
大头稗(005)与鹅头稗(031)的生物学性状与细胞学研究
2011-03-27 06:56吴姝菊李新玲唐凤兰
草业学报 2011年3期
吴姝菊,李新玲*,唐凤兰
(1.哈尔滨师范大学生命科学与技术学院;黑龙江哈尔滨 150025;2.黑龙江省农业科学院 草业研究所;黑龙江哈尔滨 150086)
稗草(Echinochloa crusgalli)为禾本科(Gramineae)稗属(Echinochloa)植物,原产于欧洲和印度[1-2]。全属近30种,遍布于全世界的热带和温带地区,我国有9个种5个变种[3]。稗草含有大量的营养物质,草质柔嫩,适宜在湿盐碱地上种植,其鲜草、干草各种家畜均喜食,营养价值较高,粗蛋白质占干物质的9.42%,粗脂肪占2.45%,粗纤维占36.15%,无氮浸出物占33.88%,粗灰分占10.3%[4]。籽实可食用及酿酒,也可作畜禽的精料,根及幼苗可入药,全草可作绿肥。稗草叶多而且大,茎叶比小、适口性好,不仅适于饲喂牛、羊,而且适于饲喂鹅、兔。其生物学特性和植物学特征适合作为一种广泛种植的牧草,将成为我国主要的栽培牧草之一。过去人们常把稗草作为除莠对象,忽视了对稗草的研究和利用。此次对稗草开展研究,目的是要加强对寒地稗草资源的开发和利用。
黑龙江省各地目前种植着很多稗草,品种分类混乱,有同名异物和同物异名的现象存在,对这些稗草之间的亲缘关系尚不清楚,利用起来十分不便。稗草在黑龙江省分布种类目前还没有确切资料可供参考,只是有部分学者从栽培角度对稗草做了一些研究,如对稗草的主要防治措施,稗草DNA的提取,稗草对稻田的影响,如何识别稗草,稗草对各种不同除草剂的抗药性等方面的研究。稗草资源情况至今没有详细的报导,相关研究资料也比较少。本次对黑龙江省内收集的大头稗(005)和鹅头稗(031)2种寒地稗草进行的生物学性状测试和细胞学分析研究,在国内尚属首次[5]。试验研究的主要目的是了解2种稗草的生物学性状和核型基础,为更好地开展寒地稗草育种工作及生产上更好地栽培利用这2种稗草提供理论依据。
1 材料与方法
1.1 材料来源
大头稗(005)采自黑龙江省宁安县;鹅头稗(031)采自黑龙江省木兰县。
1.2 试验方法
1.2.1 生物学性状测试 试验地设在黑龙江省哈尔滨师范大学生命科学与技术学院试验田内(126°42′E,45°24′N)。2007年1-10月,黑龙江省年均降水量为402mm,哈尔滨及周边市县有效年积温为2 700℃左右。试验区土质为黑壤土,土壤肥力均匀,肥力水平较高。每个重复小区面积为3 m×5 m,试验设置3次重复,采用随机区组设计。
整地:在播种前1年秋翻土地,并施入基肥,有机肥用量为20 000 kg/hm2。播种前耙耱土地,整平碾碎,以保证土壤墒情。
播种方法:人工播种,单条播,行距为20cm,播种深度为3~4cm。播种量为20 kg/hm2。播种时间为2007年5月17日。
在出苗期、三叶期、分蘖期、拔节抽穗期等生长发育关键时期,对每个品种在每个小区随机调查20株,分别测量单株的株高、茎粗、叶长、叶宽、单株分蘖数、穗长、穗色、鲜干重等;收获时测定小区实际产量,对获得的各处理试验结果进行统计分析。小区收获时间为6月25日进行第1次刈割,留茬高度为15cm,第2次刈割时间为9月25日,2次产量之和为每个小区的实际收获产量。另外,收获时每小区留置1 m×1 m作为继续观察试验使用。统计分析利用JM T JFX简明统计分计CS 11.58来完成。
1.2.2 核型分析方法 采用常规的形态分析方法[6,7]。染色体制片过程为:在上午10:00,将根长度为0.5~1.0cm的稗草种子自培养皿中取出,放入小瓶内,加入对二氯苯饱和溶液,室温下处理4~5 h后[8,9],再放入卡诺固定液固定24 h,后用1 mol/L的HCl作解离液,在60~65℃的水浴锅中解离9 min左右取出根尖。用改良石碳酸品红染色液进行染色,染色时间为24 h左右[10,11]。把染色后的根尖制片进行镜检观察。
从大量的制片中,统计约50个完整的细胞中期分裂相,确定各个品种的染色体数目。核型的具体分析参照李懋学和陈瑞阳[6]的标准。
2 结果与分析
2.1 2种稗草的主要生物学性状
从形态特征上看,大头稗株高比鹅头稗矮49.90cm,茎型均为直立,主茎粗差异明显,花序形状和大小差别不明显,穗色不同。大头稗叶片数比鹅头稗少1片,鹅头稗分蘖数较多,单株分蘖数平均可达12个(表1)。
2种稗草的播期相同,大头稗分蘖期比鹅头稗晚2 d,大头稗在9月29日成熟,鹅头稗在9月16日成熟。在黑龙江省的气候条件下,2种稗草的枯黄期都在10月5日,生育天数均为145 d。从牧草鲜干产量上看,鹅头稗比大头稗鲜重高出2.78%;大头稗比鹅头稗干重高出2.55%(表2)。

表1 2种稗草的主要形态特征Table 1 Main morphological characteristics of the two barnyard grass variety

表2 2种稗草的主要生物学性状Table 2 Main biological characteristics of two barnyard grass variety
2.2 2种寒地稗草生物学产量差异显著性测验
把2种稗草分别在3个重复区内收获的鲜干草重量,按国际制单位折算成标准产量后,进行鲜干草产量的差异显著性测验,用t检验法(表3)[12,13]。统计结果表明,大头稗和鹅头稗鲜干重差异均不显著。

表3 2种稗草鲜干产量的差异显著性测验(平均值±SD)Table 3 Significant of differences test on fresh forage and hay yield of two barnyard grass variety t/hm2
2.3 2种寒地稗草的生长动态分析
2.3.1 株高 2种稗草从出苗期至三叶期株高增长较慢(图1),分蘖期、拔节期,直到初花期株高增长迅速,开花后至乳熟期稗草株高增速放缓,乳熟到完熟期稗草的株高基本处于停滞生长状态,完熟期后株高不再增加。鹅头稗在开花期株高增长速度比大头稗快,差异显著(P<0.05),此期的株高增速最终决定了2种稗草在株高上的显著差异(P<0.05)。
2.3.2 叶面积指数 2种稗草的叶面积指数在苗期增长缓慢(图2),分蘖期至初花期,叶面积指数呈迅速上升趋势,一直保持着快速的增长,开花期后,由于叶片数量不再增加,只是叶片个体增大,因而叶面积指数处于平稳中有升的状态;2种稗草的叶面积指数都在乳熟期时达到最高值,乳熟期后叶面积指数均不再增加。2种稗草三叶期至分蘖期叶面积指数差异显著(P<0.05),拔节期至乳熟期叶面积指数差异不显著(P>0.05),完熟期时,鹅头稗叶面指数略高于大头稗,但差异不显著(P>0.05)。
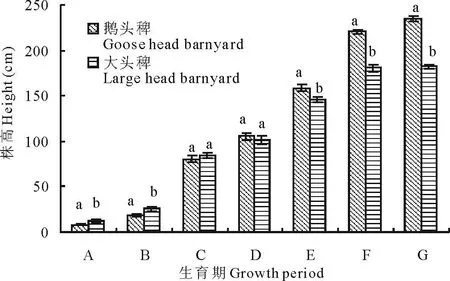
图1 2种稗草的株高生长动态Fig.1 Dynamics of plant height of two barnyard grass variety

图2 2种稗草的叶面积指数生长动态Fig.2 Dynamics of the leaf area index of twobarnyard grass variety
2.3.3 干物质积累 在生长季节内稗草全株干物质积累量均随生育时间延长而增加。2种稗草的鲜重在分蘖期时差异不显著(P>0.05)(图3);鹅头稗在拔节期至乳熟期鲜重增加速度比大头稗快,增速差异显著(P<0.05),这符合生育期间的观察结果。鹅头稗茎青叶嫩,多汁,含水量高,乳熟期后鲜重开始下降;而大头稗茎秆粗壮,生长势相对缓慢,鲜重增速没有鹅头稗快,完熟期时,鹅头稗鲜重略高于大头稗,但差异不显著(P>0.05)。
2种稗草的干重从分蘖期后随鲜重的增加快速增长(图4),初花期时,鹅头稗干重高于大头稗,差异达到显著(P<0.05);乳熟期后,2种稗草的干重增速放缓,从分蘖期至初花期,期间如加强肥水管理,不仅有利于根、茎和叶产量的增长,也同时能提高稗草籽实产量,对稗草整株产量的增加可起重要作用[14,15]。初花期后大头稗的干重增速比鹅头稗快,完熟期时大头稗干重高于鹅头稗,差异不显著(P>0.05)。

图3 2种稗草的单株鲜重增长动态Fig.3 Dynamics of fresh weight per plant of two barnyard grass variety
2.4 2种稗草的核型分析
大头稗共有27对染色体(表4),最长染色体相对长度为3.41,最短染色体相对长度为1.41,其中有23对为中着丝点染色体,4对为近中着丝点染色体。鹅头稗共有27对染色体(表5),最长染色体相对长度为2.41,最短染色体相对长度为1.09,其中有22对为中着丝点染色体,5对为近中着丝点染色体。在2种稗草的核染色体中均未发现近端和端着丝点染色体,也没有观察到有随体的染色体[16,17]。

表4 大头稗核型分析参数Table 4 Karyotype analysis parameters of Large head barnyard

表5 鹅头稗核型分析参数Table 5 Karyotype analysis parameters of goose head barnyard

图5 2种稗草核型Fig.5 Karyogram of twobarnyard grass variety

图6 2种稗草染色体配对结果Fig.6 Chromosome pairing map of two barnyard grass variety
核型分析结果(表4,5,图5,6)为:大头稗根尖细胞核内染色体数目2n=54。核型公式为2n=46m+8sm,无随体,染色体相对长度范围1.41~3.41,染色体长度比2.43,核型为2B型(图7);鹅头稗根尖细胞核内染色体数目 2n=54。核型公式为 2n=44m+10sm,无随体,染色体相对长度范围1.09~2.41,染色体长度比2.21,核型为2B型[18,19]。
3 讨论
试验测定的2种稗草均为采自黑龙江省内的寒地稗草,通过根尖细胞内染色体数目、染色体长度、染色体臂比、随体等具体指标的测试,初步确定了2种稗草的核型均为2B型。稗草在全国范围内几乎都有分布,对该物种的染色体数目记载有很大差异,已往所报导的稗草的染色体数目有2n=36,42,48,54等,数目不等[3],试验观察2种稗草的染色体数均为54条,与冯久焕和张廷壁[3]及Yabuno[5]所报导中的部分种类稗草的染色体数目一致。依据东北植物检索表中对所列出的稗草的种类和特征描述资料,可以推断,大头稗和鹅头稗不属于东北植物检索表中的稗草种类[1]。
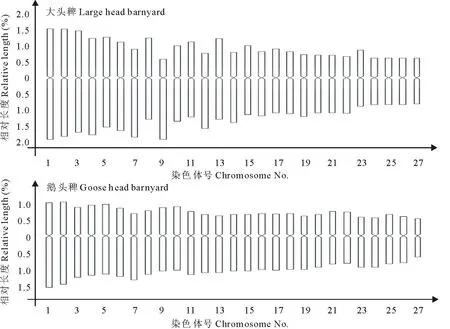
图7 2种稗草的核型模式图Fig.7 Karyotype model map of two barnyard grass variety
一般而言,核型数据如染色体的相对长度、平均臂比、随体的数量及位置等是鉴别种和亚种的重要依据。通过对2种稗草的核型研究,大头稗核型公式为2n=46m+8sm,鹅头稗核型公式为2n=44m+10sm,二者除染色体数目与冯久焕和张廷壁[3]所报导的“稗”一致外,在染色体形态特征和结构上与“稗”也有很多相似之处,可以初步推测,二者当同属于“稗”种,但二者的核型又与冯久焕和张廷壁[3]报导的“稗”有细微差别,可能是由于稗草在向栽培稗草演变过程中,发生了系列遗传性状上的改变[20,21]。核型分析数据支持2种稗草属于不同品种,不可能是同物异名种,应分属于不同栽培类型[22,23]。
综合测试结果表明,2种稗草的生物学性状均为良好。在黑龙江省的气候条件下,株型繁茂,枝叶发达,产量高,品质也较好,适宜作为省内的优质牧草进行广泛栽培。大头稗和鹅头稗都是粮草兼收的优质稗草,喜温、喜光,喜肥水,抗性好,茎秆粗壮,抗倒伏,耐寒性好,对土壤要求不严格。大头稗株高相对较矮,穗大粒多,成熟时大穗紫红色,全年生育期143 d,鹅头稗株高相对较高,叶多茎嫩,全年生育期135 d。
在试验条件下做产量差异测试,2种稗草在鲜干草产量上没有实质差异。根据生物学性状观测结果,对2种稗草的利用提出如下参考建议:鹅头稗茎叶鲜嫩,质地多汁,是很好的青饲牧草,而且该品种在分蘖期至乳熟期前植株长势好,增重快,刈割后再生长能力较强,更适宜做青饲牧草;大头稗茎秆粗壮,穗大粒多,干物质积累快,可做青干两饲牧草。
[1]傅沛云.东北植物检所表(第二版)[M].北京:科学出版社,1995:840.
[2]陈心启.中国植物志(第19卷)[M].北京:科学出版社,1999:311.
[3]冯久焕,张廷壁.国产稗属植物的细胞学研究[J].武汉植物学研究,1993,11(4):294-304.
[4]张月学,唐凤兰,严洪东.寒地牧草高产栽培关键技术(第二版)[M].哈尔滨:黑龙江科技出版社,2004:53-107.
[5]Yabuno T.Biosystomatic study of the genus Echinochloa[J].Japan Journals Botany,1996,19(2):277-323.
[6]李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
[7]李懋学,张赞平.植物染色体研究技术[M].哈尔滨:东北林业大学出版社,1991:1-30.
[8]杨艳萍,陈佩度.普通小麦与鹅观草属杂种F1及BC1的分子细胞学,育性和赤霉病抗性研究[J].遗传,2009,31(3):290-296.
[9]张宇澄,周颂东,任海燕,等.中国葱属10种20居群的核型研究[J].武汉植物学研究,2009,27(4):352-360.
[10]于海清,丁春邦,周永红.拟鹅观草属3种植物的核型研究[J].西北植物学报,2009,29(8):1590-1594.
[11]刘庆华,王奎玲,王玮,等.美丽胡枝子核型分析[J].西北植物学报,2009,29(8):1595-159.
[12]李祖发.引进的几个稗草品种在湖北的适应性及栽培利用[J].湖北农业科学,1990,(3):28-30.
[13]孙建华,王彦荣,余玲.紫花苜蓿生长特性及产量性状相关性研究[J].草业学报,2004,13(4):80-86.
[14]王斌文,曹致中,韩建国,等.苏丹草营养成份与农艺性状通径分析[J].草地学报,2005,13(3):203-208.
[15]刘建秀,郭海林,朱雪花,等.结缕草属种质资源的综合评价[J].草地学报,2005,13(3):219-222.
[16]宣继萍,郭爱桂,刘建秀,等.狗牙根辐射诱变后代外部性状变异分析[J].草业学报,2005,(6):107-112.
[17]罗乐,张启翔,白锦荣,等.16个中国传统月季品种的核型分析[J].北京林业大学学报,2009,(05):90-95.
[18]闵伶俐,宋娟娟,廖景平,等.濒危植物濒危植物兰花蕉的核型研究[J].广西植物,2009,29(5):595-598.
[19]蓝伟侦,张海洋,徐秀芳.少花龙葵与黄果龙葵染色体核型分析[J].广西植物,2009,29(5):599-602.
[20]牛西午,田如霰,李贵全,等.锦鸡儿属植物染色体制片与3个种的核型分析[J].西北植物学报,2006,26(5):1043-1047.
[21]张义贤.胡枝子的核型分析研究[J].山西大学学报(自然科学版),1990,13(1):87-89.
[22]刘建秀,郭爱桂,郭海林.我国狗牙根种子资源形态变异及形态类型划分[J].草业学报,2003,12(1):99-104.
[23]丁春邦,周永红,郑有良,等.拟鹅观草属6种2亚种和鹅观草属3种植物的核型研究[J].植物分类学报,2004,42(2):162-169.
猜你喜欢
中国农业科学(2022年8期)2022-05-16
内蒙古民族大学学报(自然科学版)(2021年4期)2021-09-02
中国种业(2021年2期)2021-03-01
草地学报(2019年3期)2019-07-25
中国奶牛(2019年1期)2019-02-15
吉林农业·下半月(2017年7期)2017-07-15
农民致富之友(2017年6期)2017-05-10
江苏农业科学(2014年8期)2014-10-23
