
环境因子对栉孔扇贝血淋巴中谷胱甘肽转硫酶活性的影响
2011-03-14 06:15:16王宜艳孙虎山周彦菊闫冬春
海洋科学 2011年10期
王宜艳,孙虎山,周彦菊,闫冬春,王 磊
(鲁东大学 生命科学学院,山东 烟台 264025)
谷胱甘肽转硫酶(Glutathione S-transferases,GST,EC2.5.1.18),也称为不含硒谷胱甘肽过氧化物酶(nonSeGPx),是一组多功能同功酶,其主要功能是催化谷胱甘肽(GSH)与各种内源性和外源性有害的亲电子化合物反应,生成无毒性或毒性小的GSH硫结合物; 同时能催化有机过氧化物还原成相应的醇,与硒谷胱甘肽过氧化物酶(SeGPx)协同作用起抗氧化作用[1]。软体动物的双壳类、腹足类和头足类的鳃、足、外套膜和消化腺等组织中都广泛存在着 GSH及其相关酶(SeGPx、GST和谷胱甘肽还原酶等),并在其抗氧化和解毒中起重要作用[2-5]。近10年来,国外有关贝类体内 GST的研究报道较多,多集中在重金属、杀虫剂和石油污染物等对GST等抗氧化因子和生物异源物质代谢系统的影响[2,6-13],对 GST等抗氧化因子的季节变化也有研究报道[14-16],研究的主要热点是探讨 GST等抗氧化因子在应激状态下的作用及作为环境污染的生物标志物的可行性; 国内有关贝类 GSH及其相关酶的研究报道极少[17-18]。对贝类体内GST性质的研究国内外均未见研究报道。作者以人工养殖的栉孔扇贝(C.farreri)为材料,采用生化测定的方法对其血淋巴中GST活性及部分性质进行了初步研究,以期为贝类防御机理的研究积累资料,也为贝类体内GST的功能研究提供理论依据。
1 材料与方法
1.1 材料和试剂
栉孔扇贝购自烟台水产市场,壳长 40~50 mm,暂养于室内充气的砂滤海水水族箱内。还原型谷肽胱甘肽、1-氯-2,4-二硝基苯(CDNB)等试剂均为国产分析纯产品。
1.2 血清和血细胞的制备
用5mL注射器和5号针头从栉孔扇贝闭壳肌血窦中抽取血淋巴,每次实验取 5只扇贝的血淋巴混合作为样品,4℃、3500 r/min离心10 min,取上清即血清用于血清中GST活力的测定。下层沉淀即血细胞中加入与血清等量的双蒸水低渗溶血后,4℃、6000 r/min离心10 min,取上清液即血细胞裂解液用于血细胞中GST活力的测定。样品制备后立即用于酶活测定,未用完的血样品置于4℃冰箱中保存不超过 12 h。
1.3 GST活性力测定
GST活力测定采用CDNB比色法[8],略有改进。对照管中依次加入0.1 mol/L、pH 6.5的磷酸缓冲液(PBS)0.1 mL、10 mmol/L的 GSH 0.1 mL及 1.25 mmol/L的CDNB 0.8 mL,测定管中以等量血样品取代PBS,25℃恒温下准确反应5 min; 然后在对照管中依次加入10%的盐酸0.1 mL、血样品0.1 mL、蒸馏水1.8 mL,测定管中用等量PBS取代血样品; 1 cm石英比色皿、340 nm下以对照管作空白,读取测定管的吸光度。测定管设5个平行组。采用Lowry等[10]的福林-酚试剂法测定样品中的蛋白质含量。酶活力单位的定义:每毫克蛋白每分钟催化生成1 μmol产物所需要的酶量为1个酶活力单位(U)。数据处理采用T检验。
1.4 GST性质的研究
1.4.1 温度及pH对血细胞中GST活力的影响
以0.1 mol/L、pH 6.5的PSB为缓冲液,分别在5、10、15、20、25、30、35、40、60℃的反应条件下测定酶活力,以测定温度对血细胞中 GST活力的影响。
以25℃为测定温度,分别以pH 5、5.5、6、7、7.5和8的0.1 mol/L的PBS为缓冲液,测定pH对血细胞中GST活力的影响。
1.4.2 血细胞中GST的热稳定性
将血细胞裂解液分别在 20、25、30、35℃恒温中放置30 min后; 在4℃中分别保存12、24 h后,以0.1 mol/L、pH 6.5的PBS为缓冲液,25℃测该酶的反应速率,以测定该酶的热稳定性。
1.4.3 金属离子铜、铅对血细胞中GST活力的影响
分别以硫酸铜和硝酸铅为抑制剂,以0.1 mol/L、pH 6.5的柠檬酸-柠檬酸钠为缓冲液,分别配制铜、铅离子终质量浓度为:0、0.05、0.25、0.5、2.5、5 mg/L的反应液,在25℃测得该酶的反应速率。
2 结果
2.1 血清和血细胞中GST活力
栉孔扇血清和血细胞中GST活力的测定结果见表1。由表1可见,栉孔扇贝血清和血细胞中均存在GST,血细胞中 GST活力远高于血清中的,是血清中的12倍。

表1 栉孔扇贝血清和血细胞中GST活力Tab 1 Activity of Glutathione S-transferases in the haemolymph of C.farreri
2.2 温度对血细胞中GST活力的影响
温度对血细胞中GST活力影响的测定结果见图1。结果表明:随温度的升高酶活力出现先升后降的趋势,该酶的最适温度范围为20~35℃。GST在5℃的低温下活力很低,仅为 48.14 U,10℃时活力有所升高,15℃时活力快速升高,为5℃时的3倍多; 30℃时酶活力达最高值,后随温度的继续升高而下降,35℃时活力较 30℃下降了 20%,40℃时其活力只有30℃时的23%,60℃时该酶已完全丧失活力。

图1 温度对栉孔扇贝血细胞中GST活力的影响Fig.1 Effects of temperature on GST activity in haemocytes of C.farreri
2.3 pH对血细胞中GST活力的影响
pH对血细胞中 GST活力影响的测定结果见图2。结果表明:pH为5时GST活力较低仅为62.31 U,pH为5.5和6时,酶活力有所升高,但变化不明显;pH为7.5时酶活力达最高值,为240.06 U。pH为7和8时酶活力分别达到最高值的61%和76%。该酶的适宜pH为7~8,最适pH为7.5。

图2 pH对栉孔扇贝血细胞中GST活力的影响Fig.2 Effects of pH on GST activity in haemocytes of C.farreri
2.4 血细胞中GST的热稳定性
4℃时血细胞中GST稳定性的测定结果见图3。结果表明:4℃时 GST活力随保存时间的延长而下降。4℃保存12 h后GST活力已下降了51%,4℃保存24 h后酶活力仅有原活力的32%。说明此酶在低温中保存也易失活,低温保存时间最好不要超过 12 h。
血细胞中GST热稳定性的测定结果见图 4。结果表明:血细胞裂解液在不同温度下保温30 min后,该酶随温度的升高活力逐渐降低,35℃时酶的活力显著降低,为20℃时的22%,为30℃的29%。因此,该酶热稳定性较差。

图4 栉孔扇贝血细胞中GST的热稳定性Fig.4 Heat stabilities of GST in haemocytes of C.farreri
2.5 铜和铅离子对血细胞中GST活力的影响
Cu2+对血细胞GST活力影响的测定结果见图5。结果表明:Cu2+对该酶活力有明显抑制作用,随着Cu2+质量浓度的升高抑制作用增强。Cu2+质量浓度为0.25 mg/L该酶活力较对照组已下降了近 40%; 当Cu2+质量浓度升高到0.5 mg/L时其活力较对照组下降了78%; 当Cu2+质量浓度升高到2.5 mg/L时酶活力已接近0。

图5 Cu2+对栉孔扇贝血细胞中GST活力的影响Fig.5 Effects of Cu2+ on GST activity in haemocytes of C.farreri
Pb离子对血细胞GST活力影响的测定结果见图6。结果表明:Pb2+对GST活力也有明显抑制作用,随着 Pb2+质量浓度的升高抑制作用增强。Pb2+为0.05mg/L时酶活力较对照组已下降了51%; Pb2+质量浓度升高到5mg/L时该酶失活。
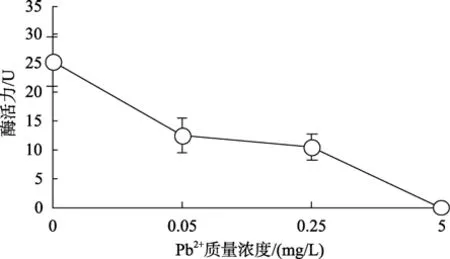
图6 Pb2+对栉孔扇贝血细胞中GST活力的影响Fig.6 Effects of Pb2+ on GST activity in haemocytes of C.farreri
3 讨论
本实验结果表明,栉孔扇贝血细胞 GST的最适温度为 30℃,较马素永等[19]CDNB法测得玉米螟(Ostrinia furnacalis)GST的最适温度41℃为低; 尹登科等[20]利用大肠杆菌表达日本血吸虫(Schistosoma japonicum)GST后,CDNB法测得其最适作用温度为37℃,也高于本实验结果; 日本血吸虫 GST对温度的稳定性也高于栉孔扇贝。玉米螟GST最适pH为6.3~6.9,日本血吸虫GST最适pH为7.0,均稍低于栉孔扇贝。GST是一组多功能同功酶,为由多种不同亚基构成的二聚体,不同物种不同组织中可有不同种类的GST[21],因此,在不同物种间可存在GST理化性质的差异。栉孔扇贝血细胞GST对温度的稳定性较差,室温下对该酶操作易失活,用于实验的样品在4℃保存时间最好不要超过12 h。栉孔扇贝不同组织中GST同功酶的分型、分布等有待于进一步的研究。本实验显示,应用CDNB法测定GST活力时,缓冲液种类有影响作用,同为pH6.5的缓冲液,25℃为测定温度的条件下,用PBS缓冲液较柠檬酸-柠檬酸钠缓冲液酶活力要高出近 8倍,其原因有待进一步研究。
贝类血细胞杀死被吞噬的病原菌等异物的主要机制之一是伴随吞噬引起的呼吸突发产生的具杀菌作用的超氧阴离子和 H2O2等活性氧,活性氧对贝类机体也有毒害作用,须及时清除。GSH可通过巯基与体内自由基结合转化成容易代谢的酸性物质,从而加速超氧阴离子、H2O2和脂质过氧自由基等自由基的排泄,同时GSH又可通过GST发挥作用,抑制脂质过氧化,保护细胞膜恢复细胞功能,并可保护细胞内含巯基的酶的活性,防止因巯基氧化导致蛋白质变性,减少自由基对 DNA的攻击,从而减少DNA损伤和突变[3]。本文结果表明扇贝的血淋巴中含有GST,说明扇贝体内也存在GST和GSH清除自由基的免疫防御机制。
贻贝(Mytella guyanensis)等滤食性双壳贝类组织中的 GSH、GST、SeGPx等对水中重金属离子等污染物比较敏感,污染物含量高的组织中 GST等的活力也较高[11,13],因此,某些贝类体内的 GST等可作为污染的生物标志物。但不同污染物对贝组织中GST活力的影响不同[6],已有报道显示不同质量浓度金属离子暴露实验对贝类GST活性影响无一致的规律,有的显示GST活性无改变[12],有的显示GST活性降低[10]。Hunaitia等[22]用不同质量浓度的氯化铅体外(37℃)孵育人全血24 h后,血液中GST活性随 Pb+质量浓度升高而下降,说明铅对 GST活力抑制作用的存在。本实验结果显示,金属离子铜和铅对栉孔扇贝血细胞GST活性有显著直接抑制作用。对贝类组织中GST性质尤其是抑制剂、激活剂的研究对GST作为污染的生物标志物的应用有一定的参考价值。
[1]陈瑗,周玫.自由基医学基础与病理生理[M].北京:人民卫生出版社,2002:126-132.
[2]Antognelli C,Francesca B,Andrea P,et al.Activity changes of glyoxalase system enzymes and glutathione-S transferase in the bivalve molluscScapharca inaequivalvisexposed to the organophosphate chlorpyrifos[J].Pesticide Biochem Physiol,2006,86:72-77.
[3]Bebianno M J,Company R,Serafim A,et al.Antioxidant systems and lipid peroxidation inBathymodiolus azoricusfrom Mid-Atlantic Ridge hydrothermal vent fields[J].Aquat Toxicol,2005,75:354-373.
[4]Pannunzio T M,Storey K B.Antioxidant defenses and lipid peroxidation during anoxia stress and aerobic recovery in the marine gastropodLittorina littorea[J].J Exp Mar Biol Ecol,1998,221:277-292.
[5]Zielinski S,Pörtner H O.Oxidative stress and antioxidative defense in cephalopods:a function of metabolic rate or age? [J].Comp Biochem Physiol,2000,125 B:147-160.
[6]Petushok N,Gabryelak T,Pałecz D,et al.Comparative study of the xenobiotic metabolising system in the digestive gland of the bivalve molluscs in different aquatic ecosystems and in aquaria experiments[J].Aquat Toxicol,2002,61:65-72.
[7]Lau P S,Wong H L.Effect of size,tissue parts and location on six biochemical markers in the green-lipped mussel,Perna viridis[J].Mar Pollut Bull,2003,46:1563-1572.
[8]Luca-Abbott S B D,Richardson B J,McClellan K E,et al.Field validation of antioxidant enzyme biomarkers in mussels (Perna viridis) and clams (Ruditapes philippinarum) transplanted in Hong Kong coastal waters[J].Mar Pollut Bull,2005,51:694-707.
[9]Looise B A S,Holwerdu D A,Foekemu E M.Induction of glutathione S transferase in the freshwater bivalveSghderium corneumas a biomarker for short-term toxicity tests? [J].Comp Biochem Physiol,1996,113C(1):103-107.
[10]Regoli F,Nigro M,Orlando E.Lysosomal and antioxidant responses to metals in the Antarctic scallopAdamussium colbecki[J].Aquat Toxicol,1998,40:375-392.
[11]Torres M A,Testa C P,Gáspari C,et al.Oxidative stress in the musselMytella guyanensisfrom polluted mangroves on Santa Catarina Island,Brazil[J].Mar Pollut Bull,2002,44:923-932.
[12]Almeida E A D,Miyamoto S,Bainy A C D,et al.Protective effect of phospholipid hydroperoxide glutathione peroxidase (PHGPx) against lipid peroxidation in musselsPerna pernaexposed to different metals[J].Mar Pollut Bull,2004,49:386-392.
[13]Cheung C C C,Zheng G J,Lam P K S, et al. Relationships between tissue concentrations of chlorinated hydrocarbons (polychlorinated biphenyls and chlorinated pesticides) and antioxidative responses of marine mussels,Perna viridis[J].Mar Pollut Bull,2002,45:181-191.
[14]Bocchetti R,Regoli F.Seasonal variability of oxidative biomarkers,lysosomal parameters,metallothioneins and peroxisomal enzymes in the Mediterranean musselMytilus galloprovincialisfrom Adriatic Sea[J].Chemosphere,2006,65:913-921.
[15]Borkovic S S,Saponjic J S,Pavlovic S Z.The activity of antioxidant defence enzymes in the musselMytilus galloprovincialisfrom the Adriatic Sea[J].Comp Biochem Physiol,2005,141C:366-374.
[16]Power A,Sheehan D.Seasonal variation in the antioxidant defence systems of gill and digestive gland of the blue mussel,Mytilus edulis[J].Comp Biochem Physiol,1996,114C(2):99-103.
[17]陈荣,郑微云,余群,等.石油污染对僧帽牡蛎谷光甘肽含量及相关酶活性的影响[J].海洋学报.2003,25(增 2):226-230.
[18]杨海灵,高波,曾庆银,等.对生蒴蛤谷光甘肽转移酶的光谱研究[J].高等学校化学学报,2003,24(8):1472-1476.
[19]马素永,周先碗,张廷芳.玉米螟谷胱甘肽转硫酶的纯化及性质研究[J].北京大学学报(自),1999,35(4):474-478.
[20]尹登科,丁虹,喻昕.壳聚糖固定化谷光甘肽硫转移酶的研究[J].生物技术,2004,14(6):17-19.
[21]聂立红,王声涌,胡毅玲.谷光甘肽硫-转移酶研究进展[J].中国病理生理杂志,2000,16(11):1240-1243.
[22]Hunaiti A A,Soud M.Effect of lead concentration on the level of glutathione,glutathione S-transferase,reductase and peroxidase in human blood[J].Sci Total Environ,2000,248:45-50.
猜你喜欢
海洋通报(2022年5期)2022-11-30 12:04:10
理化检验-化学分册(2020年5期)2020-06-15 11:36:14
家教世界·创新阅读(2019年11期)2019-12-10 09:22:04
食品与生活(2019年8期)2019-10-30 12:13:09
人生与伴侣·共同关注(2018年8期)2018-02-16 06:58:48
天然产物研究与开发(2016年6期)2016-06-05 10:29:26
分析测试学报(2015年6期)2016-01-13 06:19:00
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:36
作文周刊·小学一年级版(2014年31期)2014-04-29 00:44:03
食品科学(2013年22期)2013-03-11 18:29:30
