
PEG胁迫对沙冬青种子萌发过程的影响
2011-05-07 11:06:46段慧荣李毅马彦军
水土保持研究 2011年3期
段慧荣,李毅,马彦军
(甘肃农业大学林学院,兰州730070)
在限制植物产量的诸多环境因子中,以供水条件最为重要,据统计,全世界由于水分胁迫导致的作物减产,可超过其它因素造成减产的总和。在干旱半干旱地区,水的短缺始终是制约农林业生产的关键因素[1]。在干旱半干旱地区,通过植树造林,改善生态环境工作已成为当务之急,而木本植物抗旱性研究是该区造林树种选择的基础,是关系到该区造林成败的关键因素之一[2]。
种子是植物最重要的繁殖材料,种子萌发是种子植物生活史中的关键阶段,也是进行植物抗旱性研究的重要时期[3]。研究表明,在植物生活周期中,植物死亡至少有95%发生在种子阶段[4]。在该阶段,土壤水分是影响种子萌发及幼苗生长的重要生态因子。发芽阶段的耐旱程度一定程度上反映了该物种的耐旱状况。种子发芽状况也是判定种子质量、确定播种量的一项重要指标。近年来,PEG高渗溶液法已经成为研究种子萌发性状的重要方法,在水稻、小麦等农作物上广泛使用[5],在牧草上种子抗逆性研究方面也开始逐渐增多[6-7],而在林木种子的抗逆性研究方面相对比较少[8]。
沙冬青(Ammopiptanthus mongolicus)为亚洲中部荒漠地区唯一的常绿阔叶灌木,是古老的第三纪孑遗种,也是国家三级保护植物。其抗逆能力极强,但是人工栽培却极为困难,在保护和开发利用沙冬青的研究中,探索沙冬青的抗逆机理及其最佳繁殖栽培技术一直是人们关注的焦点。目前,有关沙冬青耐旱性研究已有一些报道,最早见于对其形态解剖特征的观察[9],但是有关沙冬青对PEG渗透胁迫的生理响应研究报道较少。基于以上认识,本试验以PEG-6000溶液模拟干旱胁迫,研究5种不同PEG浓度(5%~25%)对沙冬青种子萌发的影响及萌发幼苗细胞膜透性、可溶性蛋白、丙二醛和脯氨酸含量、超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxide,POD)和过氧化氢酶(Catalase,CAT)等各项生理指标的变化,以期系统了解沙冬青种子萌发阶段的耐旱性特征并揭示其抗旱机理,为沙冬青在干旱区播种提供理论依据。
1 材料和方法
1.1 材料
沙冬青成熟荚果于2009年8月初采自甘肃省兰州市九州台,千粒重为55.1 g。选取健壮饱满、无损伤和无虫害的灰黑色种子,用75%的酒精消毒1~2 min后,用自来水和蒸馏水冲洗干净,浸泡在100℃水5 min后,用蒸馏水冲洗冷却降温,放置24 h后进行PEG-6000胁迫处理。
1.2 试验设计
设质量浓度 5%、10%、15%、20%和 25%,对应渗透势分别为-0.054,-0.177,-0.393,-0.735,-1.25 MPa共5个不同渗透势浓度的PEG(中国医药(集团)上海化学试剂公司)溶液,按照李合生[10]的方法配备,对上述沙冬青种子进行干旱胁迫处理。处理采用纸上发芽,首先在培养皿内铺2层滤纸,将处理好的种子(大小一致无破损)置于滤纸上,然后加入等量的PEG溶液进行萌发,以加蒸馏水为对照。每个处理3次重复,每个重复50粒种子,室温下发芽,于2009年10月份在甘肃农业大学林学院实验室进行。培养过程中每天向培养皿中加入相应处理溶液,保持滤纸湿润。每3天更换1次滤纸,以防止溶液浓度变动[11]。培养条件为:14 h光照,光照强度2 000 lx,昼温20~25℃/夜温15~20℃。
1.3 测定方法
1.3.1 种子发芽率和死亡率的测定 从种子置床之日起开始观察,以种子露白为发芽标准[12],逐日定时记录发芽种子数,30 d后结束发芽试验。参照《种子生物学研究指南》[13],计算发芽率和死亡率。其中死亡幼苗以种子萌发后不再生长甚至腐烂为判断标准。
发芽率=种子发芽数/供试种子数×100%
死亡率=死亡苗数/总苗数×100%
1.3.2 生理生化指标 在室内用电子天平(精度0.1 mg)称取整株幼苗的鲜重[14],在80℃烘箱内烘干48 h,称取干重[15],发芽结束时自每个处理中分别选取同一生长时期发育良好的健康植株整株进行生理生化指标的测定,每项实验重复3次,取平均值。细胞膜透性的测定采用相对电导率法,可溶性蛋白的含量测定采用G-250考马斯亮蓝法,丙二醛(MDA)含量的测定采用硫代巴比妥酸法。超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)光还原法,以抑制氯化硝基氮蓝四唑(NBT)光化还原50%为1个酶活性单位;过氧化物酶(POD)活性用愈创木酚比色法,在470 nm下测定,以每克植物鲜重中每分钟所含酶活性单位(U)表示;过氧化氢酶(CAT)活性的测定采用过氧化氢分解法,脯氨酸(Proline)含量的测定采用酸式茚三酮显色法[14],显色后在UV-2450分光光度计520 nm波长处测定光密度值,通过标准曲线计算游离脯氨酸含量。
试验数据采用Excel 2003和SPSS 16.0软件进行作图和数据分析。
2 结果与分析
2.1 PEG胁迫对沙冬青种子萌发过程的影响
不同浓度PEG胁迫对沙冬青种子萌发的影响见图1。5%和10%PEG浓度下种子发芽率及萌发幼苗鲜重与对照无明显差异,死亡率低。PEG浓度为15%时种子死亡率为3.33%,萌发幼苗鲜重比对照降低了33.03%,差异显著(P<0.05);说明PEG浓度达到 15%时开始抑制种子萌发生长,并且随着PEG浓度的升高种子所受到的抑制程度也越高。当PEG浓度达到25%时,萌发幼苗变得矮小枯黄,鲜重降至0.116 g,极显著地低于对照(P<0.01),发芽率为2%,死亡率达到98.89%。
2.2 PEG胁迫下沙冬青萌发幼苗生理指标的变化
2.2.1 相对电导率和丙二醛含量 由图2可以看出,随着PEG浓度的增加,沙冬青萌发幼苗的相对电导率和丙二醛的含量均呈现上升的趋势。PEG浓度在5%~10%时沙冬青的电导率与对照变化不大,无明显差异。当浓度达到15%~25%时,电导率呈上升趋势,并且均与对照有极显著差异(P<0.01)。丙二醛含量随PEG浓度增大而增加,且当浓度达到15%~25%时,各含量均与对照呈极显著差异(P<0.01),分别比对照上升了162.63%、182.36%和188%。
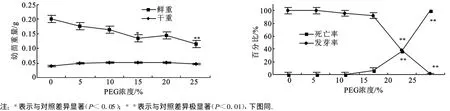
图1 PEG胁迫对沙冬青种子萌发过程的影响

图2 PEG胁迫对沙冬青萌发幼苗相对电导率和丙二醛含量的影响
2.2.2 可溶性蛋白含量 可溶性蛋白具有较强的亲水胶体性质,可影响细胞的保水力。随PEG浓度增大,可溶性蛋白含量呈现先上升后下降的趋势。PEG浓度为5%~15%时,含量随浓度增大而上升,与对照无明显差异,浓度继续增大至20%和25%时开始下降,并分别比对照降低22.37%和32.89%,与对照差异显著(P<0.05),见图3。
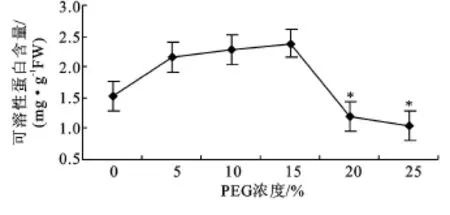
图3 PEG胁迫对沙冬青萌发幼苗可溶性蛋白含量的影响
2.3 PEG胁迫对幼苗保护性酶活性的影响
2.3.1 超氧化物歧化酶(SOD)活性 SOD的主要功能是清除氧自由基,是防止其对细胞膜系统伤害的一种很重要的抗氧化酶。由图4可见,PEG胁迫对幼苗SOD活性的影响整体表现出先上升后下降的趋势。其中5%~15%PEG浓度培养下,幼苗SOD活性随着PEG浓度的增加而增大,分别比对照上升了20.27%、31.38%和65.53%,均与对照呈极显著差异(P<0.05)。PEG浓度为20%时活性下降,仅比对照增加了2.86%,无显著性差异,说明此时抗性减弱。当浓度增加到25%时,酶活性比对照降低了26.27%,与对照呈极显著差异。
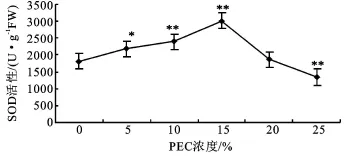
图4 PEG胁迫对沙冬青萌发幼苗SOD活性的影响
2.3.2 过氧化物酶(POD)活性 POD在植物体内具有广泛的作用,其主要作用之一是催化H2O2降解。PEG胁迫对沙冬青幼苗POD活性的影响与对SOD活性影响相似,都表现出先上升后下降的趋势。如图5所示,PEG浓度为5%时,幼苗POD活性比对照上升了50.96%,与对照有极显著差异(P<0.01)。10%PEG浓度下,幼苗的POD酶活性最高,与对照相比上升了76.38%。当PEG浓度达到15%~20%时,幼苗 POD活性开始降低并分别低于对照27.67%、56.32%,其中浓度为15%时与对照差异不显著。幼苗受到浓度为25%PEG胁迫时POD活性最小,比对照下降了73.48%,与对照差异极显著。
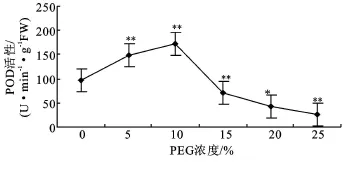
图5 PEG胁迫对沙冬青萌发幼苗POD活性的影响
2.3.3 过氧化氢酶(CAT)活性 图6表明,各浓度PEG胁迫下CAT酶活性表现为先上升后下降的变化趋势,与对照之间呈现极显著差异(P<0.01)。5%~10%PEG浓度下酶活性呈上升趋势,分别比对照上升了68.14%和136.38%。当PEG浓度达到15%时,酶的活性开始下降,但仍比对照高出81.06%,浓度为20%~25%时,酶活性比对照分别下降14.23%和63.39%。
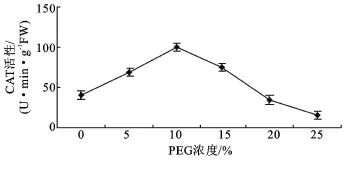
图6 PEG胁迫对沙冬青萌发幼苗CAT活性的影响
2.4 PEG胁迫对幼苗脯氨酸含量的影响
脯氨酸含量的提高是逆境条件下植物的自卫反应之一,细胞脯氨酸含量增加,维持了细胞的膨压,同时可以保护酶和膜系统免受毒害。由图7看出,PEG浓度低于20%时,随浓度的增加沙冬青幼苗的脯氨酸含量升高,说明5%~20%的PEG可以刺激幼苗脯氨酸含量极显著地上升(P<0.01),并分别比对照增加了 164.37%、293.38%、313.7%和 332.03%。当浓度达到25%时,脯氨酸含量迅速下降,并比对照下降了48.57%,与对照之间差异不明显。
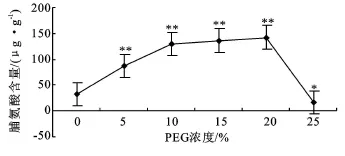
图7 PEG胁迫对萌发幼苗脯氨酸含量的影响
3 讨论
3.1 PEG胁迫对种子萌发的影响
用一定浓度的聚乙二醇(PEG)溶液模拟干旱逆境处理种子,使之产生渗透胁迫来研究水分胁迫下种子的生理反应,已有较多报道。种子吸水力与植物的抗旱性呈正相关,吸水力强的种子在干旱胁迫下能够保持较高的发芽率和发芽势,而吸水力弱的种子则相反[16]。本试验中,随胁迫程度的加深,沙冬青种子发芽率显著下降,死亡率呈明显上升趋势,鲜重下降。表明随胁迫程度的加深,沙冬青种子萌发受到不同程度的抑制,当胁迫超过种子的耐受限度(25%)时将难以萌发并致死,这与董丽华[17]等的研究结果一致。
3.2 PEG胁迫对萌发幼苗生理生化指标的影响
细胞膜是植物体内部与外部物质交换的通道,多种逆境都会使植物细胞膜系统受损,导致膜透性增大,内容物渗漏。植物组织外渗液电导率可以表征质膜透性的变化并反映胁迫条件下质膜受损伤的程度,电导率增幅越大,植物受胁迫损伤程度越重。丙二醛(MDA)是植物在受到伤害时细胞膜发生膜脂过氧化作用形成的最终分解产物,其含量代表着植物细胞遭受逆境伤害的程度和膜脂过氧化程度[18]。本试验中,随胁迫程度的加深,沙冬青萌发幼苗的电导率不断上升,同时MDA含量也不断增加,表明随胁迫程度加深,萌发幼苗的膜系统受到不同程度的破坏,膜质过氧化作用增大,抗性减弱,这与王宇超等[18]的研究结果一致。
逆境和胁迫会导致植物体内大量活性氧的产生,SOD作为重要的活性氧清除酶,能及时清除体内产生的活性氧将超氧阴离子自由基快速歧化为H2O2和H2O,保护细胞免受伤害[19]。POD和CAT是清除H2O2的重要酶类,能够分解H2O2为H2O和O2,CAT与超氧化物歧化酶SOD偶联,可以彻底清除体内超氧阴离子及H2O2等氧自由基,消除逆境对植物的伤害。试验中随胁迫程度的加深,SOD、POD和CAT活性均呈现先升高后降低的趋势,这表明在一定范围(5%~10%)内,PEG胁迫能诱导体内保护酶活性提高,从而有效地清除活性氧,但严重胁迫则会破坏体内保护酶系统的活力和平衡,损伤膜的结构并抑制酶的活性。这与左利萍等[20]在柠条叶片的抗氧化酶活性影响的结果一致。
蛋白质是生命活动的体现者,植物不同其蛋白质的种类和数量也不同。植物体内的可溶性蛋白质大多是参与各种代谢的酶类,在受到干旱胁迫时,它们会发生一定的变化,测定其含量是了解植物抗逆性的一个重要指标[21]。试验中,沙冬青萌发幼苗中可溶性蛋白的含量随胁迫程度加深先增加后降低,说明通过生成蛋白为抵抗胁迫环境提供物质基础。研究中,当PEG浓度在20%以上时,可溶性蛋白的含量明显低于对照,说明此胁迫已超过沙冬青的耐受极限,蛋白质水解加快。游离脯氨酸是作为渗透调节物质和防脱水剂而起作用的,可以降低细胞水势和维持膨压,脯氨酸除了发挥其本身的渗透、保护作用外,又为新的蛋白质的合成提供了氨基,从而促进了与抗旱有关的新蛋白质的合成。研究表明,在干旱或其它逆境条件中植物体内脯氨酸含量会大量积累。本试验结果表明,沙冬青萌发幼苗中脯氨酸含量随胁迫程度加深先增大后降低,表明脯氨酸作为细胞内的渗透调节剂,在渗透胁迫较轻时可以有效地平衡液泡渗透势,保护生物大分子不被破坏,作为活性氧的清除剂和抗冷剂抵抗渗透胁迫。胁迫浓度超过25%时,细胞被破坏,导致脯氨酸含量反而降低。
由于本次试验只是对PEG模拟干旱胁迫下沙冬青种子萌发进行了研究,认为沙冬青可以适应PEG浓度15%以下的干旱胁迫环境,至于后期生长过程中的幼苗形态和生理生化变化,还有待进一步研究和探讨。
4 结论
试验结果表明,沙冬青种子在PEG浓度为5%~10%时能够萌发生长,通过增加渗透调节物质如可溶性蛋白和脯氨酸的含量,提高保护性酶SOD、POD与CAT的活性来抵抗胁迫环境。其能够忍受PEG浓度15%的干旱胁迫环境,但已经开始出现不适现象,在高浓度(20%和25%)时保护作用被抑制,种子基本不能萌发,萌发幼苗矮小枯黄,死亡率上升,抗性下降。
[1] 邓洁,陈静,贺康宁.灌水量和灌水时期对冬小麦耗水特性和生理特性的影响[J].水土保持研究,2009,16(2):191-194.
[2] 崔大练,马玉心,石戈,等.紫穗槐幼苗叶片对不同干旱梯度胁迫的生理生态响应[J].水土保持研究,2010,17(2):178-185.
[3] 李培英,孙宗玖,阿不来提.PEG模拟干旱胁迫下29份偃麦草种质种子萌发期抗旱性评价[J].中国草地学报,2010,32(1):32-39.
[4] 班勇.植物生活史对策的进化[J].生态学杂志,1995,14(3):33-39.
[5] 安永平,强爱玲,张媛媛,等.渗透胁迫下水稻种子萌发特性及抗旱性鉴定指标研究[J].植物遗传资源学报,2006,7(4):421-426.
[6] 贾国梅,张红燕,韩京成.土壤含水量对狗牙根叶片生理生态指标的影响[J].水土保持研究,2009,16(5):199-202.
[7] 左胜鹏,王会梅,李凤民,等.半干旱区四种典型豆科牧草群落生理生态的分异研究[J].水土保持研究,2010,17(1):167-173.
[8] 曾彦军,王彦荣.几种旱生灌木种子萌发对干旱胁迫的响应[J].生态学报,2002,13(8):953-956.
[9] 张涛.沙冬青生理结构特性的研究[J].林业科学,1988,24(4):508-509.
[10] 李合生.植物生理生化试验原理和技术[M].北京:高等教育出版社,2000:260-265.
[11] 宋丽华,周月君.盐胁迫对臭椿种子发芽的影响[J].种子,2008,27(9):22-25.
[12] 秦文静,梁宗锁.四种豆科牧草萌发期对干旱胁迫的响应及抗旱性评价[J].草业学报,2010,19(4):61-70.
[13] 宋松泉,程红焱,龙春林,等.种子生物学研究指南[M].北京:科学出版社,2000.
[14] 郑光华.种子生理研究[M].北京:科学出版社,2004:706.
[15] 杜建雄,侯向阳,刘金荣.草地早熟禾对干旱及旱后复水的生理响应研究[J].草业学报2010,19(2):31-38.
[16] 郭彦军,倪郁,吕俊,等.豆科牧草种子萌发特性与其抗旱性差异的研究[J].中国草地,2003,25(3):24-27.
[17] 董丽华,王宁,姚爱兴.不同品种早熟禾种子萌发期抗旱性研究[J].农业科学研究,2005,26(2):6-8.
[18] 王宇超,王得祥,彭少兵,等.干旱胁迫对木本滨藜生理特性的影响[J].林业科学,2010,46(1):61-67.
[19] 安钰,沈应柏,张志翔.伤害胁迫对合作杨叶片膜质过氧化及抗氧化酶活性的影响[J].水土保持研究,2010,17(5):241-244.
[20] 左利萍,李毅,李朝周.渗透胁迫对柠条叶片脂质过氧化及抗氧化酶活性的影响[J].中国草地学报,2009,31(4):69-73.
[21] 史玉炜,王燕凌,李文兵,等.水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响[J].新疆农业大学学报,2007,30(2):5-8.
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
中华建设(2020年5期)2020-07-24 08:55:14
科学与财富(2019年15期)2019-10-21 13:35:48
小学生必读(低年级版)(2018年6期)2018-09-08 07:44:36
中成药(2018年8期)2018-08-29 01:28:34
发酵科技通讯(2018年2期)2018-07-06 11:38:46
——致坚守奋斗的你
音乐天地(音乐创作版)(2017年1期)2017-02-12 18:58:38
湖南林业科技(2017年1期)2017-02-06 05:29:00
医学研究杂志(2015年9期)2015-07-01 17:28:24
中央民族大学学报(自然科学版)(2015年3期)2015-06-11 02:13:46
