
东亚飞蝗染色体C带和银染带型分析
2011-04-25 09:46吴学芳何正波陶红梅蔡志华
草业科学 2011年5期
吴学芳,何正波,陶红梅,蔡志华
(1.重庆师范大学生命科学学院动物学重点实验室,重庆 400047; 2.重庆师范大学资源昆虫和分子生物研究所,重庆 400047)
近年来,我国蝗虫灾害频繁爆发,严重影响了天然草地植被的正常生长发育[1],蝗虫危害面积占草原虫害危害面积的52.4%[2]。直翅目昆虫东亚飞蝗(Locustamigratoriamanilensis)是一种洲际性农业害虫,也是引发我国蝗灾的最主要害虫。目前,蝗总科的染色体已有不少的研究,但东亚飞蝗的染色体研究尚少。对东亚飞蝗染色体的研究将为生物治蝗奠定基础[3]。蝗虫染色体的研究普遍采用压片染色显带,进行C带和银染核仁组织区(Ag-nucleolus organizer region,Ag-NOR)定位研究。本试验采用哺乳动物常用的悬浮滴片法,研究了东亚飞蝗C带和银染显带,并对其带型进行分析。
1 材料与方法
本研究所用东亚飞蝗由重庆师范大学生命科学学院资源昆虫分子生物研究所提供。
1.1染色体标本制备 根据陶红梅等[4]滴片法略做修改。东亚飞蝗活体注射0.04%秋水仙素溶液0.3 μL,8 h后活体解剖取出精巢。于0.75%的生理盐水中剔净脂肪和筋膜后,放入37℃预温的1%柠檬酸钠溶液中低渗30 min[5]。转至新鲜配制的3∶1甲醇冰醋酸固定液中30 min,换固定液重复固定数次,直至精巢发白,取出剪碎,新鲜固定液悬浮固定,1 000 r/min离心,弃上清液,反复4次,滴片。
1.2C带 C带处理采用BSG法,具体过程参照Summer[6]方法稍加修改,玻片65℃烘箱内老化24 h,0.2 mol/L盐酸溶液室温处理1 h,60℃氢氧化钡110 s,2×SSC在60℃恒温水浴锅内处理60 min。10%Gimesa染色8 min,镜检。
1.3银染 按Howell和Black[7]的方法,略做改进。玻片于65℃水浴,加50%硝酸银溶液4滴, 2%明胶显影液2滴,覆以盖片,直至玻片标本呈茶褐色(一般为3 min)。蒸馏水冲洗,10% Gimesa复染8 min,镜检。
1.4染色体核型 取30只蝗虫按照上述方法制得染色体玻片标本,每个个体选取10个分散较好的分裂中期细胞显微照相,冲洗放大。用Image-Pro Plus 5.1软件测量照片上染色体和C带带纹长度等相关数据,并对染色体进行分组、编号,分析核型、C带和银染带型,计算染色体的相对长度和异染色质含量[8]。
2 结果与分析
2.1核型 东亚飞蝗的染色体数目为2N(♂)=23。性别决定机制为XO。染色体组式为4L+4M+3S+XO,包括大型染色体4对,相对长度RL值13.99~11.11;中型染色体4对,相对长度RL值8.16~5.16;小型染色体3对,相对长度RL值3.22~2.44;X染色体为大型染色体,RL值17.37,染色体长度在整个染色体组中位居第一(表1)。本试验显示的染色体核型和组式与贾潇凌和马恩波[9]、马恩波和欧晓红[10]的研究结果一致,可确定此为东亚飞蝗的基本核型。
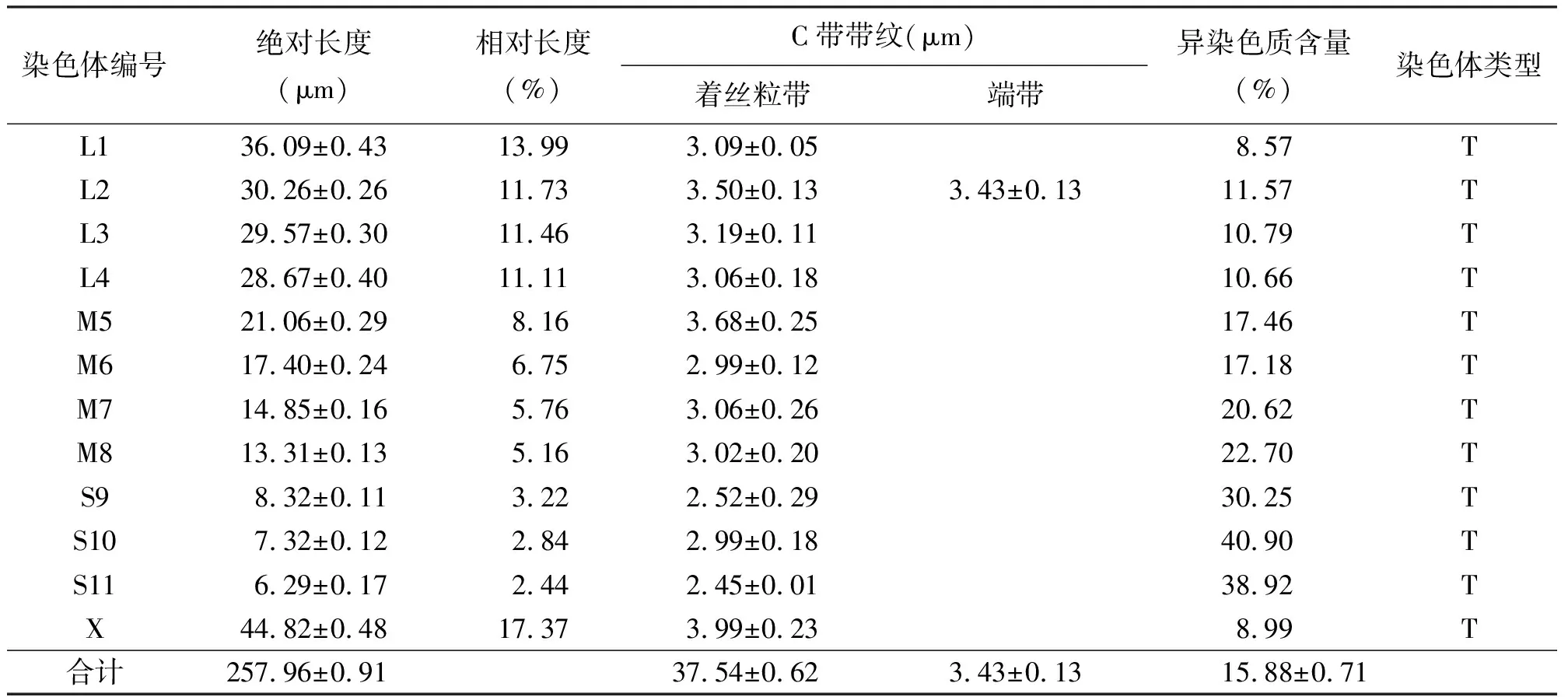
表1 东亚飞蝗染色体和C带数据统计
2.2C带 东亚飞蝗所有染色体具有明显的端着丝粒带,与已进行染色体研究的大多数蝗总科种类一致。早前期细胞着丝粒深染点附着在细胞核膜上,减数分裂Ⅰ和减数分裂Ⅱ中期染色体都具有着丝粒C带。其中,第1、4、7对和X染色体着丝粒较浅,仅隐约可见;第2、3、5、6、9、10、11对染色体着丝粒呈现大小不同的深染区;第8对染色体着丝粒为中度着色,第2对染色体端粒深染;未见居间带(图1)。染色体组中异染色质总含量为(15.88±0.71)%。
2.3银染 间期细胞银染深染点为3~6个不等,多为5个,有的深染点紧贴于细胞核膜上。减数分裂Ⅰ前期观察到黑色团块Ag-NOR,多为3个。减数分裂Ⅰ中期,观察到2号染色体近端部次缢痕处呈较规则圆形Ag-NOR,较为恒定。极少数细胞除2号染色体的深染点外,1号染色体近着丝粒初缢痕处有形状不规则Ag-NOR。 其他各期未发现明显的Ag-NOR(图2)。
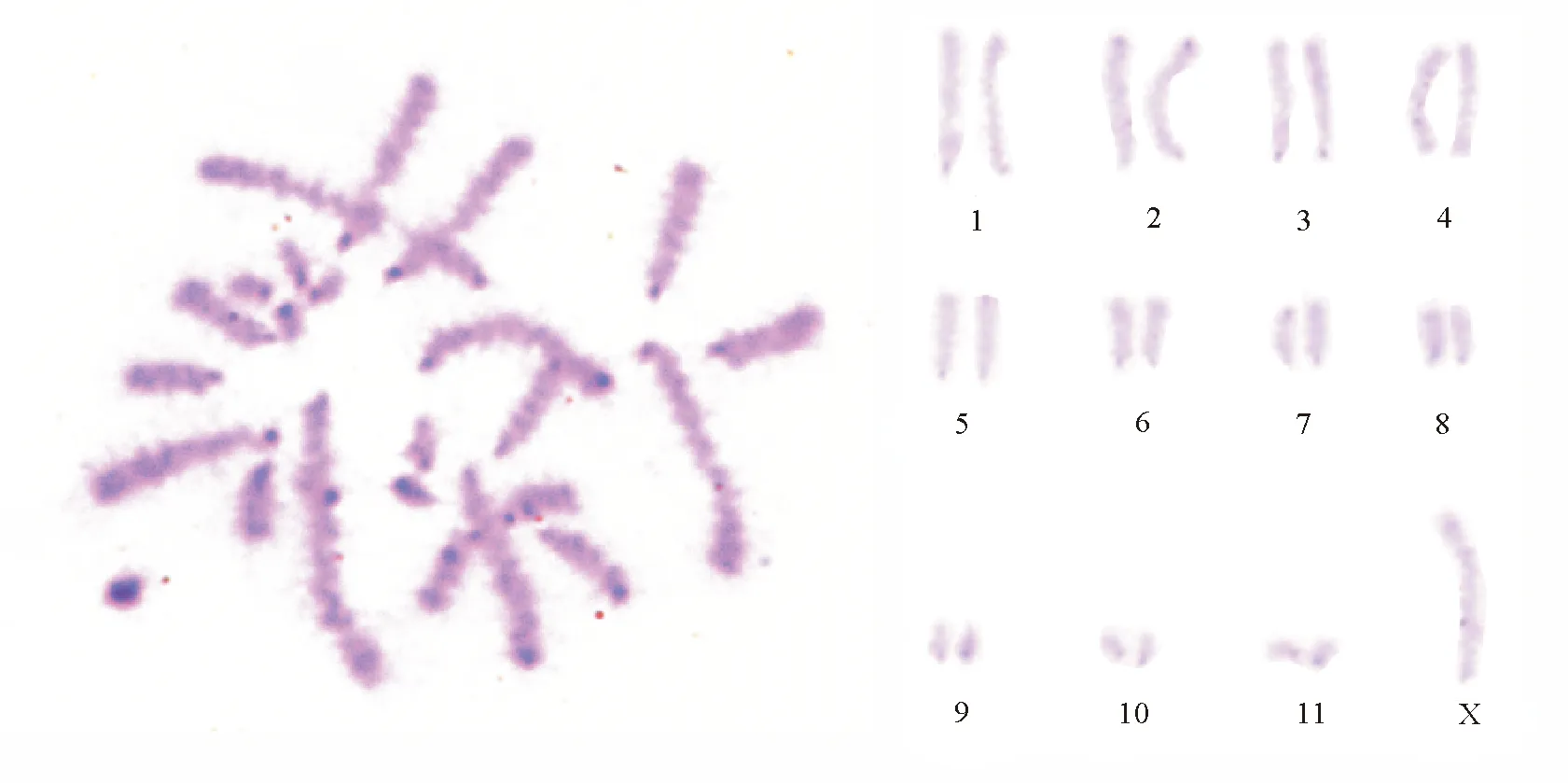
图1 东亚飞蝗中期染色体C带及排序
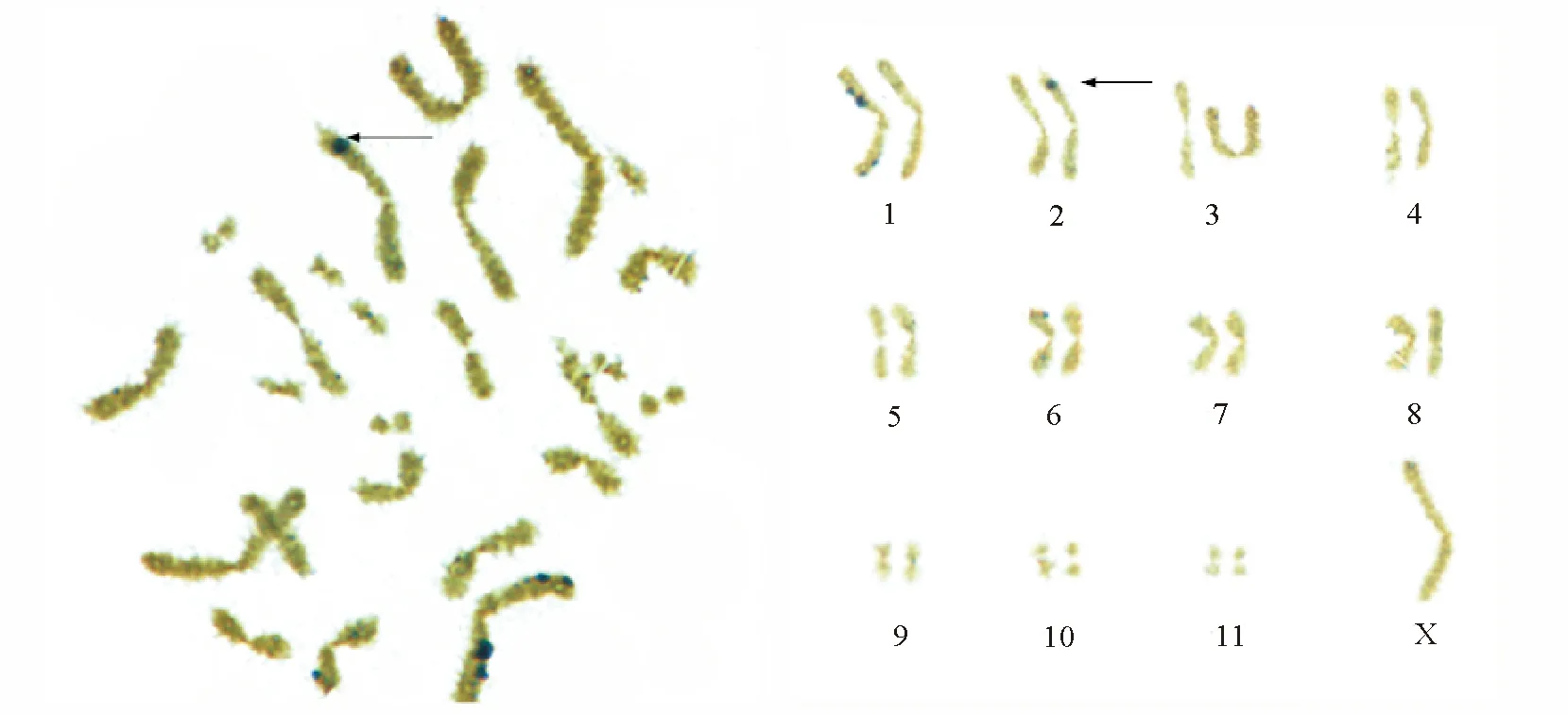
图2 东亚飞蝗中期银染及排序
3 讨论与结论
以往蝗虫染色体研究多采用冰冻揭片法或压片法,本试验采用改进后的悬浮滴片法[4],玻片较其他方法更为干净,染色体形态良好,细胞质抽提干净,利于观察和显带研究。
制片过程中,秋水仙素的注射量和低渗时间是影响东亚飞蝗染色体标本形态主要因素。秋水仙素注射量过大,染色体凝缩,甚至可呈点状;注射量过小,则无法起到阻断细胞分裂的作用,中期细胞少。经反复试验,每只虫体注射3 μL 0.04%秋水仙素,处理8 h,效果最好。低渗时间也会影响染色体形态。低渗时间过短,染色体伸展不好,呈紧缩状;时间过长,染色体疏松,崩解,甚至出现灯刷染色状外观,若继续低渗会导致细胞破裂,染色体丢失。1%柠檬酸钠溶液低渗30 min可达到较好的效果。
3.1C带 近年来,人们利用分子生物学研究手段证实C带主要是AT与GC高度重复序列[11]。C带标本中深染部分即为结构异染色质部分,着色深浅程度与异染色质含量高低有直接联系,可因此进行种间的比较。根据Webb[12]的观点,深黑色为结构异染色质密集区域,灰黑色带则为结构异染色质与常染色体夹杂的区域。第2对染色体端粒处出现深染,说明较其他染色体,该染色体端粒处结构紧密且非组蛋白含量也更为丰富。东亚飞蝗均为端部着丝粒染色体,且某些染色体出现了端带,而以往研究的大多数蝗总科[9,12-15]的种类也出现了此特点。对比发现,东亚飞蝗与其他种类C带差异表现在端带、居间带的有无,个别染色体上端带的大小、着色强弱,结构异染色质含量多少等方面。
从遗传学角度来看,C带所显示的结构异染色质区域在遗传上转录活性低,在种内水平上具有一定稳定性,低级阶元分类上是一个有用的指标[11,16]。但与银染带型相比,C 带的进化速率较快,往往在相近种间C 带异染色质的结构及组成都具有差异[13]。就异染色质含量来看,进行C带研究的蝗虫异染色质含量普遍在20%左右[8,11,13-17]。而姚世鸿[13]研究的同科的疣蝗(Trilophidiaannulata)异染色质含量为21.15%,韦仕珍和邓维安[17]研究的隆叉小车蝗(Oedaleusabruptus)和红胫小车蝗(O.manjius)异染色质含量分别为22.36%和22.07%,相比之下,东亚飞蝗的含量较低,仅为15.88%,与傅鹏和郑哲民[8]研究的癞蝗科友谊华癞蝗(Sinotmethisamicus)含量(15.18%)较为接近。说明异染色质含量在同科不同种间含量存在较大差异。一般情况下,C带处理的结果往往只能显示一个物种或种群结构异染色质主要的分布和含量特征[18]。而贾潇凌和马恩波[9]的东亚飞蝗C带结果显示其异染色质含量为18.87%,未见端带。根据Max和Bernard[19]报道,同物种的C带带型会随着如酸碱处理时间不同和染色体伸展程度等因素的影响而变化。因此,只有在同一细胞分裂时期并在基本相同的分带处理条件下才可能得出较为一致的结果。
3.2银染核仁组织区 银染是中期染色体的核仁组织区(NOR),Howell[20]和Hsu等[21]的研究显示,深染区域是具有转录活性的18S+28S rDNA。因此理论上深染点应该为偶数,但是本试验却观察到深染点大多呈奇数,且仅在同源染色体的其中1条上出现,陈晓光和何麟[22]在研究褐斑大蠊(Periplanetabrunnea)时也出现此现象。可推测东亚飞蝗只有一条18S+28S rDNA基因功能上有活性。间期及早前期深染点多,到减数分裂Ⅰ中期,只可见1处或2处深染点,可见,rDNA转录活性降低。减数分裂Ⅰ后期、末期以及减数分裂Ⅱ未见深染。2处深染的情况并不多见,另一处出现在1号染色体近着丝粒端的深染可能仅是2号染色体rDNA未充分表达时,作为补偿性的开放。此种核仁组织区的多态性,在以往研究的多种动物中也有发现,如灵长类[23]、两栖类[24]、鱼类[25]等。18S+28S rDNA转录酸性蛋白,而酸性蛋白又与染料中银离子特异性结合而着色[26]。
与C带、G带所不同的是,核仁组织区的定位以及其活性,具有较慢的进化速率和相对较大的保守性,一般较为恒定。正是由于核仁组织区定位在遗传学上的这种特点,使得该项技术成为揭示一个大分类单元(高级阶元)中种系发生关系的一个有用指标[27]。 本研究的结果显示,东亚飞蝗的核仁组织区定位于2号染色体端部次缢痕处(图2),经反复试验,发现其具有恒定特点。而贾潇凌和马恩波[9]研究的东亚飞蝗银染定位在M8上,产生差异的原因尚不明白,需做进一步研究。
马恩波和郭亚平[14]在研究斑腿蝗科发现银染核仁组织区与C带有相关性,出现Ag深染区域也有C带带纹,而在稻蝗属、卵翅蝗属、伪稻蝗属[28]中也发现此对应关系。但是本试验的结果却未显示出这种关联,东亚飞蝗的核仁组织区位于2号染色体靠近端粒区域,C带在次缢痕区域未有带型,仅出现端粒深染的情况。在有关飞蝗属的文献中也未发现有此种联系,这种联系并不存在于飞蝗属中。
东亚飞蝗核仁组织区定位于2号染色体,且2号染色体的C带带型丰富,因此可作为东亚飞蝗的遗传标记。
[1] 孙涛,赵景学,田莉华.草地蝗虫发生原因及可持续管理对策[J].草业学报,2010,19(3):220-227.
[2] 王俊梅.生物技术对草原蝗虫的控制效果及应用前景[J].草业科学,2009,26(9):206-211.
[3] 王俊梅,豆卫,谭成虎.绿缰菌复合油剂对草原蝗虫的防治试验[J].草业科学,2009,26(10):155-159.
[4] 陶红梅,蔡志华,何正波,等.东亚飞蝗染色体的减数分裂观察[J].重庆师范大学学报(自然科学版),2010,27(2):20-22.
[5] 赵建,何孟元,郝水.红翅皱膝蝗减数分裂染色体的螺旋与轴结构[J].实验生物学报,1990,23(3):261-271.
[6] Summer A T.A simple technique for demonstrating centromeric heterochromatin[J].Experimental Cell Research,1972,75:304-306.
[7] Howell W M,Black D A.Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method[J].Experientia,1980,36:1014-1015.
[8] 傅鹏,郑哲民.两种华癞蝗的染色体C-带核型[J].遗传,1989,11(3):26-28.
[9] 贾潇凌,马恩波.斑翅蝗科部分种属细胞分类学的研究[J].山西大学学报(自然科学版),1998,21(3):291-296.
[10] 马恩波,欧晓红.蝗总科染色体研究及科级综合比较(直翅目)[J].昆虫分类学报,2002,22(1):6-10.
[11] 李新江,张道川,王文强.蝗虫染色体C-带核型研究进展(昆虫纲:直翅目)[J].河北大学学报(自然科学版),2005,24(3):332-336.
[12] Webb G C.Cytogenetics of the parthenogenetic grosshopper warrmba (formerly noraba) virgo and its bisexual relatives.Ⅳ Chromosome banding studies[J].Chromosoma,1980,67:309-339.
[13] 姚世鸿.贵州蝗总科5种蝗虫的核型和C-带[J].贵州师范大学学报,2006,24(2):18-22.
[14] 马恩波,郭亚平.四种斑腿蝗科昆虫染色体带型的比较[J].昆虫学报,2001,44(3):268-275.
[15] 马恩波,郑哲民.五种稻蝗染色体核型和C带带型的比较[J].昆虫学报,1989,32(4):399-404.
[16] 霍满鹏,王文强,蒲力群,等.菊蝗和黄佛蝗三个种染色体C带带型分析[J].陕西师范大学学报(自然科学版),1997,25:48-52.
[17] 韦仕珍,邓维安.两种小车蝗染色体C带核型研究(蝗总科:斑翅蝗科)[J].四川动物,2009,28(1):7-10.
[18] Sants J L,Giraldz R.C-heterochromatin polymorphism and variation in chiasma localization inEuchorthippuspulvinatusgallicus(Acrididae,Orthoptera)[J].Chromosoma,1982,85:507-508.
[19] Max K,Bernard J.Regularities and restrictions governing C-band variation in acridoid grasshoppers[J].Chromosoma,1980,76:123-150.
[20] Howell W M.Chromosome core structure revealed by sliver staining[J].Chromosoma,1979,73:61-66.
[21] Hsu T C,Spirito S E, Pardue M L.The distribution of 18+28S ribosomal gene in mammalian Chromosomes[J].Chromosoma,1975,53(1):25-36.
[22] 陈晓光,何麟.褐斑大蠊的银染核仁形成区及染色体骨架研究[J].第一军医大学学报,1988,8(4):343-339.
[23] 曹莜梅,陈宜峰,罗丽华.十一种灵长类染色体核仁组织者(NORs)的比较研究[J].动物学研究,1983,2(2):119-123.
[24] 尚克冈,李士鹏,金昊.花背蟾蜍的核型分析Ⅱ:银染核仁组成区在数目上的多态及在剂量上的补偿效应[J].遗传学报,1984,11(1):52-60.
[25] 王蕊芳,施立明,贺维顾.几种鲤鱼染色体核仁组织者的银染观察[J].动物学研究,1985,6(4):391-398.
[26] 马恩波,郭亚平,张冰.九种蝗虫核仁组成区定位及细胞分类学意义[J].山西大学学报(自然科学版),1993,16(2):217-222.
[27] 马恩波,郭亚平,郑哲民.斑腿蝗科10属NOR定位及其细胞分类学意义[J].动物学研究,1992,14(3):271-276.
[28] 马恩波,郭亚平.伪稻蝗及其近缘属的细胞学分类[J].昆虫学报,1995,38(1):54-60.
猜你喜欢
生物技术进展(2022年1期)2022-02-11
民间故事选刊·上(2022年1期)2022-02-09
中国畜牧杂志(2020年10期)2020-10-19
小小说月刊·下半月(2020年9期)2020-10-09
科学之谜(2018年9期)2018-12-17
华夏地理(2017年5期)2017-08-16
新农村(2016年10期)2016-03-06
遗传(2015年4期)2015-02-04
Zoological Research(2012年6期)2012-08-15
台湾农业探索(2012年6期)2012-02-15
