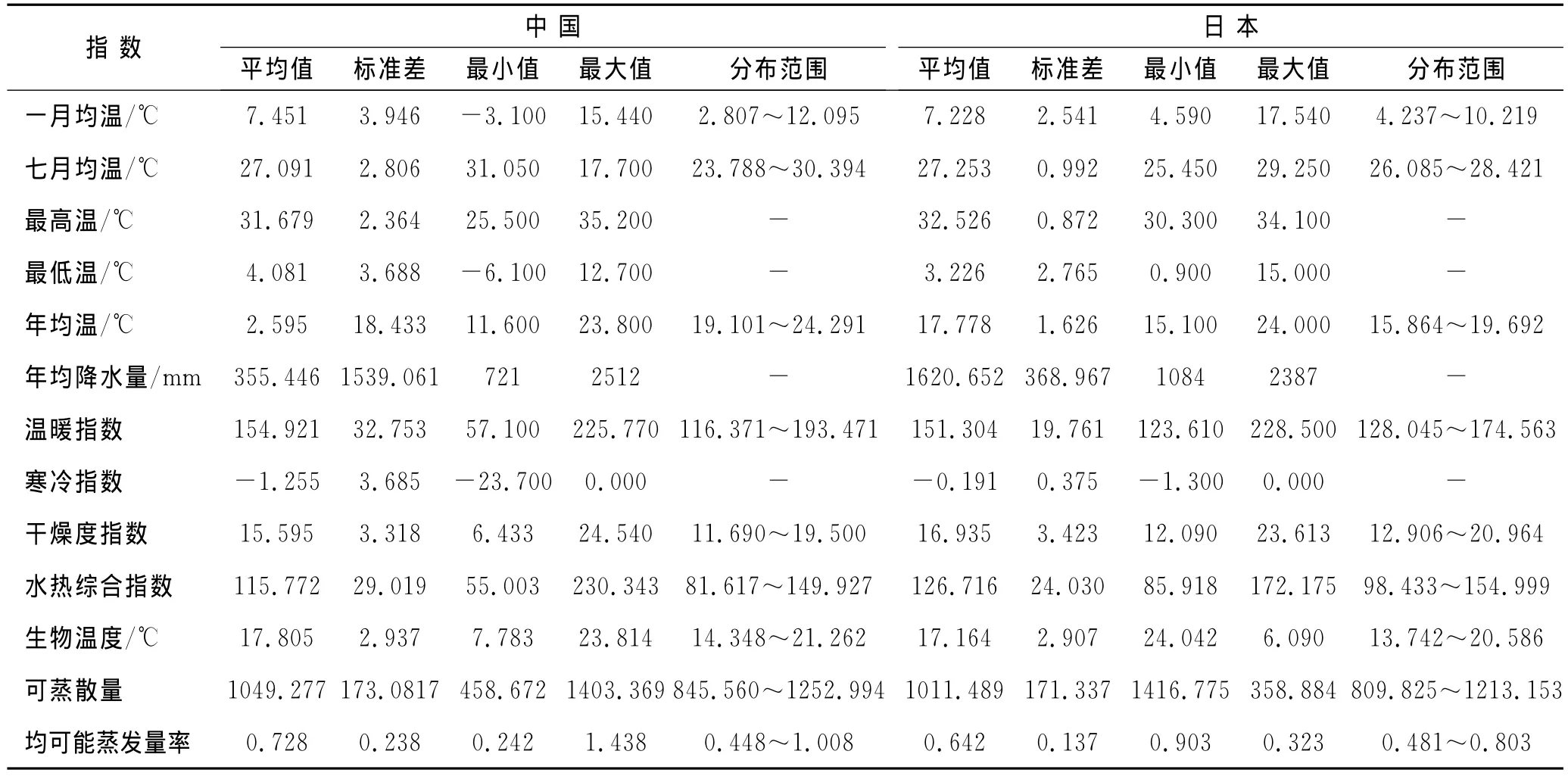
表1 乌冈栎地理分布气候指数
表1前6项指数是原始气候参数的综合反映,从这些数据可以看出,乌冈栎适应的气候范围相对是较宽泛的。如1月与7月均温的平均值分别为7.451℃和27.091℃,最适宜范围为2.807~12.095℃和23.788~30.394℃,这与我国秦岭以南大部分地区1月和7月气温相符合;日本诸岛屿的情况与我国相似,但其变化幅度要小于我国内地,1月与7月最适宜范围为4.237~10.219℃和26.085~28.421℃。从年均降水量可以看出,中国内地分布区和日本诸岛屿年均降水量均在700mm以上,其范围分别为721~2 512mm与1 084~2 387mm,两地降水量都较为充沛,并且都呈现出雨热同期的气候特点,较为集中的降水集中在5-8月。另外,从年均温看,两地均温较为接近,但最适宜分布范围,中国内地要高于日本诸岛4℃左右。另外,通过4个气候指数的累积分布图(图2)可以看出,年均温度大多集中在17℃左右,年均温超过20℃区域约为10%(A);年均降水量以1 600mm为中间值,大于2 000mm分布的区域仅在5%的范围,大部分地区降水都超过了1 000mm(B);另外,等温性累积图说明在乌冈栎分布区中,大多数地区年温度变化为25~35℃,温度变化较大,即季节性温度的更替明显(C);最后,最低温度的变化范围中,近20%区域是在0℃以下,其余大部在0℃以上,这表明乌冈栎分布区并没有真正意义上的严寒(D)。
年均生物温度、降水量和可能蒸散发量率是生命地带分类系统的反映指标[17],中国内地乌冈栎分布区内的年均生物温度为BT=17.805℃,年均可能蒸发量率为PER=0.728,日本诸岛屿则分别为17.164℃和0.642。与其它树种进行比较发现,乌冈栎的BT值和PER值与泛亚热带分布的青冈(Cyclobalanopsis glauca)的值接近(BT=16.9℃和PER=0.778)[8],而与西部白梭梭(Haloxylon persicum)、沙拐枣(Calligonum mongolicum)等10种植物的BT值和PER值相距甚远[18]。因此,两地乌冈栎分布区符合Holdridge(BT>17℃)和中国植被(BT>14℃)[19]所划分的亚热带区,归属于典型的亚热带树种。
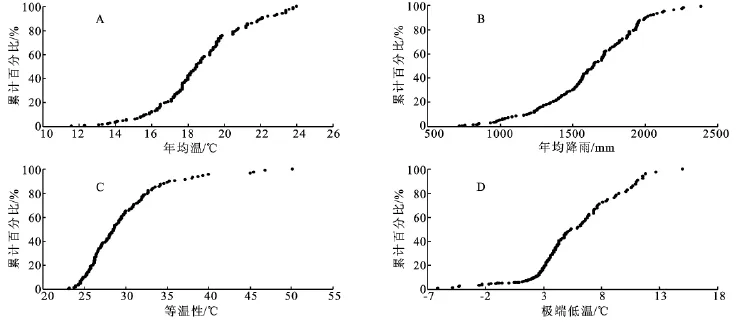
图2 四个气候指数累积分布图
2.3 热量分布曲线
以横坐标代表温度指标(WI),纵坐标代表树种出现的频率(f),将不同温度指标值上出现的树种频数连成一曲线[20],对中国内地及日本列岛的乌冈栎热量分布曲线进行讨论(图3)。对分布频数划分为若干级别:WI<60为一级,此后每隔20为一级直至220,最后WI>220为一级别,共分为10级。树种的分布与热量条件之间的关系通常是两侧对称的正态分布曲线[21],从图3可以看出,乌冈栎在中国内地的分布和在日本中部至南部诸岛屿的分布都呈正态分布。中国内地热量分布在50~230,6、7级占有较大的比例,但整体分布范围较为宽泛;日本岛屿热量分布在120~230,变化幅度较中国内地小,第5级占有的比例较为突出;而两地最适分布值也集中在5~7级,证明了该物种为偏温性树种,大约在北纬23°以南就没有乌冈栎的天然分布了。分布与热量条件之间正态曲线的形式在另一方面也反映了树种分布的广度,由此不难看出中国内地乌冈栎分布的范围要远大于日本的分布,这与图1分布点反映出来的情况相吻合。
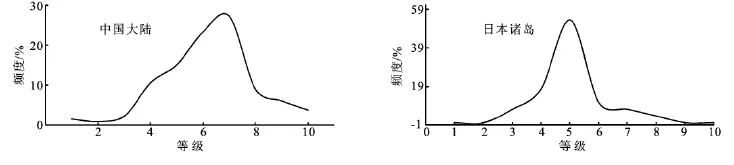
图3 乌冈栎温暖指数分布曲线图
2.4 气候因子主成分分析
主成分分析(Principal Component Analysis)是一种通过降维的方法将多个指标简化为少数几个综合指标,使这几个少数综合指标可以反映原来所有指标的信息的多元统计方法,它从本质上讲是聚类方法的一种;但除此之外,主成分分析还可以提供有关性状间相关性的信息,以及某一综合性状在分类中的意义及其对分类的影响程度等多种信息,因此说主成分分析有着它独特的优越性[22]。
利用MVSP 3.1计算选取经度、纬度、海拔、温度、水分等9个变量主成分数值,结果见表2。由于当m=3时,9个主成分累计贡献率达85.51%(>85%),它足以代表原始因子所代表的大部分信息,即主成分分析方法在对乌冈栎分布区气象指数相关性研究上是可行的。表2中第一主分量的贡献率为55.56%,与其相关的主要指数是年平均温度(0.425)、极端最低温(0.405)、1月均温(0.374)等,说明其分布受温度的影响有较大。第二主分量的贡献率为19.31%,与其相关的指数主要是海拔(0.517)、纬度(-0.462)、极端高温(-0.458)、7月均温(-0.386)等,说明在纬度梯度或海拔梯度影响下,高温对乌冈栎分布区有较大影响作用。第三主成分的贡献率为10.63%,其影响较大的是经度(-0.891)和年均降水量(-0.313),反映了年均降水量与经度的关系,这也与我国降水量由东至西逐渐减少的降水规律相吻合。其余6个主分量的贡献率都较低,仅占贡献率的14.49%,对整体结果分析影响不大。综合上述的前三个主成分分析可以看出,影响乌冈栎分布的最主要因素集中在温度方面,而降水的重要性在第三主成分中才体现出来,此时的贡献率(影响作用)已经不大,因此以乌冈栎为优势种的植被类型的性质——干旱性植被得到了进一步的验证。

表2 气象指数主分量特征根及其特征向量
根据前2个主成分做出2维主成分分析图(图4),可以将其归结为以下几个大类群,这一划分与水热条件分布有极大的相关性。首先可以清楚地看到,A类群较为独立于其他三个类群,该类群所有分布点属于日本诸岛屿,其典型的岛屿海洋性气候与中国内地的大陆性气候有较大的区别;其次C、D类群是较为分散的类群,该类群主要包括的分布点是云南、贵州、广西西南部、陕西、甘肃、湖南西部以及湖北西北部,这些地区大多属于云贵高原边缘带或是秦岭-大巴山一带地区,该地区降雨量较东部有所减少、温度指标有所降低、海拔高度抬升,因此形成了西部类群,并且高原性气候与我国其他地方的气候特点也有较大的区别;再次B类群主要为东部地区分布点,集中了浙江、安徽、福建、江西等部分地区,该类群分布区气候最大特点为降雨量有较大的提高,并且其热量指数等也有所增高,平均分布海拔高度有所降低;最后为E类群,其主要为中国南部地区分布点,包括广东、广西西北部、湖南南部等。通过这4个类群的划分可以知道,热量指标是影响乌冈栎分布区的最主要因素。
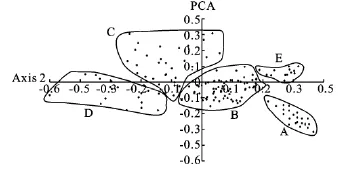
图4 气象指数两维主分量排序图
2.5 乌冈栎生物地理模型
大尺度的植被分布与气候之间存在着某种对应关系,特定的生物气候特征必然导致特定的植被类型[23],因此模型研究是气候植被研究的重要手段[24]。乌冈栎分布区气候因子主成分分析得知,温度是影响乌冈栎分布的最主要因素,因此模型采用一月均温(TM-Jan)、七月均温(TM-Jul)、年均温(TM-A)等温度指标与纬度、经度、海拔的关系,应用多对多回归方法建立乌冈栎地理分布模型,模拟预测乌冈栎地理分布(表3)。
从表3可知,通过逐步线性回归,剔除相关性不大的因子得到相应的回归方程中:经度与温度并没有太大的相关性,在方程1,2,3中均没有经度因子;海拔也不是限制其分布的主要因子,它在各方程中比重都较轻(系数0.002~0.02);纬度的影响最大,它直接影响了地球热量的分布。此外,方程3的R2=0.583,说明七月均温对乌冈栎的分布影响不大,因此在使用时可以不加考虑。以已知引种成功的南京(N=32°,E=118.8°,H=7m)作为模型检验,其结果分别为TM-A=16.928℃、TM-Jan=6.537,与文献记载的气象数据TM-A=15.9℃、TM-Jan=3相接近。模拟的结果与实际值存在一定的偏差,这是因为一方面仅能利用现有的地理分布数据,而许多未被记录的地理分布数据无法收集;另一方面,有地理分布数据的并不一定有气象记录观察点,因此只能利用空间插值法获取数据。但地理分布模型仍具有一定的参考价值,只是在使用时结合当地实际情况加以修正即可。
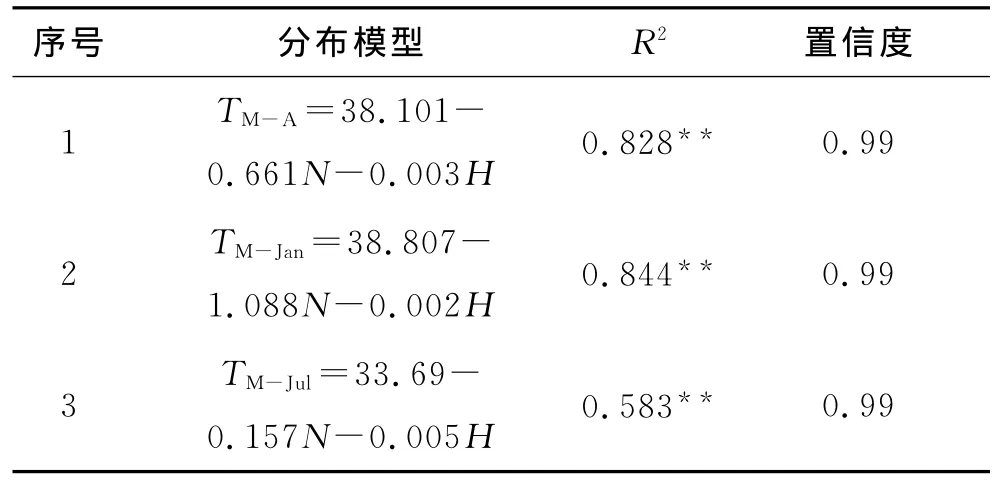
表3 乌冈栎地理分布模型
3 结论
气候对生物的影响极为深刻,不仅直接限制生物的生活和分布,而且也通过其他环境因子对生物产生间接影响,这些因子并不是单独或孤立地起作用,而是作为一个整体相互制约地对生物产生影响,从而使地球表层形成了不同的生物群[25]。通过温暖指数(WI)、寒冷指数(CI)、干燥度指数(K)、水热综合指数(S)、年均生物温度(BT)等13项气候指数的研究,对乌冈栎分布区生态适应性有了清晰的认识。东亚地区乌冈栎分布大致主要集中在中国秦岭以南至北回归线的范围(23°-34°N),日本集中分布在琉球群岛、九州岛、四国岛和本州岛的海岸沿线与岛中高山地带(26°-37°N)。通过环境指数的分析,乌冈栎属于亚热带树种,其分布区内的水热综合指数S=115.772、年均生物温度BT=17.85、年均可能蒸散发率PER=0.728等指标与青冈栎较为接近,因此该树种亦为耐寒树种。将中国内地分布区与日本诸岛热量分布曲线比较发现,中国热量分布曲线宽度要大于日本热量分布曲线,说明中国乌冈栎分布区类型要多于日本。主成分分析的结果表明温度是影响乌冈栎分布区的主导因子,由此可将乌冈栎分布区划分为5个类型,包括:日本区(A)、中国华东区(B)中国西南部区(C)、中国西北部区(D)和中国南部区(E)(图5)。以基本的温度指标为因变量,以经度、纬度和海拔为自变量,建立乌冈栎地理分布模型,取得了一定的预测效果;另外,模型本身也说明了乌冈栎分布区受经度、海拔的影响较小。
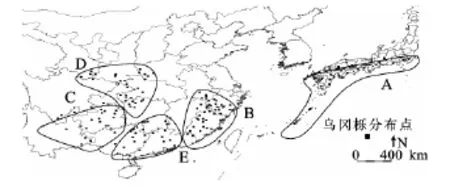
图5 乌冈栎分布区地理区划
任何一物种的分布都与其分布区环境有着密切的关系。东亚地区仅在中国和日本有乌冈栎的分布,并且两地乌冈栎都能形成纯林,那么乌冈栎的岛屿分布与大陆分布在大的气候环境上有何相似之处呢?中国乌冈栎分布区为典型的亚热分布区,其气候属于亚热带季风性湿润气候,体现在冬季温和少雨、夏季炎热多雨、雨热集中;而乌冈栎在日本的分布地的气候可以囊括为中央高地气候、太平洋侧气候和西南诸岛气候,这些气候中中央高地气候为典型的内陆性气候、太平洋侧气候为梅雨强台风多、南西诸岛气候为夏季炎热冬季温暖降雨量大。通过比较不难看出,日本西南诸岛气候与我国的华东区气候较为接近,太平洋侧气候又与长江流域气候相似,而中中央高地气候则接近于西部云贵高原一带;同时,由于冬季的低温,中国亚热带所处的纬度偏南,其北界比理论上的界线南移4~5个纬度。因此,乌冈栎在大陆和岛屿两种大的格局生境中的良好分布得到了清楚的解释。
[1] 王婧,王少波,康宏樟,等.东亚地区栓皮栎的地理分布格局及其气候特征[J].上海交通大学学报:农业科学版,2009,27(3):235-240.
[2] 余振,孙鹏森,刘世荣.中国东部南北样带主要植被类型物候期的变化[J].植物生态学报,2010,34(3):316-329.
[3] Cox C B,Moore P D.Biogeography an Ecological and Evolutionary Approach[M].6th ed.Oxford:Blackwell Science Ltd,2000.
[4] 柯文山,钟章成,杨毅,等.四川大头茶地理分布与环境因子的关系[J].生态学杂志,1999,18(6):24-27.
[5] 倪健,宋永昌.中国亚热带常绿阔叶林优势种及常见种的分布与 Kira指标的关系[J].生态学报,1997,18(3):248-262.
[6] Yim Y J.Distribution of forest vegetation and climate in the Korean peninsula.Ⅲ.Distribution of tree species along the thermal gradient[J].Japanese Journal of Ecology,1977,27(3):177-189.
[7] Federici F,Pignatii S.The warmth index of Kira for the interpretation of vegetation belts in Italy and SW.Australia two regions with Mediterranean type bioclimates[J].Vegetatio,1990,93:91-99.
[8] 倪健,宋永昌.中国青冈的地理分布与气候的关系关[J].植物学报,1997,39(5):451-460.
[9] 苏小青,陈世品,童建宁.福建主要树种的分布与气候条件关系的研究[J].福建林学院学报,2001,21(4):371-375.
[10] 苏建荣,张志钧,邓疆.云南红豆杉的地理分布与气候关系[J].林业科学研究,2005,18(5):510-515.
[11] 李峰,周广胜,曹铭昌.兴安落叶松地理分布对气候变化响应的模拟[J].应用生态学报,2006,17(12):2255-2260.
[12] Hijmans R J,Cameron S E,Parra J L,et al.Very high resolution interpolated climate surfaces for global land areas[J].International Journal of Climatology,2005,25:1965-1978.
[13] Hara M,Ozaki K,Isoya T.Habitat and Distribution of Quercus phillyraeoides Trees at its Northeastern Distribution Limit,Kyonan-machi,Boso Peninsula,Central Japan[J].Journal of the Natural History Mu-seum and Institute,Chiba,2005,8(2):11-16.
[14] Miyawaki A.Outline of Japanese Vesgtation.JIBP Synthesis,Studies in Conservation of Natural Terrestrisal Ecosystems in Japan,Part 1[J].Vegetation and Its Conservation,1975,8:19-28.
[15] Numata,M.The Flora and Vegetation of Japan[M].Kodansha Limited Elsevier Scientifie Publishing Company,1974.
[16] Yoshiwo Horikawa.Atlas of the Japanese Flora I:an introduction to plant sociology of East Asia[M].Japan Tokyo:Gakken Co.,Ltd.,1972.
[17] 翁恩生,周广胜.用于全球变化研究的中国植物功能型划分[J].植物生态学报,2005,29(1):81-97.
[18] 蒋霞,倪健.西北干旱区10种荒漠植物地理分布与大气候的关系及其可能潜在分布区的估测[J].植物生态学报,2005,29(1):98-107.
[19] 张新时.研究全球变化的植被-气候分类系统[J].第四纪研究,1993,5(2):157-169.
[20] 洪必恭,李绍珠.江苏主要常绿阔叶树种的分布与热量关系的初步研究[J].生态学报,1981,1(2):105-111.
[21] 徐文铎.东北主要树种的分布与热量关系的初步研究[J].东北林学院学报,1982(4):1-9.
[22] 王贤荣,谢春平,伊贤贵,等.不同居群野生早樱形态变异研究[J].植物研究,2007,27(6):746-752.
[23] 李双成.植物响应气候变化模型模拟研究进展[J].地球科学进展,2001,20(3):217-226.
[24] 赵东升,李双成,吴绍洪.青藏高原的气候植被模型研究进展[J].地球科学进展,2006,25(4):68-78.
[25] 殷秀琴.生物地理学[M].北京:高等教育出版社,2004.
猜你喜欢
野生动物学报(2024年4期)2024-11-28 00:00:00
疯狂英语·新读写(2021年8期)2021-11-05 08:44:26
安徽农业科学(2021年14期)2021-09-03 09:55:59
炎黄地理(2019年5期)2019-10-20 01:59:19
小学生优秀作文(高年级)(2018年4期)2018-09-11 01:23:24
少儿科学周刊·儿童版(2018年12期)2018-01-26 12:02:22
少儿科学周刊·少年版(2018年12期)2018-01-26 12:01:02
安徽农业科学(2016年9期)2016-10-14 22:12:23
少儿科学周刊·儿童版(2016年1期)2016-03-14 04:00:33
山东青年(2016年1期)2016-02-28 14:25:28
