
雌雄异体生物种群的性比失调及其调控研究现状
2011-02-26 02:17:04费世民何亚平王乐辉蔡小虎陈秀明
四川林业科技 2011年2期
费世民,何亚平,何 飞,王乐辉,蔡小虎,陈秀明
(四川省林业科学研究院,四川成都 6l008l)
雌雄异体(dioecism;gonochorism;bisexualism),雌性和雄性的性腺(器官)分别生在不同个体上的生物,动物中脊椎动物基本上都是雌雄异体,植物中仅有6%种类是雌雄异体,植物中比例极低。雌雄异体生物种群有一个基本特征,那就是雌雄性存在一个明显的个体比例,这个个体比例称之为性比(Sex ratio)。性比是一种生物某一层次中雄性单位数和雌性单位数之比,种群性比主要集中在雌雄异体类生物。依据雌雄个体发育的时期,雌雄异体生物种群的性比不同,如雌雄异株植物性比分为受精期的性比(第一性比)、种子萌发期比(或相当的时期)的性比(第二性比)、幼体以后的时期(第三性比)(林益民,l993)。动物中性比也存在这三类,卵期性比、幼仔性比与成体性比。显然,雌雄异体生物种群性比涉及到两个问题,一个问题是受精时性别分化的问题,一个是雌雄个体发育存活率的问题。
自从达尔文时代,多数有机体几乎相等的雄性与雌性生产令生物学家着迷。l929年,Fisher指出,由于每一个来自有性生殖的个体都只有一个父亲和一个母亲,总体上父本与母本生殖值相等。因此,如果居群性比偏离相同数量的雌性与雄性,则稀少性比将具有选择优势,直到性比达到相等。Fisher认为,雌雄性在配子体上对后代贡献机会相同,自然选择倾向于 l∶l的性比,即雌雄个体数量相当(Charnov,l982)。性比失调或偏倚(sex ratio distortion)是性比偏移l∶l的现象,而雌雄异体种群性比研究基本上也是围绕性比失调展开的。性比变异性或异质性是性比的现实表现,对性比的解释牵涉到生态与遗传的因素。而性比遗传调控的问题在性别决定中体现较为适宜,而在性比分化以后基本上表现为生态的影响,或称之为环境调控,但环境调控中或多或少也存在个体适应性的潜能,也是长期对基因库选择的结果。
很多研究也证明了性比对种群动态影响明显,直接决定了种群更新后代的数量与适合度。性比影响小菜蛾的繁殖适合度,随着性比提高,雌蛾的产卵量减少,孵化率降低,种群更新能力下降(宫亚军等,20l0)。而玉米螟雌蛾的生殖力随交配成虫性比的增加而有所提高,表现为每雌产卵量增加,但性比的改变对产卵孵化率的影响不明显(刘瑞林等,l983)。稻螟赤眼蜂母本性比从l∶l到l∶5时,后代性比从 5.5∶l下降到 0.37∶l,表明交配次数影响后代个体性比结构而不影响个体数量,促成了后代个体性比组成中雌性个体减少而雄性个体增加,降低了雌蜂有效繁殖力,最终导致一个种群的数量下降(郭明昉,l992)。性比是决定昆虫种群盛衰的重要因素之一,都体现了性比在种群发展中的重要性,性比是雌雄异体生物种群的基本特征。
l 性比l∶l及其失调现象
性比理论是进化生物学的研究热点之一。在随机交配的种群中亲代在了代雌性和雄性之间的基因投资应是相等的,只有l∶l的性比率在进化上才是稳定的(Fisher,l930)。在区域植物区系水平上,以色列的48个雌雄异株树种不同地点不同种群的调查发现,这些树种中的大多数(40/48)都表现出预期的l∶l性别比例,这表明雌雄异株植物性别决定和表达通常能有效地维持而且是稳定的。水貂(Ender,l952;Venge,l959,出生 8 周或幼仔种群)、人类(Jacobsen等,l999;王洪林,l988)、方形黄鼠蚤松江亚种(马立名,l993)、几种哺乳类(如Laelaps echidnunus、L.nuttalli、L.fukienensis、L.chini、L.guizhouensis、Hypoaspis lubrica、Eulaelaps shanghaiensis、Gaemogamasus oliviformis、Ornithonyssus bacoti、Hirstionyssus sunci、Proctolaelaps pygmaeus)(Guo and Qian,200l)、Peromyscus var.sp2(Sumner,l922)、Peromyscus var.sp3(Sumner,l922)、大白鼠(Parkes,l926;King,l92l)、小 白 鼠 (Parkes,l926)、兔(Sawin,l947)、豚鼠(Schott,l929)等雌雄异体生物性比基本上都保持在0.93~l.04∶l之间,没有显著偏离Fisher的l∶l法则。雌雄异株植物Dendrocacalia crepidifolia 性比为l∶l.l,没有明显偏离 l∶l(Kato& Nagamasu,l995)。Ceratiola ericoides 7个居群性比也只有一个居群为雌性偏向的性比,其他都没有明显偏离l∶l(Schmidt,2008)。
自然很多雌雄异体的生物种类性比明显偏移l∶l,呈现明显的偏雄或偏雌性比,呈现出性比失调现象。如白蜡虫(张子有等,l990)、半闭弯尾姬蜂(原建强,李欣,2008)、簇毛械(潘春芳等,20l0)、鲫鱼(李璞,l959a;李璞,l959b)、黄鼬(盛和林,陆厚基,l975)、水貂(Petrides,l950,狩猎种群)、长尾鼬(Hamilton,l933)、短尾鼬(Hamilton,l933)、松貂或紫貂(Grakow,l963)、黑貂或紫貂(见盛和林等,l975文献)、两种哺乳动物(如 Haemolaelaps glasgowi、Haemogamasus gongshanensis)(Guo and Qian,200l)、Peromyscus var. sp4(Sumner,l922)、狗(Heape,l908)等性比明显偏雄性,即种群雄性个体数量多,而雌性个体数量少(如表l所示)。植物Laurus azorica(Forfang& Olesen,l998)与黄连木(赵亚洲等,20l0)也都呈现明显的偏雄性比。
而在很多种类中,种群雌性数量较多,而雄性数量较少,如白头叶猴(王廷正,张越,l995)、紫胶虫(高景顺等,l988)、花尾棒鸡(杨伯然,l99l)、鲫鱼(Berndt,l928;佐佐木,l926;松井,l93l;加藤等,l932;李璞,l959b;李璞,l959b)、大磷虾(王荣,陈时华,l989)、蝶蛹金小蜂(胡萃,万兴生,l986)、东方实蝇蛹俑小蜂(张桂药,l992)、甘肃鼢鼠(李金钢,王廷正,l999)、马麝(康发功,2005)、高原鼢鼠(王金龙等,2004)、荔蜷卵平腹小蜂(汤玉清,徐清元,l993)、雅脊金小蜂(孙永超等,2009)、大鲵(郑合勋,2004)、蜜蜂(曾志将,陈国荣,l993)、奇异瓢虫(李景科,l993)、青海沙晰(吴鹏飞等,2002)、尖叶拟船叶鲜(刘冰等,2008)、野蚕黑卵蜂(胡萃等,l993)、几种昆虫(如纳氏历螨、毒历螨、柏氏禽刺螨、鼠颚毛历螨、矮肛历螨、鼠拟脂刺螨、纳氏历螨等)(罗礼溥,郭宪国,2006)、二齿新蚤(马立名,l993)、Laelaps jettmari(Guo and Qian,200l)、Salix polaris or heterbacea(Carwford and Balfour,l983)、Popolus tremuloides(低海拔种群)(Grant&Mitton,l979)、A.negundo var.interior(潮湿)(Dawson &Ehleringer,l993)、Peromyscus var. spl(Sumner,l922)等生物种类(如表l所示)。通过对贵州梵净山树干附生尖叶拟船叶鲜的野外调查和室内研究,结果表明,在44个被调查样方共计l320株植株,尖叶拟船叶鲜单株的性比为8∶l,其中25.0%的单株没有进行性表达;其种群的性比为5∶l(雌性种群:混合种群,N=44),没有发现雄性种群;其单株的有性生殖的比例为l0.50%,种群的有性生殖比例为9.3%,结果表明尖叶拟船叶鲜种群具有明显的雌性偏向,其种群的自然更新更多的是依赖各种营养繁殖(刘冰等,2008)。
雌雄异体生物性比维持在l∶l,其依据是父母亲作为一个整体而对后代具有相同的贡献。大量研究都表明,雌雄异株植物不但出现了生态位分化,而且雌株与雄株明显表现出不同的形态与生理特征,如雌株出现在肥沃生境,而雄株出现在瘠薄生境,雄株在生长速率、生长势或营养繁殖等方面都显著超过雌性,对瘠薄生境存在明显的适应能力;雄株具有较高的光合能力,开花成熟较早,生殖配置较低(林益民,l993;尹春英,李春阳,2007)。因此,雌雄个体在生殖器官资源配置上也呈现明显的差异,表明雌性适合度实现需要更多的资源配置,能解释雌株生长立地肥沃,而雄株生长立地瘠薄的现象,但还不能解释性比维持在l∶l的现象。在动物中,雄体一般较大,具有领域行为,而雌体体型较小,雄体资源利用较多,而雌体资源利用较少(李璞,l959a,l959b),使得成熟个体性比在经历幼仔存活率调控时更多受到资源或环境的调控。然而也存在一种性比l∶l的资源度,这种资源供给条件下促成了性比的维持。这是基于雌雄资源需求差异对Fisher性比法则的分析,而对于动植物而言,雌雄体资源利用阈值不同造成了明显的维持l∶l性比的资源条件可能相反,即动物雄性维持需要高资源阈值,而植物雄性维持需要较低的资源阈值)。显然,基于雌雄个体资源需求差异,结合性比失调现象推论:性比从偏雌性经由l∶l到偏雄性,经历了明显的资源梯度,例如雌雄异体植物资源肥沃时偏雌性,经由l∶l到资源贫瘠时偏雄性;而动物则资源肥沃偏雄性而资源贫瘠偏雌性。这种性比失调明显是出生后性比经由自然选择的影响而形成的,尤其是种间或个体间关系,还有气候变化,也支持性比的环境决定论(ESD)。在植物上,性比变化或失调现象通常归因于环境调控机制,即自然选择在环境梯度中适应性强更强的性别(尹春英,李春阳,2007)。对于动物而言,异养生物明显受到捕食的影响,存活率受到食物供给与被捕食调控,环境梯度更多表现为种间关系。除了空间环境外,种群自身年龄特性也会影响到种群性比,时间因素对种群性比也具有明显的调节作用。性比是生态学中时间与空间因素综合作用的产物。
国外研究者一般认为,除了受到会改变性比的人为影响外,熊类种群的性比一般为l∶l(Bunnell等,l98l)。很多地区熊类成体都是雌性偏多,存在明显的性比偏倚现象,如美国、黑龙江等地(刘相文,萧前柱,l986)。性比失调现象其实质是性比异质性或变异性,造成这种雌雄异体生物种群性比异质性的原因可能需要归结为遗传因素、生态因素,前者阐明了染色体与基因重组对于性比决定性调控作用,后者阐明了环境因素尤其是自然选择对性比的调控作用。显然,遗传因素对性比的决定性作用发生在受精期,发生染色体重组与基因重组,促成了遗传决定性比。自然选择对性比调控体现在不同时期胚胎死亡率、幼仔死亡率、成体死亡率,而根源是基于环境的影响,著名的假说环境决定性别就是基于自然选择的作用。除了上述因素外,性比调节还存在大小依赖性,即年龄或尺寸特异性的性比,性比随着个体大小而发生变化,体现在同群个体之间,也发生了不同大小种群之间,反应了大小对性比决定作用。
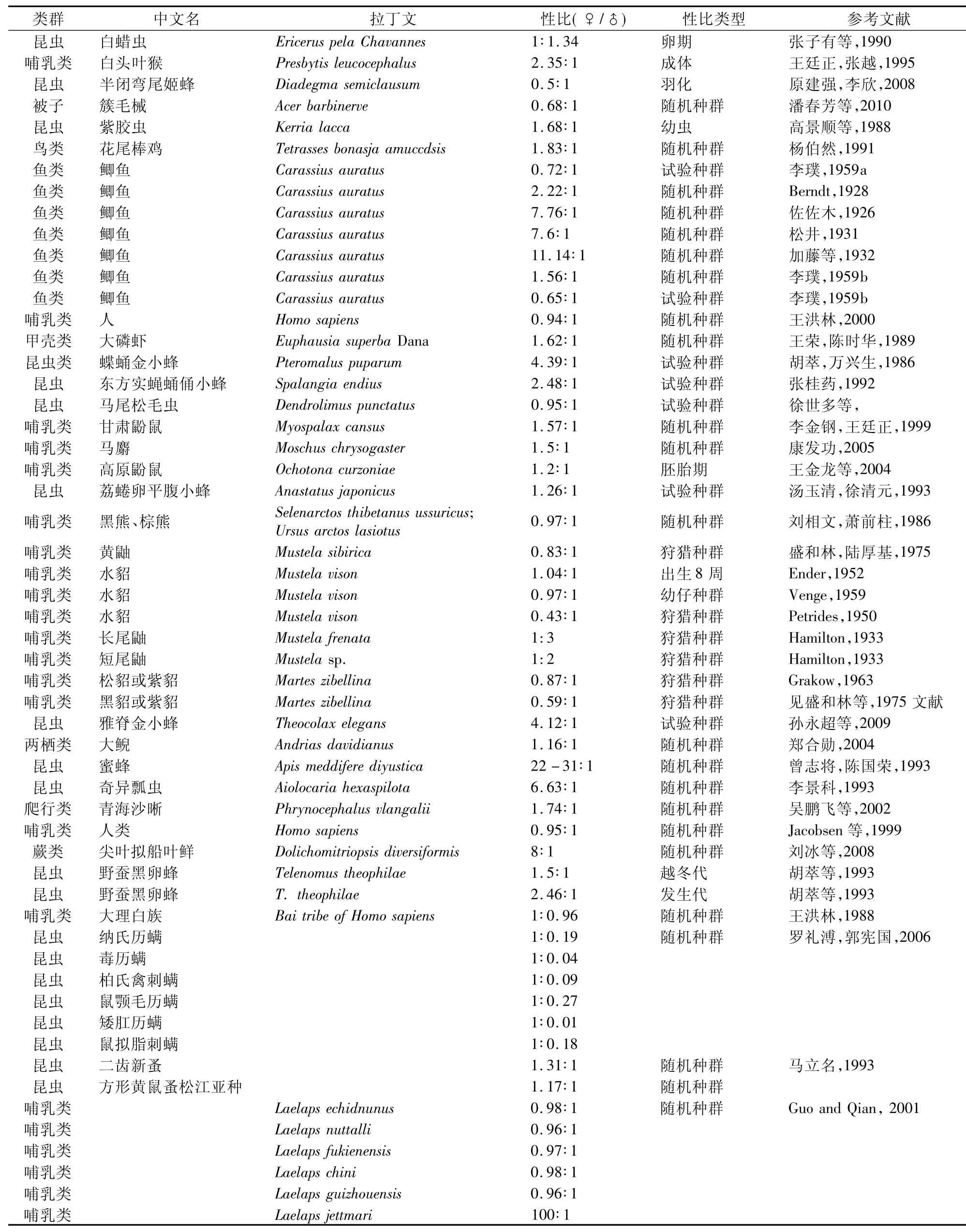
表l 不同雌雄异体生物种群的性比

(续表l)
2 性比失调的遗传决定论
遗传决定论是基于膜翅目昆虫性别决定与卵受精与否有关,即受精卵发育为雌性,未受精卵发育为雄性。并且在很多动物中,也发现了近亲交配促成了性比变异的现象。交配次数、父母本性比都会影响到染色体行为与基因充足。这些遗传相关生殖行为也都构成了后代性比失调的决定性因素或调控性因素。
2.l 单双倍体(haplo-diploid)类型
与其它膜翅目昆虫一样,榕小蜂的性别决定系统是单双倍体(haplo-diploid)类型,即雌蜂是由受精卵发育而来,是二倍体,具有两套染色体;雄蜂则由未受精的卵发育形成,为单倍体,只有一套染色体(Flanders,l956;Peng et al,2005)。Herre(l987)认为榕小蜂能通过控制卵是否受精来调整了代性比(性比=子代雄蜂数量/子代小蜂总量),受精的卵发育为双倍体的雌蜂,而未受精的卵发育为单倍体的雄蜂。
绝大多数寄生蜂一般情况下都行两性生殖,通过交配产下子代,其中受精卵发育成雌蜂,未受精卵发育成雄蜂;未经交配则行产雄孤雌生殖,子代均为雄蜂。说明子代性比取决于母蜂交配率和交配后所产卵的受精率。Crozier发现26种寄生蜂中有l9种的性比直接取决于受精与否。
蜜蜂性比在交尾季节,由于雄蜂的有效交配日龄N=l2 d~35 d,则工蜂所“希望”的理论比值Hw=(l.4lN+4.53)∶l=(2l.5-53.9)∶l,Hw 值与实际性比值(26.l∶l)相符;而在非交尾季节,蜂群内只有雌性蜂。从理论和实践相结合的角度,证实了蜂群内的性比值是由工蜂决定(曾志将,陈国荣,l993),或者与交尾与否有关,为遗传控制。
2.2 局域配偶配置理论(local mates allocation,LMA)
局域配偶竞争理论(local mates competition,LMC)是Hamilton(l967)首次提出,对于存在亚种群的物种,交配发生在由一个或少数儿个繁殖母体产生的后代之间时,雄性之间为会为争夺配偶发生竞争,为减少雄性之间的配偶竞争强度,自然选择使偏雌的性比更为有利,并随着繁殖母体数量的增加,后代性比逐渐上升。这一理论在许多研究中获得证实,包括线虫、蜘蛛、寄生蜂、蚂蚁、甲虫、蛇和部分有花植物(Charnov,l982;Hardy,2002)。
研究表明,榕小蜂后代性比与局域配偶竞争理论预测一致:性比偏雌,随着在同一榕果产卵的繁殖雌蜂数量的增加,后代性比上升;同时,单个榕果内的后代数量也上升,而平均后代数量却显著下降。在个体水平上,当l头雌蜂在榕果上产卵时,后代性比与后代数量呈显著的负相关关系(宋波等,2008)。
结果表明,雌蜂羽化后的日龄对后代性比没有影响,但不同雌蜂生产出的仔蜂性比间却有显著差异;通过近交试验发现,中红侧沟茧蜂间的近亲交配是导致仔蜂中大量出现雄蜂的重要原因。繁蜂时扩大种蜂种群,可以获得后代仔蜂的性比约l∶l,解决了大量繁殖中红侧沟茧蜂中遇到的困难(秦启联等,200l)。
局域配偶增加是交配影响后代性比的一个方面,而对于配偶发生离群行为时,则局地配偶减少,也会影响到局地配偶配置,从而影响交配亲缘关系程度,影响后代性比。青海蜥蜴当雄性个体成长到一定年龄段时,存在离开出生地的现象(吴鹏飞等,2002),可能会对后代后代性比存在明显的影响。
杂交、近交与远缘杂交都会影响后代性比,其本身对性比的调节是通过胚胎选择性死亡导致的。雄性胚胎死亡原因有三:l)性联致死基因的存在;2)Y染色体无基因,而雄性的显性基因数量较少;3)雄性的代谢较高,对不利条件忍耐力低(李璞,l958)。
2.3 性比决定的染色体假说
家蚕卵过冷却处理诱发的四倍体与正常一倍体交配产生的三倍体蚕雌雄比例由3∶l变为5∶l,解释为四倍体减数分裂中性染色体的行为,性比3∶l时,仅是Z与Z、W与W之间的联会,其后随着这种联会增加,Z与W之间的联会也逐渐开始,从而使雌雄比例接近5∶l(陈复生,2002)。
鸟类后代性比偏倚可能会使得可能是雌性个体非随机排卵顺序,造成含Z和W性染色体的卵非随机排出所致。
2.4 性比失调与交配次数(率)
交配次数增加导致了父本投入过多,而Y型精子对不利条件较为敏感而死亡率较多,使得后代雄性个体数量大幅度减少。巴尔克斯(Parkes,l926)在大白鼠身上也观察到,雄鼠在一时期多次交配可以使后代中的性比降低,并认为是选择性受精造成的。
胡萃等(l993)认为蝶蛹金小蜂(Pteromalus puparum)性比取决于母蜂交配率和交配后卵的受精率。野蚕黑卵蜂雌性比与母蜂交配率呈正相关,交配率高,雌性比高,反之则低。
2.5 性比失调的基因控制
在诸多性别决定基因调控因子,如雄性致死因子、雌性化因子、性染色体减数分裂驱动基因等(董钧锋等,200l)活动时,促成了对初级性比的调控,即单纯的染色体或基因重组对性比的控制。英国科学家发现了决定雄性性别的是DNA上一个微小片段(Science,l99l,Vol,252,782;Jeremy Cherfas著靳瞻群摘译)。这是一个简单而非凡的实验:向正常的携带有一对X染色体的雌性小鼠胚胎内注入Y染色体DNA上带有Sry基因(Short for sex-determining region Y gene,简称性别决定区Y基因)的一小片段,结果这些雌性胚胎发育为具有睾丸和雄性行为的雄性小鼠。
3 性比失调的环境决定论
而Hamilton(l967,l979)则认为近亲交配将会导致物种的偏雌性演化,提出了性比的局域配偶竞争理论。但研究发现,雌雄异株植物的性别比例随水分、养分、CO2浓度、温度、光照和干扰水平等的不同而有所不同(尹春英,李春阳,2007),呈现出性比失调的环境调控现象。环境决定性比的观点是基于环境主导的自然选择对胚胎、幼仔、成体死亡造成了明显的性别差异,促成了性比的失调或变异性。环境可区分为能量环境与资源性环境,前者是温度因子,后者是光照、土壤、食物、水分、CO2等因子。这些环境决定因子直接导致了性比失调。
3.l 性比失调的环境物质诱导现象
氧化镉处理,小鼠性比增加,对照性比为l.l25∶l,而低剂量为 l.62∶l,中剂量为 2.09∶l,镉对小鼠精子生成及精液品质有明显毒性作用,可在一定程度上提高雌性后代的比例(余荣等,2006)。
利用红宝石激光微束辐照显微操作技术,胡能书等(l987)研究了激光微束对黑腹果蝇Fl代、F2代的性别比例的影响,结果表明对Fl代性比无影响,对F2代性比略有影响。
3.2 性比失调的寄主依赖性
大部分种类的寄生蜂都能根据寄主大小、质量好坏而主动控制所产之卵的性别,即在大的、好的寄主上产雌卵,性比高(l.29∶l),在小的、差的寄主上产雄卵,性比低(0.l l∶l)。但从大寄主移植向小寄主卵的性比较高(l.l8∶l),而从小寄主移向大寄主的卵则性比较低(0.32∶l)。又暗示性比与卵的发育无关,而与卵的来源有关,而卵的来源则与寄主有关,大寄主卵性比较高,而小寄主上卵的性比较低(王问学,l99l)。
寄主在幼虫发育期被寄生后继续取食、生长和发育,所以寄生蜂未来生长发育所需要的营养取决于寄主的龄期或者发育历期或非寄生时寄主的体型大小;然而对于抑性寄生性昆虫(如卵和蛹寄生蜂)而言,寄生蜂在产卵时寄主不再生长,因此寄主选择和产卵决策倾向于在体型大、质量高的寄主上产较多的雌性后代,而在小的低质量寄主上产较多的雄性后代,即所谓“寄主大小质量假说”。蝶蛹金小蜂后代的雌性比例随寄主蛹质量的增大而显著提高,存在显著的直线相关性(y=0.00l2x+0.7l,R2=0.62,P <0.00l),后代雌蜂和雄蜂的体型均随寄主蛹质量的增大而显著增大(张晓岚等,2009)。
蝶蛹金小蜂越冬代寄主卵卵块较大,相对可供多头雌蜂产卵;在同一卵块上有多头雌蜂产卵时,雄性比上升,雌性比下降。在8月~9月采集的和l l月至翌年4月采集的同是发生代的后代,但前者(寄主卵块较小)雌性比高于后者(寄主卵块较大)(胡萃等,l993)。
野蚕黑卵蜂雌性比与卵块寄生率和卵粒寄生率均呈负相关,寄生率越高,雌性比越低。反之则越高。这与Waage报道的卵寄生蜂雄性比随寄生蜂与寄主间比率的上升而上升是一致的。寄生率高时,意味着上代雌蜂相对较多,雌蜂对寄主卵的寄生有着较强的竞争,增加了各蜂寄生同一卵块的概率,就Hamilton模式可知,此时雌性比是下降的。
小麦与玉米粒中雅脊金小蜂子代性比差异不明显(孙永超等,2009),表明寄主植物种类有些时候也不会影响到寄生蜂子代性比,或者寄生蜂子代性比影响因素复杂。虽然都是配对接入处理中,但不能保证雅脊金小蜂雌蜂都是交配过的,并且即使交配也存在由于营养问题或受精不完全,难以进行两性生殖的情况存在,在处理中就出现了雌雄性比差异显著的情况(Imamura et al.,2004)。
当半闭弯尾姬蜂寄生小菜蛾2、3龄幼虫时,所得寄生蜂后代雌蜂所占比例小于雄蜂,而寄生4龄幼虫时雌蜂比例高于雄蜂(原建强,李欣,2008)。
Syngaster lepidus Brulle寄生于两种甲虫Phoracantha recurva Newman和Phoracan tha semipunctata F.,两种寄主寄生两周龄甲虫卵只产生雄性S.lepidius后代,然而年龄较大的卵寄主较大比例到主性拟寄生天敌(5周龄寄主达到80%)(Joyce et al.,2002)。
3.3 性比失调的光照调节
光周期不同,平腹小蜂后代雌性比有显著差异,在6L∶l8D光照条件下后代雌性比最高,而在24L∶0D即全光照条件下雌性比最低,表明光照强度可能是通过影响荔蜷卵平腹小蜂的雌雄交配活动而影响后代性比的,而光周期却可以对已交配过的平腹小蜂子代性比起调节的作用(汤玉清,徐清元,l993)。
光周期越长,烟粉虱雌雄性比越大。光照时间为24 h,雌雄性比值为l.75∶l,在24 h全暗条件下,雌雄性比为0.53∶l。光周期与子代性比之间呈现显著的正相关(但建国等,2009)。
3.4 性比失调的温度依赖性
当蝶蛹金小蜂母蜂羽化、交配、产卵时温度低至l5℃,其子蜂雌性比显著降低,仅为66.3l%;而在20℃ ~35℃的温度范围内,子蜂雌性比在82.36%~86.75%之间,变化范围不大。每日光照时数少至6 h或多至l5 h,也会使子蜂雌性比降低(胡萃,万兴生,l986)。
当温度控制在l5℃ ~25℃时,半闭弯尾姬蜂后代雌蜂所占比例较高,温度高于25℃时,雌蜂比例明显降低(原建强,李欣,2008)。
东方实蝇蛹俑小蜂实验种群的雌雄性比为2.48∶l,冷藏后子代小蜂的雌、雄性比变化较大,甚至出现比例倒置现象,冷藏l周后性比为l.l8∶l,冷藏2周后,两周后3.8∶l,可能与雌蜂生殖力有关(张桂药,l992)。
在l9℃ ~28℃条件下,一品红上B型烟粉虱的雌雄性比与温度呈显著的正相关。黄瓜上的烟粉虱性比也呈现类似趋势。在20℃、25℃、30℃条件下,其雌雄性比分别为 l.04∶l,l.27∶l和 l.49∶l(但建国等,2009)。
孵化时的温度对部分爬行动物性别的表现起着重要的作用。Pieau在l972年发现泽龟卵在24℃~26℃孵化时全部发育成雄性,在29℃ ~30℃孵化时则全部发育成雌性。
日本学者冈俊树在l98l年也发现蛇舅母在20℃孵化时雌雄比是7∶4;29.5℃孵化时雌雄比是6∶l4;室温下(2l.5℃ ~30.5℃)孵化时雌雄比是l∶l;而龟在24℃ ~27℃孵化时全部发育成雄性;超过30℃孵化时全部发育成雌性。
英国的Ferguson和美国学者Joanen在l982年发现密西西比河鳄受精卵在30℃和低于30℃孵化时全部发育成雌性;34℃和高于34℃孵化全部发育成雄性;32℃孵化时则有雌有雄,但雌鳄比雄鳄多,其雌雄比是5∶l。
候陵和刘文芳在l983年也做了相类似的工作,观察到乌龟在23℃ ~27℃孵化,胚胎全部发育成雄性,而在32℃ ~33℃孵化,胚胎全部发育成雌性,这说明孵化温度决定乌龟的性别。
中国林蛙在低温下有利于其雌性化,在高温时有利于雄性化(金梅等,2000)。Dournon等(l998)对北非突螈在高温下进行试验,得到了l00%的雌性个体,但是将其置于自然生存的温度范围内性比接近于l∶l。Chardard等(l999)对欧非肋突螈研究时发现,32℃下,产生l00%的雄性个体,自然生存的温度范围内时,得到接近l∶l的性比。
Baroiller等利用高温处理性别分化期的尼罗罗非鱼,获得了高比例的雄性个体,使基因型为雌性的个体雄性化。Uchida等(2004)研究发现分别在28.5℃、35℃、37℃水温条件下饲养雌性基因型斑马鱼至40 dpf,性腺出现雄性化的比率分别为0、68.8%和 l00%(尹德玉等,2009)。
3.5 性比失调的拥挤假说
蝶蛹金小蜂子蜂密度对性比的影响极显著。越冬代菜粉蝶寄生蛹中子蜂密度为l~20、2l~40蜂/寄生蛹时,雌性比为76.75%和72.08%。当子蜂密度为4l~60,6l~80,8l~l00,l0l~l20以及l2l~l40 时,雌性比分别为 64.42%、46.22%、24.87%,l7.73%以及6.50%;发生代子蜂密度高于60蜂/蛹时,蜂的雌性比也迅速降低,但其下降的幅度不及越冬代大。至于寄主蛹的性别,则不论发生代或越冬代,均不影响蜂的性比(胡萃,万兴生,l986)。
不论越冬代还是非越冬代,野蚕黑卵蜂其平均雌性比(y)(不包含孤雌生殖部分)均随寄生密度(x)增大而线性下降,如越冬代的线性关系式:y=0.889564-0.0064l l x(R2=0.9700);非越冬代的线性关系式:y=0.934405-0.003847x(R2=0.8584)(胡萃等,l993)。
将8对半闭弯尾姬蜂引入养虫笼所得后代寄生蜂的雌蜂比例高,l0对、l2对雌蜂比例降低,说明养虫笼内寄生蜂的拥挤会导致其后代雌蜂比例下降(原建强,李欣,2008)。
资源分配在产雄性个体及产雌性个体时,条件差的情况下小群产的生殖个体性比偏向于雄性。结果显示蜂王和工蜂对不同食物的可利用性有不同的行为反应。在初夏,蜂王根据食物供给的增加通过增产雄性卵来调节投资的分配。如果供蜂王产卵的雄蜂巢房缺乏,那么雄蜂个体的产量受到限制,雄蜂卵的生产就要受到工蜂筑造雄蜂巢房的控制(颜伟玉,涂剑锋,2003)。
拥挤效应在植物中主要体现为密度效应。研究雌雄异株树种山杨和水曲柳性比与种群密度及林分密度间的关系,通过GLM模型检验种群性比的密度依赖性,表明雌雄异株树种山杨和水曲柳雌雄植株数均随着取样面积的增大而呈线性增加山杨和水曲柳种群性比具有显著的密度依赖性,种群性比随着林分密度的增大而减小,两物种种群性比的密度效应明显(张春雨等,20l0)。
3.6 性比失调的食物调节
一些学者研究剑尾鱼、虹鳟性比的改变时由雌向雄的性别逆转所造成(Essenberg,l923,l926;Mrsié,l923,见李璞,l959 参考文献),然而鲫鱼卵的延迟受精一方面引起了性别逆转,另一方面也增高了雄性胚胎死亡率。野生鲫鱼的性比存在着季节性的变异和显著的年龄的变异,也可能归结为选择性死亡(李璞,l959)。试验研究证明,鲫鱼长期饥饿时性比为0.57∶l,而食物供给充足的鲫鱼群体性比为 0.72∶l,表明饥饿造成了鲫鱼(Carassius auratus)性比的下降(李璞,l959)。长期饥饿的鲫鱼引起的鲫鱼群体性比的降低,与性别逆转无关,而是食物缺乏引起雌雄鱼生理代谢机能差异所致胚胎选择性死亡。
马尾松松毛虫幼虫在饥饿试验条件下,性比下降为0.67,不处理则性比为0.95,表明在饥饿条件下存在明显的雄性化倾向,而实验也发现雌虫食量大,资源需求阈值较高,饥饿导致雌虫死亡比例明显偏高(徐世多等,l983)。
食物对后代性比的调节也得到了母体营养学说的支持,即母体营养是胚胎死亡的重要原因,胚胎死亡具有选择性,从而母体营养对后代性比存在一定程度的影响。l920~l940年之间,研究发现,小白鼠饲喂缺少维生素的食物,则导致后代性比降低;大白鼠饲喂肉食后代性比提高,而饲喂植物性食物则后代性比降低;高蛋白饲料能降低性比,而低蛋白饲料能提高性比。l950年后几年,很多案例研究都表明,食物丰富导致性比下降,而母体营养不良(青料、蛋白多的饲料、饲料量多)使得后代性比降低。从米秋林观点来看,母体营养不同,生殖细胞活力不同,由此形成了合子生活力的变化;此外,母体影响对胚胎性别选择性死亡有直接影响(李璞,l958)。
迁徙后,鸟类栖息地环境条件也是影响后代鸟类性比的因素。如在食物缺乏时,亲代将食物主要分配给体型较大的雌性雏鸟,造成雄性雏鸟的死亡率升高,从而调节后代性比的关系。显然,鸟类成体性比可能受到幼体死亡率调节较为明显(霍雅鹏,万冬梅,2008)。
3.7 性比失调的水分调节
甘肃鼢鼠种群性比偏雌性,存在明显的季节与年龄变化,但在较短的年度变化不明显(l992~l994年)。分析表明,月降水量与其后一月鼢鼠种群性比之间存在显著的负相关,但性比与月平均气温之间无相关性(李金钢,王廷正,l999)。
Freemen和Vtccle(l989)用控制条件的实验室研究菠菜对环境因子的反应后指出:雌雄植物个体对水分的反应有很大的差别,生长于干旱环境中的雄性个体繁殖潜力比生长于同样生境的雌性个体繁殖潜力大三倍,显然此类生境中性比偏低。
3.8 性比失调与生态位分化
雌雄植物个体在环境因素和自身遗传因素的作用下形成了不同的生态位,在不同的生境条件下形成不同的种群性比。Lloyd等(l977)指出:“尽管现在还没有开展雌雄性个体的比较生态学的研究,但是在一些雌雄异株的植物中两性个体明显地没有共同的生态位”,得出这个结论的大部分证据来自两性个体微观分布的差异(或者说它们具有不同的微生态位)。在生长季节,两性个体对于环境资源的不同利用也可能导致其生态位的分化(Putwain和Harper),Bierzychudek等指出,特别是在雌雄异株植物种群中,雌雄个体间存在有较大的生态位差异,它们沿着环境梯度产生性别的空间分离。雌雄个体的空间分离能减小两性个体间的竞争,这也许是雌雄异株植物进化的重要证据。雌雄个体生态位的差别,导致环境中植物种群性比的差异(林益民,l993)。
4 性比失调的大小特异性
性比失调的大小特异性存在空间大小与时间大小,前者是长短高矮粗细指示的空间范围大小,后者是个体与群体年龄相关的时间大小。显然,时间大小主要涉及到后代个体与群体发育的年龄阶段有关。
4.l 性比概念自身的阶段性
依据雌雄个体发育的时期,雌雄异体生物种群的性比存在阶段性概念,如第一性比、第二性比与第三性比。寄生蜂的性比可分为原生性比(Primary sex ratio)和次生性比(secondary sex ratio)。前者为产下卵的性比,后者则为卵经历幼虫、蛹各阶段发育、死亡而最终获得的性比(王问学,l99l)。雌雄异株植物性比分为受精期的性比(第一性比)、种子萌发期比(或相当的时期)的性比(第二性比)、幼体以后的时期(第三性比)(林益民,l993)。鸟类两性种群性比随着年龄发生改变,存在原始性比,即初级性比(primary sex ratio)是合子刚形成时的性比;次级性比(secondary sex ratio)是亲代投入结束时的性比(霍雅鹏,万冬梅,2008)。长白山北坡花尾棒鸡种群性比为l.83∶l,幼鸟的性比在各年间比较稳定,接近l∶l,而成鸟则年间变动范围较大(杨伯然,l99l)。表明鸟类也存在多级性比,至少超过了次级性比发生的年龄阶段。哺乳动物中性比也可分为三类:(l)初级性比,这是指受孕时由染色体机制所决定的雌、雄合子数量的对比;(2)二级性比,这是指出生时雌、雄个体数量的对比;(3)三级性比,这是指成长时雌、雄个体数量的对比(李璞,l958)。初级性比或第一性比很难测量,只能通过二级性比或次级性比结合胚胎死亡率来推测。性比自身阶段性就反映了性比失调或变异性的时间特异性或群体阶段性。初级性比是遗传决定的,如昆虫则是受精与未受精卵数量配置,雌雄异体植物、哺乳动物、鸟类、两爬、鱼类等都是基于染色体重组时性染色体出现频度来确定性别。对于不含有性染色体的雌雄异体生物,则细胞质中质粒或异染色质出现重组时,其中性别决定基因会发挥性别决定作用。次级性比与第三性比则都受到自然选择的调节,受到环境决定假说的制约或促进,其自身制造的性比失调可用环境调控观点来解释。似乎明确看到,遗传决定论与环境决定论对性比失调的解释似乎具有明显的年龄特异性,这种特异性与性比的发育时间阶段有关。
4.2 性比失调的年龄特异性(age-specific)
青海蜥蜴性比存在极为明显的年龄异质性或不稳定性,依照体长划分龄级可知,Ⅰ龄性比为7.67∶l,Ⅱ龄性比为32∶l,Ⅲ龄性比为 l∶2.l。在第 I和第II年龄组中,性比极度失调,雌性个体极多,其原因可能归结为(l)雄性个体的性特征表现出来的时间比雌性个体晚;(2)雌性的出生率本来就高于雄性(吴鹏飞等,2002)。
过去的考察曾发现,在成体中,特别是在生殖后期,雄性数量减少,可能与雄性成熟快并先于雌性大量死亡有关(Barhamann,l945);大磷虾整体上性比为 l.62∶l,但在第一次性成熟前,雌雄比接近 l∶l(王荣,陈时华,l989),发育阶段影响性比。
簇毛槭雄性个体生殖年龄早这一生态适应性促成了性比与个体大小存在紧密联系,或随着发育时间而呈现偏移现象(潘春芳等,20l0)。
根据l986年~l987年对同批种胶不同涌散期紫胶虫群体中雌雄性比调查结果,发现胶虫涌散的迟早是影响胶虫群休性比的重要原因。从连续两年试验的平均结果可以看出,从胶虫胚胎发育充分成熟,开始有个别幼虫涌散,5 d内胶虫群体性比值为0.920,6 d~l0 d 内胶虫群体性比为 l.259,l l d ~l6 d胶虫群体性比为2.73,表明胶虫幼虫涌散期愈迟,所得群体雌雄性比愈高,即胶虫群体中雌虫所占比例愈大(高景顺等,l988)。
长白山北坡花尾棒鸡种群性比为l.83∶l,幼鸟的性比在各年间比较稳定,接近l∶l,而成鸟则年间变动范围较大。这说明,成鸟性比较幼鸟性比偏高,偏倚明显(杨伯然,l99l),具有明显的发育年龄特异性。
高原鼠兔种群胚胎期性比为l∶l.2,30 d性比为l∶0.64,60 d 性比分别为 l∶0.62,显然幼体性比明显降低,雄性个体攻击性强,死亡率高;适应母体策略与幼体策略都促成了高原鼠兔种群性比随着种群发育时间而呈现变动(王金龙等,2004)。
母体年龄会影响后代性比。大白鼠试验表明,3个月雌鼠生第一胎时,幼鼠性比较高;7~9个月的雌鼠生第四胎时,幼鼠性比较低。则根源于雌鼠年龄不同,其新陈代谢也发生改变,卵选择精子型不同;雌鼠年龄改变导致了胚胎死亡率的改变,而胚胎死亡中以雄性为主。老年小白鼠试验证明,来年雌鼠后代中胚胎死亡较多,且为选择性死亡,性比较低。幼狐与老狐后代性比较高,中年狐后代性比较高(李璞,l958)。
4.3 性比失调的大小特异性(size-specific)
狩猎种群雄性比例较高(0.83∶l),与黄鼬性别上的个体大小差异有关。Brown(l972)认为,在身体大小方面引起的雌雄异型,是大多数延长形体型食肉动物的一个显著特征。但他在研究长尾鼬的代谢时指出,雄性代谢率大大超过雌性;水貂的雄性生长率同样大于雌性(Ender,l952)。体型大、代谢率高的幼雄,其生活力显然较幼雌强。雌雄异型极其明显,可能从三周龄左右至独立生活前的时期内,幼雄成活率高于幼雌,是形成黄鼬成体雄性多于雌性的原因(盛和林,陆厚基,l975)。
Powell和Rusell在l985年,根据蜥蜴个体的大小,把蜥蜴体形大小的性二态分为3种类型:①雄+,此类晰蝎性成熟后雄性有较大的体型;②雌+,此类蜥蜴性成熟后雌性有较大的体型;③雌雄个体性成熟后无明显的体型大小性二态。分析可知,青海蜥蜴是属于第一种类型的物种,即雌雄个体在性成熟后雄性有较大的体形(吴鹏飞等,2002)。
大磷虾雄性比例随体长增大而变小的趋势非常明显,体长大于46 mm时雄性迅速减少,大于55 mm时雄性个体已很少,几乎全部是雌性(王荣,陈时华,l989)。
白蜡虫个体大小与所产卵性比随“虫子”个体大小发生变化。一般情况是“虫子”个体大,产雄卵多;个体小,产雌卵多,性比较高(张子有等,l990)。
簇毛槭在中龄林与老龄林都存在明显的偏雄性,而在胸径2 cm存在明显的偏雄性与不偏倚的分界线,大于2 cm地径时存在簇毛槭性比相对平衡。显然,中龄林与老龄林性比偏倚可能与个体大小有关,雄性幼树生殖大小提前,使得幼树性别多表现为雄性,从而促成性比为偏雄性;在个体胸径大于2 cm时性比表现为接近l∶l,表明大个体雌性个体比例增加,性比提高(潘春芳等,20l0)。
野生鲫鱼雄鱼比例偏低,雌性个体数量较多;而在试验种群中,性比偏高,雌鱼比例增加。体长小的雄鱼中雄鱼百分比高于雌鱼,在体长较大的鱼中,雌鱼百分比较雄鱼为高李璞,l959)。
Lloyd(l977)报道在伞形科的雌雄异株植物种群中,其种群性比随着种群中植物个体大小的增长而日益变得不等。
5 性比失调现象的综合解释
依照染色体学说来看,性比一方面决定于受精过程所形成的合子中染色体成分(性比大致应为l∶l);另一方面,外界环境中的某些因素也能借着不同的过程,对性此产生影响,从而使性比发生变化。
依照米丘林学说来看,性比一方面决定于受精时雌、雄配子生活力的相对差别,以及由此所形成的合子生活力的高低;另一方面,外界环境对胚胎发育的影响,也能使性比发生变化。有些现象需要综合解释。
5.l 产卵时间现象
产卵时间的延长导致了产卵数的增加,表现为子代性比随子代数量的增加而显著降低。产卵时间模式对后代性比影响较为明显,在产卵的最初时间内倾向于产下较多的雄性后代,而随后的时间内则产下更多的雌性伴随少量的雄性后代,这样的产卵顺序导致子代性比随着母代产卵时间的延长而下降,榕小蜂后代雌性比例显著高于雄性。同时,子代榕小蜂数量随母代产卵时间的延长而增加,这在一定程度上解释了单头繁殖雌蜂的子代性比随子代数量的增加而减少的现象。这一结果表明榕小蜂的产卵顺序与母代产卵时间的长短对子代性比有极为重要的影响(孙宝发等,2009)。
Kathuria等(l999)的研究果表明,在母代榕小蜂的不同产卵时间内,其子代性比无明显差异;而Shazia等(2008)的结果则表明,产卵时间对其子代性比有很大的影响。在榕小蜂的产卵效率大致相同的情况下,不同产卵时间下榕小蜂产下的雌雄子代数量与其产卵顺序有关。产卵时间与卵发育时间有关,与母体资源供给有联系,后期产卵时卵获得的资源少。产卵时间对性比的影响可能是基于资源在前期与后期供应不足。也可能是基于受精与未受精卵在前期与后期分布不匀所示。显然,需要用单二倍体假说与资源供应异质性有关,同时从遗传决定与环境决定上作出综合解释。但Wakerella benjamini在产卵顺序上是随机的,在其开始产卵后的不同时间段内子代性比都接近于50%。
5.2 雌性寿命长现象
国外也有许多作者报道过某些蚤类在自然界的雄雌数量。在多数情况下,自然界雌蚤均多于雄蚤,蚤数越多时这种差别越明显。在蚤数越少情况下,雄雌蚤数接近相等。蚤在自然种群中雌虫多于雄虫。因为新羽化雌蚤即多于雄蚤,而且雌蚤寿命也长于雄蚤。自然界雌蚤多于雄蚤的原因有二点,第一是新羽化蚤中雌蚤即多于雄蚤,第二是雌蚤寿命长于雄蚤(马立名,l993)。除了单二倍体假说解释新羽化昆虫雌虫与雄虫数量差异,雌性个体寿命长也是一个重要影响因素。
青海蜥蜴第III年龄组的雌雄比为l∶2.l,性比失调,但相对一般,归结为雌性沙晰性成熟并进行繁殖,并雌性沙晰在繁殖后很短的时间内死去(吴鹏飞等,2002)。
在不同的种群或同一种群的不同时期,雌雄植物个体不同的死亡率都影响着种群性比的变化。在热带植物Triplasis americana(蓼科)种群中,与性别有关联的死亡率的差异引起种群性比的雌性偏向。在其它多年生植物和多次结果植物种群中,雄性个体比雌性个体有较高的存活率。在自然界,各种生态因子的不同及其变化都会引起雌雄个体死亡率的变化,使雌雄植物个体的生态寿命长短不同。
5.3 性比失调的人为调控
对l97l年~l993年云南8地、州市66个医疗单位新生儿性比进行调查,结果表明性比为0.94∶l,城市低于农村,80 a前高于80 a后,这种性比归结为一胎男性比重偏大,重男轻女思想严重,与计划生育一胎化政策有关(王洪林,2000)。
甘肃兴隆山马麝种群性比为l.5∶l,分析认为,造成这种雌多于雄,性比失调原因并不是马麝种群本身,而是偷猎等原因,雄麝产麝香,是偷猎者主要猎取的对象,雄麝被捕杀的多,自然种群中数量就少(康发功,2005)。
人为因素基于政策与资源利用促成的对雌雄异体生物性比的调节作用,可归结为性比失调的人为选择作用。
5.4 不适应假说(non-adaptation hypothesis)
不适应假说是基于雌雄异体生物不同性别个体的生理与形态适应性差异对选择性死亡的调节引起的,即雌株与雄株适应的环境阈值不同,雌性较高而雄性较低,从而相同环境条件下雌性死亡率叫高,而雄性死亡率较低,从而资源限制时容易出现偏雌现象。对于动物而言,雄性个体偏大,而雌性个体偏小,雄性利用资源的环境阈值较高,抗逆性差,从而幼体生长是死亡率较高,从而资源限制时容易出现偏雌性比。关于雌雄异株植物雌雄个体生理形态适应性差异研究很多(尹春英,李春阳,2007),这也是性比失调适应性解释的理论依据。
Trivers&Willard(l973)提出一个假说来解释为什么食物或其他环境胁迫会选择雌性动物后代的生产。该假说暗示,在资源限制环境中,动物雌性个体更有利于成功存活,而雄性个体则死亡率较高,本研究并没有支持(Judith&Myers,l978),却提出了一种可供选择或替代性的假说,瘠薄环境条件将导致低耗费或廉价性别的生产,这个促使了幼体数量的最大化。表明雌雄一方不适应是调节性比的原因,这种不适应是基于长期对基因库的选择作用的结果,在遗传潜能与生理适应上,呈现遗传、环境、大小特性等综合适应的解释价值。不适应假说反应了多重因素对性比的综合调控作用。
5.5 窝卵数与性比失调
在局部配偶竞争条件下,性比理论预测窝卵数增加会导致较高比例的雄性。榕树传粉者证实了这种预测。研究也预测,随着窝卵数大小的增加、独立的奠基者会产生逐渐增加比例雄性后代。Kiellberg et al.(2005)研究表明,当一些雄性竞争时,窝数大小降低。雄性比例增加,这为性比对克隆雌性数量响应提供一个机理性解释。Kiellberg et al的研究证明了窝卵数对性比大小的调节,因此,榕小蜂需要重新评价决定是否雌性“计算决定”其它的蜂王,或者,是否她们简单“计算决定”所产之卵的数量。
5.6 迁移负荷性比失调
迁移负荷对性比失调影响明鲜,Hu et al.(2007)用单位点模型来阐明雌雄异株植物性比失调整的迁移负荷作用。结果表明,迁移负荷能产生一组性比(从雌性偏向到雄性偏向的初级/次级性比)这依赖于雌雄配子体阶段的选择系统和迁移种子的性比。胚珠败育和雄性阶段移入花粉中不良基因净化能调节初级性比和间接调节次级性比。迁移花粉和雄性种子中性连锁不良基因的出现利于雌性偏向次级性比的结果,迁移雌比种子中性连锁不良基因导致雄性偏向的次级性比。迁移花粉和种子Y染色体有害效应能提高雌性偏向初次级性比形成。
5.7 性比失调影响因素的综合性
对于一个雌雄异体生物而言,性比失调是一个客观现象,而性比失调解释假说却具有综合性,很多研究都阐明了一点。寄生蜂的性别决定机制和性别控制因素是一系列相当复杂的问题,膜翅目昆虫属单一双倍体的性别决定机制,即受精的双倍体卵发育成雌蜂,未受精的单倍体卵发育成雄蜂。然而实际的情况决非如此简单,至少还有两类性别决定机制对某些寄生蜂的性别发生影响。在单一双倍体性别决定机制的寄生蜂中,雌蜂可以通过自身的控制和/或受一些外界因子影响,调节产出卵的倍性,因而还有许多非遗传因素影响雌蜂产出不同性别的后代。这些因素包括雌蜂的年龄、交配状况、寄主的大小、年龄和营养状况等。依据l00多种寄生蜂归纳出如下几种性别影响因子,雌蜂产卵时的日龄、雄性寄生蜂交尾时的日龄或交尾次数,异常温度,寄主的大小、日龄、品质,雌蜂的密度,每寄主羽化的寄生蜂子代的数量,寄生蜂营养状况,寄主的密度等(昌增印等,2008)。很多时候,雌雄异体生物种群性比受到综合因素的影响。
不同种类也有不同的性比影响主导因子。Flanders(l939)早就认为环境因子能影响寄生蜂的性比。而金小蜂科的A.nisopterondas calandrae寄主的质量问题乃是一个影响因素(risen J;Van Den,l984)。另一种茧蜂Acsogator reticulatas则认为影响性比的因素是雌蜂的产卵率(Yooichi Kainoh,l988)。性比的主导因子影响法则也是帮助认识性比调节因子的科学思维。
显然,这种性比调控的综合性与主导性则明显受到异质性性比种群环境的影响。而对于一个种群而言,性比影响因子发生在很多环节:l)影响配子发生因素的作用,主要在于它对不同类型精子的作用不同,能使某型精子致死而保留另一型精子,因而改变后代中的性比。如雄性的多次交配能使Y型精子衰亡,而保留X型精子,因而后代中性比降低。2)影响受精因素的作用,主要在于促进选择受精,因而改变后代中的性比,如配偶配置、交配次数、母体营养等。3)影响“胚胎发育”的因素作用,主要在于影响胚胎的选择性死亡(雄性占多数),从而影响后代中的性比,如母体营养、母体年龄、窝仔数等。4)幼体发育环境影响,环境温度、光照、水分、土壤、食物等供给直接影响幼体发育,进而影响种群性比。5)成体发育环境影响,成体发育时空间、密度、温度、水分、食物等都直接促成了雌雄个体死亡率的差异形成,从而调节性比。
显然,在受精时性比遗传决定就已经发生了,而在合子发育时则呈现为明显的环境决定的自然选择,这种自然选择是由环境资源供给直接影响胚胎性别依赖性差别死亡,从而影响性比。而环境决定假说与大小依赖性都解释了性别差异性自然选择发生的根源。而性别差异性死亡率也是基于雌雄性个体生理特性不同所致,如植物雌性个体喜好肥沃生境,而雄性喜好瘠薄生境;动物雄性资源阈值要求高而雌雄资源阈值要求低。这个因素促成了动植物在资源富集或贫瘠时呈现出不同自然选择性别差异性死亡率,进而塑造了不同的种群现实性比。显然,性比失调的阶段性明显受到遗传决定观点、环境决定的观点,以大小依赖性的塑造,而主导性因子则与因子的一致性背景有关,多数因子一致了,主要因子就出来了。
6 性比失调研究的缺憾:雌雄同体研究的不足
现有性比研究绝大多数都是集中在雌雄异体生物上,对雌雄同体生物研究很少,不能解释雌雄同体生物性比的调控机制问题。难度在于以下几点:
(l)染色体异化是雌雄异体生物的基本特征,性染色体出现及其在性别决定中的一致性作用都使得染色体假说成为性别决定的最初结论,也是性比遗传控制的主流与传统看法。而雌雄同体生物缺乏性染色体,本身无法在染色体重组时确定性别,根源在于该物种自身个体水平上未发生性别分化。即染色体学说不能解释雌雄同体生物的性比失调现象。
(2)雌雄同体生物生态位分化不明显,或者未发生明显的生态位分化,或分化等级较低,都位于同一个体上,没有呈现雌雄异体生物的雌体与雄体生理与形态学上的诸多差异,其差异可能主要体现在雌雄性器官分化上,这也是雌雄同体生物一个显著的特点。雌、雄个体死亡率不同及其阶段性自然选择特点不同,这种驱动性比失调的生物学特性在雌雄同体中不存在,但主要作用在器官水平。器官水平性比是雌雄同体生物的基本特点,可与一般性结论进行类比。
(3)雌雄同体生物性比体现在两方面:一方面是雌、雄花(花序)比例(雌雄同株),另一方面是花粉胚珠比(雌雄同花)。这一突出了个体下等级组织结构自身对性比的决定性作用,强化了结构限制而弱化了环境决定的观点的影响,也增加了性比频度差异在不同环境中的异质性作用。简单例子,雌雄同株同化序瘠薄立地中单枝也能促成较高的性比,而肥沃生境中较高的枝条密度也能促成较小的性比形成。
(4)对于雌雄同体生物性比需要证明性比的遗传决定性,需要发现或创制突变体,这样才能套用雌雄异体生物性比的遗传决定的观点,或者说性比遗传决定的。若不存在这样的突变体,则雌雄同体生物性比就不能用遗传决定的观点来解释。突变体可能是性比遗传决定的材料,但实际上性别与性比的确受到基因或遗传控制,只是突变体没有呈现时认为是另种因素起主导作用而已。
(5)结构存在大小与年龄特异性,这一点在雌雄同体与异体生物之间寻找到共同点。即都存在大小依赖的性别决定特点,可用大小来解释所有生物性别决定的现象,性比都存在大小依赖性。这种生殖器官着生结构组织的大小现象对于解释性比作用明显,需要重视回归分析在性比研究中的作用。
(6)雌雄同花现象性比自身表现在花粉胚珠比,具有更强的结构依赖性,通常表现为明显的结构性比(发育上花或花序结构表现出来的)与功能性比(发挥适合度功能的雌雄比例),并且很多两性花都表现为单性功能而呈现两性花的结构。结构性比与功能性比需要开展观测与试验研究,主要考虑花粉、柱头寿命、花寿命、花开放式样、传粉者活动等,也受到时间、环境的影响。
(7)雌雄同体生物性比的环境决定的观点主要体现在群体水平上,需要借助性比或功能性状环境梯度分析的方法来研究,并且采用构件生长分析法,这样才能得到全面系统的分析信息。十一五期间麻疯树性比研究中就采用了种思路。
(8)雌雄同体研究需要注重两个尺度,一个是生态学等级的空间尺度(器官?构件?个体?群体?生态系统?景观?区域?全球);另一个是谱系进化的时间尺度(科属种发生的进化时期与性表达演化);后者的研究历史悠久,但案例还不够多,而前者则更缺乏。
(9)雌雄同体性比调控的试验生态学研究需要加强。雌雄异体植物性比环境决定在进行试验生态学研究时发现雌雄个体生物学特性差异太明显,这种资源利用能力差异还是生物学特性或遗传决定的,这就降低了该类群性比环境资源斑块性中环境决定性比假说解释的权威性,可能遗传因素与环境决定比重天平需要重新调整。需要对遗传决定性性比进行均一环境的检测,使得遗传特质得到相同环境诱导下的等量表达,从而突出遗传差异,从而检测性比差异的原因。
(l0)性比、生殖生态学与生态学综合研究日益重要。单纯性比研究不能解释性比的适应性问题,需要把性比与生殖,性比与生态学进行综合研究,并重视尺度,这样才能客观合理的认识性别、性比的生态适应性,而不是局限于一隅或窥豹一斑,片面认识是认识本身的绊脚石。
需要加强雌雄同体生物性比失调的研究,从野外调查与控制试验上全面探讨性比失调的决定机制,以满足对该类群性比失调研究结果的渴望。还需要强调空间因素和时间因素对雌雄同体性比的调控,如年际变化、立地类型、海拔经度、纬度等,并且这种时空调节还有生态学等级性,即各等级都存在明显相似的时空表现。并且,对雌雄异体生物而言,还需要补充很多新案例,并从试验角度探讨性比失调的决定机理:环境的,还是遗传的(证明)?我国这方面研究也很缺乏。突变体找寻也是研究性比遗传控制机制的基本材料,一方面通过筛选自然变异体,另一方面通过研究性别决定基因来创制突变体。性比失调研究最终需要揭示性表达之间演化上的相互关系,最终阐明自然界生物多种多样性表达进化的适应性问题,也是生态适应研究的入手点之一。同时,且矛盾具有时间与空间特性,这样因子时空作用范围与相互规律也能很快得到阐明,利于促进性比研究的迅速发展。
[l]Carwford R M M,Balfour J.Female predominant sex ratios and physiological differentiation in arctic willows.J of Ecology,l983,7l:l49~l60.
[2]Chardard D,Duumon C.Sex reversal by aromatase inhibitor treatment in the newt Pleurodeleswaltl.Journal of Experiment Zoology,l999,[[3]283:43 ~50.
[4]Dawson T E,Ehleringer JR.Gender-specific physiology,carbon isotope discrimination and habitat distribution in boxelder,Acer negundo.Ecology,l993,74:798 ~8l5.
[5]Dournon C,Guillet F,Boucher D,etal.Cytogenetic and genetic evidence of male sexual reversion by heat treatment in the newt Pleurtxleles pioireti.Chromosoma,l984,90:26l ~264.
[6]Ender R K.Rproduction in the mink.Proc.American Philosophy Society,l952,96(6):69l ~755.
[7]Grakov N N.Studying the sex and age structure of populations of the pinemarten.Biological Abs.,l963,45(l3):54402.
[8]Grant M C,Mitton JB.Elevational gradients in adult sex ratios of Populus tremuloides and sexual differentiation in vegetative growth rates Evolution,l979,33:9l4 ~9l8.
[9]Guo X G,Qian T J.Sex ratio and age struture of gamasiimites from smallmammals in western Yunnan,China.Entomologia sinica,200l,8(2):l66 ~l74.
[l0]HamoltonW J,Jr.Theweasels of new York.American Middle Nature,l933 ,l4:289 ~344.
[l l]Imamura T,Uraichuen J,Visarthanonth P et al.Effects of temperature on developemet of Theocolax elegans parasitizing larvae of themaize weevil Sitophilus zeamais in brown rice.Appl.Entomology of Zoology,2004,39(3):497 ~503.
[l2]Jacobsen R,et al.人类性比的自然变化.Hum Reprod(Natural variation in the human sex ratio),实用妇产科杂志,杨丹译.,l999.
[l3]Myers JH.Sex ratio adjustment under food stress:maximization of quality or number of offspring?The american naturalist,l978,l l2(984):38l~388.
[l4]Petrides G A.The determination of sex and age ratios in fur animals.American Nature,l950,43:355 ~382.
[l5]Venge O.Reproduction in fox and mink.Animal breed.Abs.,l959,27(2):l29 ~l45.
[l6]昌增印,杨建全,黄居昌,等.影响布氏潜蝇茧蜂子代性比的因素[J].福建林学院学报,2008,2l(3):225 ~228.
[l7]陈复生.过冷却处理家蚕卵对四倍体诱发率及后代性比的影响[J].蚕业科学,2002,28(4):294 ~297.
[l8]菊培,符悦冠.主要农林粉虱性比变化影响因素剖析[J].热带作物学报,2009,30(8):l l25 ~l l3l.
[l9]吴术德,曾惠民,刘建美,等.不同涌散期紫胶虫群体性比调查[J].林业实用技术,l988,(l2):30 ~3l.
[20]材路虹,石宝才,姚金亮,等.不同性比对小菜蛾繁殖及田间种群数量的影响[J].昆虫知识,20l0,47(l):63 ~66.
[2l]赤眼蜂寄生行为研究——雌蜂交配行为与子代性比[J].昆虫天敌,l992,l l(2):5l ~53.
[22]何丽芬,邱鸿贵,符文俊,等.影响松毛虫赤眼蜂性比的因素[J].昆虫天敌,l990,l2(2):66 ~70.
[23]候陵,刘文芳.温度对爬行动物性别的影响[J].生物学通报,l985,l l:l4 ~l5.
[24]胡萃,万兴生.蝶蛹金小蜂Pteromalus puparum性比的主要影晌因子[J].科技导报,l986,2(5):28 ~29.
[25]胡萃,叶恭银,李子川.蝶蛹金小蜂性比、寄生密度分布及两者关系的数学模拟[J].昆虫天敌,l993,l5(l):33 ~38.
[26]胡萃,叶慕银,王选民.野蚕黑卵蜂性比的研究[J].浙江农业大学学报,l993,l9(2):l2l ~l27.
[27]胡能书,朱泽瑞,吴秀山.红宝石激光微束照射黑腹果蝇(Drosophila melanogaster)的辐射生物学Ⅱ.对各发育阶段的时间、死亡率及性比的影响[J].湖南师范大学(自然科学学报),l987,l0(2):74 ~8l.
[28]霍雅鹏,万冬梅.鸟类性别鉴定的方法及性比研究的进展[J].辽宁大学学报(自然科学版),2008,35(4):358 ~362.
[29]金梅,崔玉颖,白庆余,等.中国林蛙性别控制及其生长发育的研究[J].两栖爬行动物学研究,2000,2000,8:l75 ~l80.
[30]康发功.甘肃兴隆山马麝种群年龄结构与性比初步研究[J].甘肃林业科技,2005,30(4):l2 ~22.
[3l]李金钢,王廷正.甘肃鼢鼠种群性比的研究[J].动物学研究,l999,20(6):43l ~434.
[32]李景科.奇变瓢虫的性比和雌、雄虫的区别[J].植物保护,l993,(5):5l.
[33]李璞.关于哺乳类的性比[J].动物学杂志,l958,2(l):l9~24.
[34]李璞.长期饥饿状态对鲫鱼(Carassius auratus)性比的影响[J].动物学报,l959a,l l(l):42 ~48.
[35]李璞.鲫鱼(Carassius auratus)性比的变异和选择性死亡[J].动物学报,l959b,l l(l):49 ~56.
[36]林益民.植物种群的性比[J].生态科学,l993,(2):l44~l48.
[37]刘冰,李昔,田启建,等.树干附生尖叶拟船叶鲜性比和有性生殖的比例[J].广西植物,2008,28(4):440 ~442.
[38]刘瑞林,王新省,傅鸣实,等.不同性比交配对玉米螟雌蛾生殖力的影响[J].山西大学学报,l983,93 ~96.
[39]刘相文,萧前柱.黑龙江省熊类年龄构成、性比及繁殖情况的初步研究[J].兽类学报,l986,6(3):l6l ~l70.
[40]罗礼溥,郭宪国.云南省25县(市)黄胸鼠体表革蜻性比及年龄构成分析[J].热带医学杂志,2006,6(3):293 ~295.
[4l]马立名.中国北方部分蚤类性比的研究[J].昆虫学报,l993,36(l):63~66.
[42]潘春芳,张春雨,赵秀海,等.不同林龄阔叶红松林林下簇毛械的性比格局及雌雄个体的空间分布[J].生物多样性,20l0,l8(3):292 ~299.
[43]秦启联,王金耀,徐世新,等.影响中红侧沟茧蜂后代性比的因素[J].中国生物防治,200l,l7(4):l55 ~l58.
[44]盛和林,陆厚基.黄鼬种群性比在狩猎生产上的意义[J].动物学报,l975,2l(4):344 ~349.
[45]宋波,彭艳琼,管俊明,等.西双版纳鸡嗦果榕内一种非传粉小蜂的性比调节[J].应用生态学报,2008,l9(3):588 ~592.
[46]宋波,彭艳琼,杨大荣.母体亲缘关系在杨氏榕树金小蜂后代性比调节中的作用[J].昆虫学报,2008,5l(4):4l l ~ 4l6.
[47]孙宝发,王瑞武,胡忠.榕小蜂的产卵模式及其对子代性比的影响[J].动物学研究,2009,30(5):559 ~564.
[48]孙永超,高燕,张中润,等.寄主及寄主食物对雅脊金小蜂子代性比体型和翅型的影响[J].昆虫知识,2009,46(5):749~755.
[49]汤玉清,徐清元.光周期对荔蜷卵平腹小蜂日产卵量及后代性比的影响.生物防治通报,l993,9(4):l48~l5l.
[50]王洪林.云南大理白族自治州人口的性比[J].人类学学报,l988,8:94 ~98.
[5l]王洪林.出生性比研究[J].人类学报,2000,l9(l):57 ~63.
[52]王金龙,魏万红,张堰铭,等.高原鼠兔种群的性比[J].兽类学报,2004,24(2):l77 ~l8l.
[53]王荣,陈时华.大磷虾性比、怀卵量与某些生殖特点的分析[J].南极研究,l989,l(3):6l ~68.
[54]王廷正,张越.白头叶猴的性比与社会结构[J].兽类学报,l995,l5(4):3l0.
[55]王问学.麦峨茧蜂性比与寄主的关系[J].生物防治通报,l99l,7(l):l6 ~l8.
[56]吴鹏飞,王跃招,王硕果,等.青海沙晰(晰蝎亚目:鬉晰科)种群的年龄结构与性比[J].四川大学学报(自然科学版),2002,39(6):l l34 ~l l39.
[57]肖绍军,张春兰,钟象景,等.性比对非繁殖期圈养白鸥行为的影响[J].安徽农业科学,2008,36(22):9559 ~9562.
[58]徐世多,谢国林,江建国.对影响马尾松毛虫幼虫性比因子的初步探讨[J].中国森林病虫,l983,4:9 ~l2.
[59]颜伟玉,涂剑锋.影响蜂王调节性比的营养因素[J].养蜂科技,2003,(5):38 ~40.
[60]杨伯然.长白山花尾棒鸡种群性比与年龄结构的初步分析[J].动物学报,l99l,37(3):334 ~335.
[6l]余荣,龙翔,耿利英,等.氯化镉对小鼠精液品质和仔鼠性比的影响[J].华中农业大学学报,2006,25(6):650 ~653.
[62]原建强,李欣.半闭弯尾姬蜂性比的影响因素研究[J].河南农业大学学报,2008,42(3):334 ~336.
[63]曾志将,陈国荣.蜜蜂性比的研究[J].蜜蜂杂志,l993,(4):4~5.
[64]张桂药.东方实蝇蛹俑小蜂性比、寿命和生殖力及低温冷藏对其影响的实验研究[J].生物防治通报,l992,8(l):l9~2l.
[65]张晓岚,孟玲,李保平.菜粉蝶蛹体型大小对蝶蛹金小蜂后代数量、性比及体型大小的影响[J].生态学杂志,2009,28(4):677~ 680.
[66]张子有,邵孟明,齐胜利.白蜡虫优良种虫及种群性比组成的研究[J].林业科学,l990,28(l):46 ~52.
[67]郑合勋.卢氏县大鲵种群年龄结构和性比的初步研究[J].动物杂志,2004,39(6):50 ~53.
[68]赵亚洲,辛雅芬,马钦彦,张春雨.雌雄异株树种黄连木种群性比及空间分布[J].生态学杂志,20l0,29(6):l087 ~l093.
[69]张春雨,王德胜,魏彦波.山杨、水曲柳种群密度与种群性比的关系[J].林业科学,20l0,46(7):l6 ~2l.
[70]Anne-Sofie Forfang,Jens M.Olesen.Male-biased sex ratio and promiscuous pollination in the dioecious island tree Laurusazorica(Lauraceae).Plant Systematics and Evolution,l998,3:l43 ~l57.
[7l]Makoto Kato,Hidetoshi Nagamasu.Dioecy in the endemic genus Dendrocacalia(Compositae)on the Bonin(Ogasawara)Islands.Journal of Plant Research,l995,4:443 ~450.
[72]John Paul Schmidt.Sex ratio and spatial pattern ofmales and females in the dioecious sandhill shrub,Ceratiola ericoidesericoides(Empetraceae)Michx.Plant Ecology,2008,2:28l ~288.
[73]Hu X S,Yeh FC,He FL.Sex-ratio distortion driven bymigration loads.Theoretical Population Biology,2007,72(4):547 ~559.
[74]Kiellberg F,Bronstein JL,Ginkel G V,et al.Clutch size:amajor sex ratio delerminant in fig pollinating wasps?Comptes Rendus Biologies,2005,328(5):47l ~476.
[75]Joyce A L,Millar JG,Paine TD,et al.The effect of host size on the sex ratio of syngaster lepidus,a parasitoid of Eucalyptus longhorned borers(Phoracanta spp.).Biological control,2002,24(3):207~2l3.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23 06:22:28
世界科学技术-中医药现代化(2021年5期)2021-11-05 06:56:18
中国科技纵横(2019年24期)2019-05-28 09:42:09
现代园艺(2018年2期)2018-03-15 08:00:10
安徽农业科学(2017年7期)2017-08-13 08:38:51
湖南中医药大学学报(2015年1期)2016-01-06 01:06:39
Coco薇(2015年12期)2015-12-10 02:40:50
山东农业科学(2014年12期)2015-03-10 11:33:38
中国医学科学院学报(2013年6期)2013-03-11 20:26:08
食品科学(2013年17期)2013-03-11 18:27:07
