
5种冷季型草坪草的耐热性研究
2011-04-25 09:44张芸芸
草业科学 2011年11期
孙 彦,张芸芸
(中国农业大学动物科技学院草业科学系,北京 100193)
草坪业作为一门新兴行业在我国迅速发展,而冷季型草坪草因其绿期长、质地柔软、色泽嫩绿、外貌美观等优点,更是深受广大人们的喜爱[1]。随着温室效应造成全球气候变暖,全球平均气温升高[2-3]。7―9月为北京的高温高湿期,平均温度高于30 ℃的高温天气可持续数周之久,有时极端温度超过40 ℃,或者在极强光照下,草坪草叶表温度可超过50 ℃,平均湿度也可达76%,给草坪草生长造成极大的甚至是致死的伤害。因此,冷季型草坪草的耐热性研究就显得非常重要。
高羊茅(Festucaarundinacea)、多年生黑麦草 (Loliumperenne)、草地早熟禾(Poapratensis)、匍匐剪股颖(Agrostisstolonifera)和粗茎早熟禾(P.trivialis)均属于禾本科冷季型草坪草,被广泛应用于普通绿化、高尔夫球场、足球场、保龄球场等草坪建植[1]。
针对高羊茅、草地早熟禾、匍匐剪股颖、多年生黑麦草各个草种的生理指标研究有报道[4-7]。但综合各个指标比较研究还鲜见报道。石永红等[8]对多年生黑麦草的高温半致死温度及耐热进行了研究。赵海明等[9]在夏季高温时对不同草地早熟禾品种坪用质量的影响进行了研究。王婷婷等[10]评价了利用细胞工程技术选育的草地早熟禾新种质的耐热性。对于5种草坪草同时比较耐热性在国内未见报道。
本研究通过实验室培养箱模拟北京地区的高温高湿环境,对这5种冷季型草坪草在热胁迫过程中的外观指标和生理生化指标进行分析,比较5种冷季型草坪草之间耐热性能的差异,从而筛选出耐热性能好的草坪草种,为北京地区建植耐高温冷季型草坪和降低草坪养护管理费用具有重要的理论与实际意义。
1 材料与方法
1.1试验材料 试验用草坪草为高羊茅的普通品种(F.arundinaceacv.Popular)、草地早熟禾的肯塔基品种(P.pratensiscv.K.B.G)、匍匐剪股颖的派尼A-4(A.stoloniferacv.PennA-4)、多年生黑麦草蒙特利品种(L.perennecv.Monterey Ⅱ),种子来自中国农业大学牧草种子实验室,粗茎早熟禾威罗品种(P.trivialiscv.Bariviera)种子由北京绿冠种业公司提供。
1.2试验准备 试验于2007年7月在中国农业大学院内进行。先将泥土、草炭、蛭石以1∶1∶1的比例混合,加入适当的沙子调节土壤透性,装入45盆直径为13 cm的塑料花盆中待用。将5种草坪草种子均匀撒播至盆中(每种草种植9盆),再覆一层细薄土,轻轻压实。出苗前经常用喷雾器浇水,保持土壤的湿润。出苗后按照常规方法管理,每隔2 d浇一次水,以浇透为原则;每周进行一次修剪,修剪高度高羊茅、草地早熟禾、多年生黑麦草和粗茎早熟禾为6 cm,匍匐剪股颖为3 cm。待草坪草生长60 d后转移到可控温湿培养箱(日本三洋)中生长。
1.3培养箱设置 将45盆草坪草随机分成2组,对照组15盆(每种草坪草3盆),胁迫组30盆(每种草坪草6盆,每2盆作为1个处理),随机放入两台温度设置为25 ℃/20 ℃(昼/夜),湿度75%,光周期12 h,光强3 000 lx的培养箱中进行培养。待草坪草生长1周后,调整胁迫组培养箱的参数,使其温度变为35 ℃/30 ℃(昼/夜),湿度75%,光周期12 h,光强3 000 lx,作为高温胁迫处理;对照组培养箱的各个参数不变,作为对照处理。处理期间各盆草坪草按照常规方法进行管理,保证土壤干湿交替。分别在草坪草生长的第0、5、10、15和20天取样,放入-70 ℃的冰箱中待用,然后进行形态指标和生理生化指标的测定。每个处理重复3次。
1.4测定项目及方法 各项测定指标均以鲜质量为基础进行计算。
1.4.1叶片含水量的测定 剪取的草坪草新鲜叶片(FW),放入110 ℃的烘箱中快速杀死植物组织以防止呼吸作用的消耗,再将其放入70 ℃的烘箱中烘24 h,冷却至室温后进行称干质量(DW)[11]。
叶片含水量=(FW-DW)/FW×100%。
1.4.2叶片相对电导率的测定 剪取新鲜叶片0.1 g,将其洗净后浸在20 mL去离子水中,常温下震荡24 h后用电导仪测其溶液的电导率Cinitial,然后再放入沸水中处理20 min以杀死组织,冷却至室温后再测定溶液的电导率Cmax[11]。
相对电导率=Cinitial/Cmax×100%。
1.4.3叶绿素含量的测定 叶绿素含量的测定采用95%丙酮-乙醇提取液的方法[11],准确称取0.5 g(精确到0.1 mg)植株叶片,加6 mL双蒸水和少量CaCO3,匀浆,定容至25 mL,过滤后取2.25 mL滤液放入50 mL比色管中,加入23.75 mL 95%丙酮-乙醇(丙酮∶乙醇=2∶1)溶液匀浆暗中提取叶绿素20~30 min,将提取液分别在663 nm和645 nm下进行比色,用Arono公式计算叶绿素的含量。
叶绿素a (Chla)(mg/L)=12.7A663-2.69A645
叶绿素b(Chlb)(mg/L)=22.9A645-4.68A663
叶绿素总量(Chl)(mg/L)=Chla+Chlb
1.4.4抗氧化物酶的测定 称取0.5 g叶片,加入5 mL预冷提取液(含50 mmol/L pH值7.0的磷酸缓冲液和1%PVP),在冰浴中研磨成匀浆,匀浆经10 853 r/min转速下离心30 min,上清液即为粗酶液,保存于4 ℃条件下备用,用于测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性[11]。
SOD的测定:取样品50 μL酶液,加入3.9 mL反应混合液含50 mmol/L pH值7.8的磷酸缓冲液、77.12 μmol/L硝基四唑蓝(NBT)、0.1 mmol/L乙二胺四乙酸(EDTA)、13.37 mmol/L蛋氨酸和0.1 mL 80.2 μmol/L核黄素,以不加酶液作为对照,放在3 000 lx下光照10 min后,于560 nm处测定SOD值。以每分钟抑制NBT光氧化还原50%的酶用量为1个酶活单位[U/(min·g)]。
CAT的测定:取0.1 mL酶液,加入3 mL反应液(100 mL 0.05 mol/L pH值7.0的磷酸缓冲液+85 μL 30%H2O2)后,以3.1 mL 0.05 mol/L pH值7.0的磷酸缓冲液作为对照,立即于240 nm下比色,酶活性以1 min中内H2O2在A240下降的吸光度值为一个酶活单位[U/(min·g)]。
POD的测定:取20 μL酶液,加入1 mL 0.1 mol/L pH值6.0的磷酸缓冲液和3 mL POD反应液(50 mL 0.1 mol/L pH值6.0的磷酸缓冲液+20 μL愈创木酚+19 μL 30%H2O2)后,以4 mL 0.1 mol/L pH值6.0的磷酸缓冲液为对照,立即于470 nm下比色,酶活性以ΔOD470nm/(min·g)表示。
1.4.5每株绿叶数的测定 在每盆草坪草中随机选取10株植株,分别测定每株的绿叶数目,即为绿叶数/株。
1.4.6植株生长高度的测定 在每盆草坪草中随机选取10株植株,测定植株基部到叶片最高点之间的距离,作记录。处理5 d后,再在每盆草坪草中随机选取10株植株,测定植株基部到叶片最高点之间的距离,两者之差即为草坪草在这5 d的植株生长高度。
1.5统计方法 对试验条件下测定的指标采用one-way ANOVA进行方差分析,对5种草坪草各个指标含量的聚类分析采用的是系统聚类分析(hierarchical cluster)方法,统计分析中所用的分析软件为SPSS 11.5。
2 结果
2.15种草坪草不同高温胁迫下叶片相对含水量的变化 在不同的高温胁迫时间下,5种冷季型草坪草的叶片相对含水量的变化之间差异显著(P<0.05)。高羊茅在高温胁迫的第5天比第0天相对含水量显著降低,第10天与第5天之间差异不显著(P>0.05),第10、15和20天三者之间下降的相对含水量均差异显著。多年生黑麦草在高温胁迫的第5天其叶片相对含水量下降的幅度较第0天差异不显著,其余各个处理间差异均显著。而匍匐剪股颖,草地早熟禾和粗茎早熟禾这3种草坪草的5个处理间差异显著(表1)。
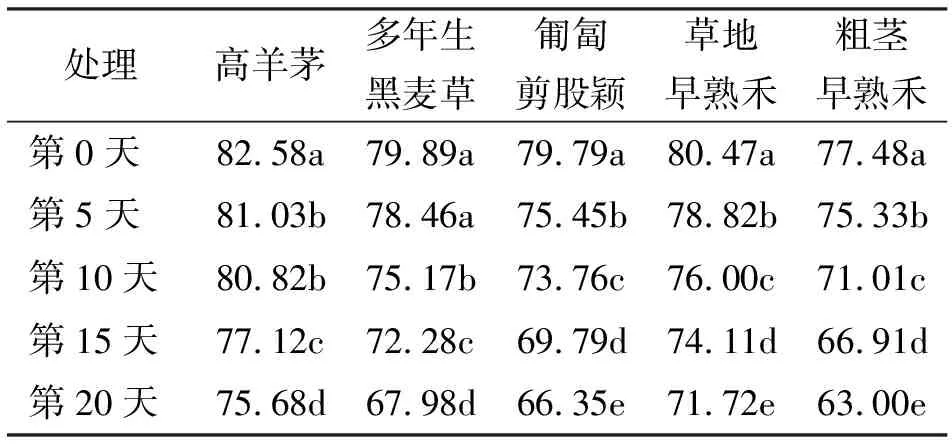
表1 5种草在高温胁迫下叶片相对含水量的变化 %
2.25种草坪草高温胁迫下叶片相对电导率的变化 在不同的高温胁迫下,5种冷季型草坪草的叶片相对电导率之间存在显著差异,多年生黑麦草和草地早熟禾各个处理间都是第15和20天之间相对电导率的增加差异不显著(P>0.05),其余各个处理间存在着显著差异(P<0.05)。而高羊茅、匍匐剪股颖和粗茎早熟禾3种草在5个不同处理时间的相对电导率均显著增加(表2)。
2.35种草坪草在高温胁迫下叶片叶绿素含量的变化 高羊茅在高温胁迫的当天与第5和10天之间叶片叶绿素含量差异不显著(P>0.05),而第10天较第5和15天叶绿素下降显著(P<0.05),第20天与第15天之间差异不显著。多年生黑麦草和匍匐剪股颖之间差异显著情况一致,均是在高温处理的当天和第5天之间差异不显著,第10天比第5天叶绿素含量下降显著,第15天与第10天之间差异不显著,第20天与第15天之间差异显著。草地早熟禾5个处理间叶绿素含量下降显著。粗茎早熟禾在高温处理的第5天与第0天之间差异不显著,其余各个处理间的差异均显著(表3)。
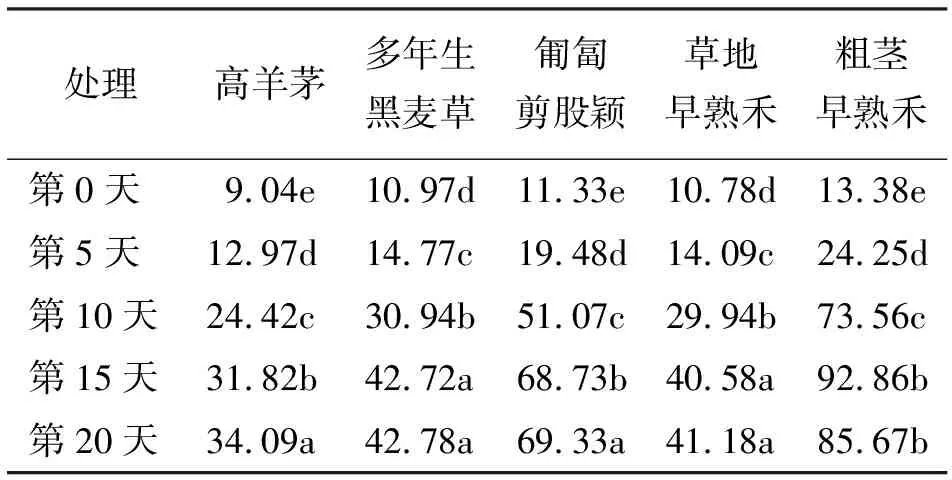
表2 5种草坪草在高温胁迫下叶片相对电导率的变化 %
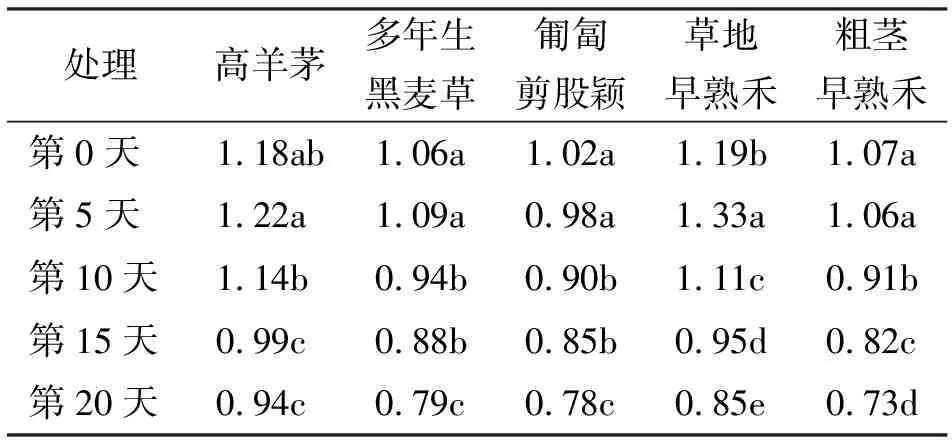
表3 5种草坪草在高温胁迫下叶片叶绿素含量的变化 mg/g
2.45种草坪草不同高温胁迫下叶片抗氧化酶活性的变化 高羊茅、草地早熟禾与粗茎早熟禾的各处理之间差异显著情况表现一致,均是第5天比第0天SOD活性显著增加(P<0.05),第10天与第0天相比下降不显著(P>0.05),第15天和第20天之间下降显著,且与其他各个处理间的也表现出差异显著。多年生黑麦草和匍匐剪股颖2种草坪草的5个处理间差异均显著(表4)。
在高温胁迫的5种不同时间处理下,5种冷季型草坪草的CAT活性均表现为随着胁迫时间的延长显著下降(P<0.05)(表5)。
高羊茅在高温胁迫的第5天比第0天的POD活性值显著增加,第10天与第5天的活性值差异不显著,第15天与第5和10天之间差异显著,但却与第0天之间差异不显著,在高温胁迫的第20天,其POD活性下降,与其他各处理间差异均显著。草地早熟禾在高温胁迫的第5和10天,POD活性值均较第0天表现出显著增加,而第15天与第0天之间差异不显著,第20天显著低于其余各处理。多年生黑麦草、匍匐剪股颖和粗茎早熟禾3种草坪草之间的显著性表现一致,均是高温胁迫的第5天比第0天POD活性增加,且差异显著,第10天与第0天之间差异不显著,但与第5天之间差异显著,第15和20天2个处理间差异显著,且与其他各个处理间的差异也显著(表6)。

表4 5种草坪草在高温胁迫下叶片SOD活性的变化 U/(g·min)
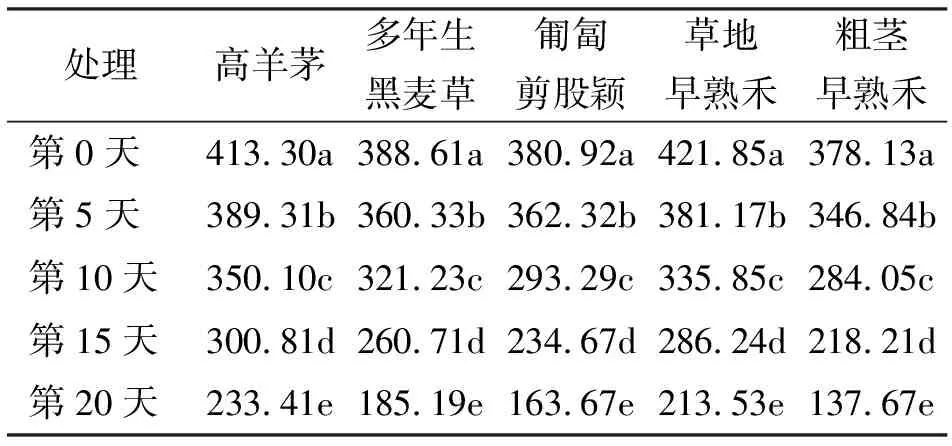
表5 5种草坪草在高温胁迫下叶片CAT活性的变化 U/(g·min)

表6 5种草坪草在高温胁迫下叶片POD活性的变化 OD470 nm/(g·min)
2.55种草坪草不同高温胁迫下每株绿叶数的变化 高羊茅在高温胁迫条件下,其每株绿叶数减少趋势较为平缓,在第0、5、10和15天4个不同高温时间的处理下,每株绿叶数之间差异不显著,在第20天其每株绿叶数的减少才表现出显著差异。草地早熟禾第5天与第0天之间,每株绿叶数下降,差异不显著,但在高温胁迫的第10、15天和第20天之间差异显著。多年生黑麦草、匍匐剪股颖和粗茎早熟禾3种草坪草的5个不同高温胁迫处理间均表现出显著降低(表7)。
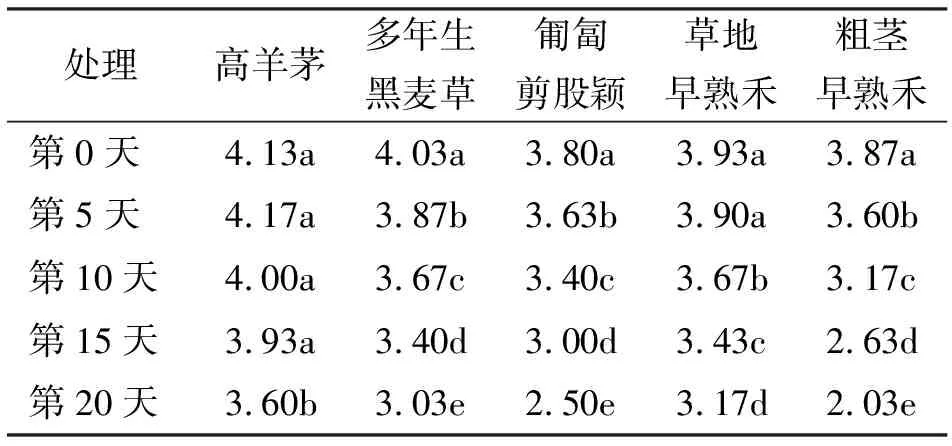
表7 5种草坪草在高温胁迫下绿叶数/株变化 数/株
2.65种草坪草不同高温胁迫下植株生长高度的变化 高羊茅、多年生黑麦草和匍匐剪股颖在不同高温胁迫时间下差异显著性表现一致,第5天与第0天植株的生长高度差异不显著,而在高温胁迫的第10、15和20天之间显著递减。草地早熟禾与粗茎早熟禾在高温胁迫的5种不同时间处理下均表现出显著递减趋势(表8)。
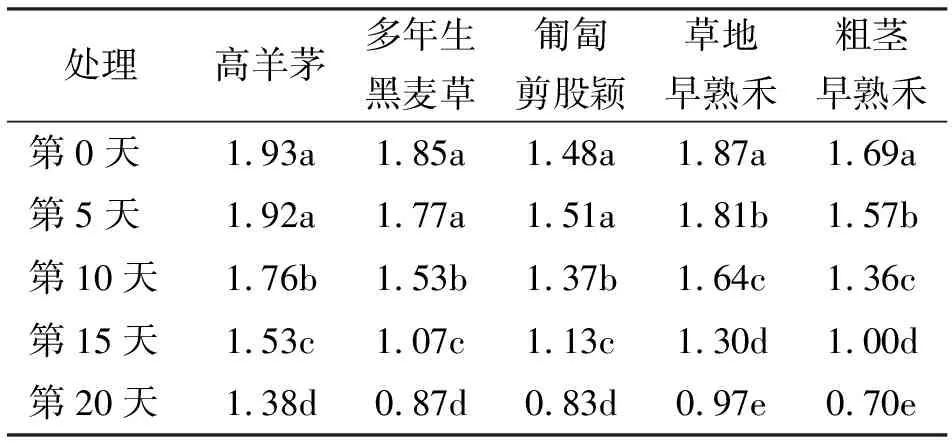
表8 5种草坪草在高温胁迫下植株生长高度变化 cm
2.7耐热性综合评价 综合上述生理生化指标和外观质量指标在处理的不同时间点的增加率和降低率,将5种草坪草进行聚类分析,结果表明(图1)5种草坪草可以大致聚为3类,其中高羊茅单独聚为一类,其各项指标总体变化稳定,耐热性最好;多年生黑麦草、草地早熟禾、匍匐剪股颖3种草坪草聚为一类,其各项指标总体变化相对稳定,耐热性相对较强;粗茎早熟禾单独聚为一类,其各项指标总体变化不稳定,耐热性较差。

图1 5种草坪草各个指标综合聚类分析
3 讨论与结论
3.1高温下叶片相对含水量的变化 在昼夜35 ℃/30 ℃的高温胁迫下,5种草坪草叶片相对含水量下降。草坪草叶片相对含水量的平均值从高到低为高羊茅>草地早熟禾>多年生黑麦草>匍匐剪股颖>粗茎早熟禾。高羊茅的叶片相对含水量显著高于其他4种草坪草。逆境中植物叶片相对含水量的大小,可以部分反映植物抗逆性的能力[12]。根据本试验得出的结果可以认为当供试草坪草处于昼夜35 ℃/30 ℃高温环境中,其体内的相对含水量均显著下降,下降的幅度随着处理时间的延长而逐渐增大。这说明处理时间越长,蒸腾强度越大,草坪草体内水分亏缺加剧。
3.2高温下叶片相对电导率的变化 当植物受到恶劣的环境胁迫时,其质膜透性增大已被许多研究证实[13-15]。5种草坪草叶片相对电导率上升,其平均值从低到高为高羊茅<草地早熟禾<多年生黑麦草<匍匐剪股颖<粗茎早熟禾。粗茎早熟禾的相对电导率显著高于其余4种草坪草。试验表明,高温胁迫下叶片的质膜透性呈持续增长趋势,且在某两个特定时间出现两个转折点,首先在第0天到第5天之间质膜透性缓慢增长,之后在第5天到第15天突然急剧上升,最后在第15天出现平缓上升趋势,当高温胁迫时间增加到一定程度时,可能引起膜蛋白的凝聚和变性,膜结构失去活性,膜对细胞内外物质交换和控制能力逐渐丧失,使外渗液急剧增加;时间继续延长,细胞膜功能完全丧失,外渗液电导率趋近死亡电导率,细胞开始大量死亡。
3.3高温下叶片叶绿素含量的变化 5种冷季型草坪草叶片叶绿素含量在高温环境中变化趋势不一致,高羊茅、多年生黑麦草、草地早熟禾的叶绿素含量呈先上升后下降的趋势,而匍匐剪股颖和粗茎早熟禾则呈持续下降的趋势。5种草坪草的叶绿素平均值的高低为高羊茅>草地早熟禾>多年生黑麦草>匍匐剪股颖>粗茎早熟禾。
3.4高温下酶的变化 在适应高温逆境的过程中,草坪植物体内也形成了一套清除这些自由基或活性氧的保护酶体系,即酶促防御体系[16-17],其中包括超氧化物歧化酶、过氧化氢酶、过氧化物酶等都是植物体内自由基的清除剂,三者协调一致使生物自由基维持在正常水平从而防止自由基伤害。在高温逆境下,植物体内的过氧化物酶和超氧化物歧化酶能阻止膜过氧化产物丙二醛含量增加,使自由基产生与清除平衡,增强植物抗热性[18]。本试验5种草坪草的超氧化物歧化酶、过氧化物酶活性呈现先上升后下降的变化趋势。过氧化氢酶活性下降,且随着时间的延长,过氧化物酶和过氧化氢酶活性下降的幅度增大。
3.5高温下绿叶数的变化 在高温胁迫过程中,5种草坪草每株绿叶数降低。高羊茅的每株绿叶数显著高于其他4种草坪草。
3.6高温下植株高度变化 随着高温胁迫时间的延长,5种草坪草植株生长高度降低。高羊茅与草地早熟禾之间的植株生长高度差异不显著,但却显著高于其余3种草坪草;草地早熟禾与多年生黑麦草之间差异不显著,但显著高于匍匐剪股颖和粗茎早熟禾;多年生黑麦草、粗茎早熟禾与匍匐剪股颖三者间植株生长高度差异均不显著。
3.7综合分析 通过聚类分析可将5种草坪草大致聚为3个耐热级别:1级(相对耐热)高羊茅;2级(中等耐热)草地早熟禾、多年生黑麦草、匍匐剪股颖;3级(相对敏感)粗茎早熟禾。
[1]孙彦.草坪实用技术手册[M].北京:化学工业出版社,2001:34-59.
[2]陈育峰.全球变化的基本特征及其与大气CO2浓度增加的关系研究[J].大自然探索,1998,17(64):32-35.
[3]Houghton J T,Meiro Filho L G,Callander B A.Climate change 1995:The Science of Climate Change[M].Cambridge,UK:Cambridge University Press,1996:55-100.
[4]赵昕,李玉霖.高温胁迫下冷地型草坪草几项生理指标的变化特征[J].草业学报,2001,10(4):85-91.
[5]Kenneth B M,Cell membrane thermostability and whole plant heat tolerance of Kentucky bluegrass[J].Crop Science,1998,38:1214-1218.
[6]Lyons E M,Pote J,DaCosta M,etal.Whole-plant carbon relations and root respiration associated with root tolerance to high soil temperature forAgrostisgrasses[J].Environmental and Experimental Botany,2007,59:307-313.
[7]Xu S,Li J,Zhang X,etal.Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation,antioxidant metabolites,and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress[J].Environmental and Experimental Botany,2006,56:274-285.
[8]石永红,刘建宁,王运琦,等.多年生黑麦草高温半致死温度与耐热性研究[J].草业科学,2010,27(2):104-108.
[9]赵海明,刘君,杨志民.夏季高温对不同草地早熟禾品种坪用质量的影响[J].草业科学,2010,27(1):4-10.
[10]王婷婷,徐国荣,张举仁,等.细胞工程技术选育的草地早熟禾新种质的耐热性分析[J].草业学报,2009,18(2):60-65.
[11]王忠.植物生理学[M].北京:中国农业出版社,2005:500-570.
[12]Chen C T,Chen L M,Lin C C,etal.Regulation of proline accumulation in detached rice leaves exposed to excess copper[J].Plant Science,2001,160:283-290.
[13]Liu X Z,Huang B R.Heat stress injury in relation to memebrane lipid peroxidation in creeping Bentgrass [J].Crop Science,2000,40(2):503-510.
[14]Steward C R,Hanson A D.Proline Accumulation as a Metabolic Response to Water Stress,Adaptatin of Plant to Water and High Temperature Stress[M].New York:Academic Press,1980:173 -189.
[15]王洪春.植物抗逆性与生物膜结构功能研究进展[J].植物生理学通讯,1985(1):60-64.
[16]余叔文.植物的热害和叶组织电阻的变化[J].植物学报,1984,26(4):397-404.
[17]Jiang Y,Huang B.Drought and heat stress injury to two cool-season turfgrasses in relation of antioxidant metabolism and lipid peroxidation[J].Crop Science,2001,41:436-442.
[18]王代军,温洋.温度胁迫下几种冷季型草坪草抗性机制的研究[J].草业学报,1998,7(1):75-80.
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14
中国果业信息(2021年10期)2021-12-07
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
养殖与饲料(2020年6期)2020-02-18
农业与技术(2020年2期)2020-02-05
小学生必读(低年级版)(2019年5期)2019-08-30
中国种业(2019年8期)2019-08-26
山西农经(2019年5期)2019-02-22
家教世界(2017年8期)2017-06-21
孩子(2016年4期)2016-04-13
