
Go/NoGo实验下NoGo-N2、P3效应分析
2011-02-10 05:45:36刘铁军
电子科技大学学报 2011年3期
尹 刚,刘铁军
(电子科技大学神经信息教育部重点实验室 成都 610054)
在使用事件相关电位(ERP)研究反应抑制的脑机制时,通常采用Go/NoGo实验范式,即要求被试只对特定的靶刺激做出行为反应(Go任务),而对于非靶刺激,克制行为反应(NoGo任务)。在Go/NoGo实验范式下,很多研究者发现了与前额叶相关的抑制机制的ERP证据,其中两个最重要的ERP成份为:1) 在刺激后的大约200~300 ms内的N2成份,NoGo任务下的N 2相对于Go任务有更大的负向幅度;2) 在刺激后的大约300~600 ms内的P3成份,NoGo任务下P3相对于Go任务有更大的正向幅度。上述现象也称为NoGo-N2效应和NoGo-P3效应,被研究者认为是与反应抑制密切相关的[1-5]。
尽管很多研究结果表明在视觉或听觉Go/NoGo任务中能得到稳定的NoGo-N2、P3效应,然而在Go任务中由于有行为反应的参与,与行为反应有固定锁时关系的运动相关成份有可能与刺激引发的N2、P3成份产生相互交叠(刺激引发的N2、P3与刺激出现的时间有固定的锁时关系),造成真实的NoGo-N2、P3效应被Go任务中的运动相关成份所干扰。因而很多研究者对反应抑制过程中是否出现NoGo-N2、P3效应产生了争论,争论的焦点主要集中在NoGo-P3效应上[6-9]。文献[4]和文献[10]发现前额叶脑区的NoGo-P3效应仅出现在需要按键反应的实验条件下,并不出现在计数条件下。而文献[5]和文献[11]并没有得到类似结果。因此NoGo-P3效应是反映反应抑制的相关过程还是反映运动相关成份的干扰过程,依然是研究反应抑制脑机制中所争论的问题。
在Go任务下,刺激成份是锁时于刺激出现的时间,而运动相关成份锁时于反应时,因此直接以刺激时间为参考的叠加平均ERP中既包含刺激成份又包含运动相关成份,从而掩盖了Go任务下刺激引发的真实ERP成份。基于该问题,文献[12]提出了刺激、反应成份分解方法,能有效地恢复Go任务下真实的刺激、反应成份,该算法已经推广到多事件相关成份的分解[13]。
本文首先运用刺激、反应成份分解方法恢复Go任务下刺激引发真实的ERPs成份,然后对比Go/NoGo两种任务下N2、P3成份之间的差异,对NoGo-N2、P3效应做进一步的讨论。
1 刺激、反应成份分解方法

2 实验数据
本文中使用的数据来自网上免费数据(http://www.sccn.ucsd.edu/~arno/fam2data/publicly_a vailable_EEG_data.htm l)
2.1 实验设计
14个被试者参与动物分类和识别任务两种任务条件,本文采用分类动物与非动物图片的实验数据。靶刺激图片和非靶刺激图片以等概率(50%)的方式呈现。实验采用经典Go/NoGo范式,行为反应的方式是抬起压在触敏按钮上的手指。对于Go任务,要求被试者看到靶刺激图片时尽可能快和准确地做出行为反应。
实验开始时,黑色背景屏幕中央出现0.1°视角的固定点,刺激图片为宽4.5°视角、高6.5°视角,呈现时间为20 ms,要求被试在1 000 ms内做出行为反应。刺激呈现的时间间隔为1 800~2 200 ms。
2.2 数据获取
EEG数据记录采用32导脑电采集系统(Oxford Instruments),以头顶电极(CZ)作为参考电极。EEG的采样率为1 000 Hz,并对EEG数据做30 Hz的低通滤波。在迭加平均脑电数据之前需要去除伪迹(眨眼、眼动以及肌电),参考电极被重新计算为平均参考。
3 结 果
本文分别对所有14个被试者计算Go任务下的刺激、反应成份分解的结果,得到Go任务下刺激引发的纯刺激成份。本文选择N2成份的时间范围在刺激后200~300 ms,图中的阴影区分别表示N2、P3时间段,P3的时间范围在刺激后的300~500 ms。其中Go条件的ERP波形有两种,一种是直接叠加平均ERP波形(Go直接平均),另一种是通过刺激反应成份分解恢复出的纯刺激成份ERP波形(Go恢复)。
图1显示了Go条件下纯刺激成份ERP和直接迭加平均ERP以及NoGo条件下的平均ERP波形。为了说明方便,将Go任务下分解后得到的纯刺激成份ERP称为Go恢复,将Go任务下直接刺激时间叠加平均ERP称为Go直接平均。在比较两种任务下的N2和P3成份时,选择刺激后200~300 ms和刺激后300~500 ms时间段内ERP的平均幅度分别作为N2和P3成份的幅度。
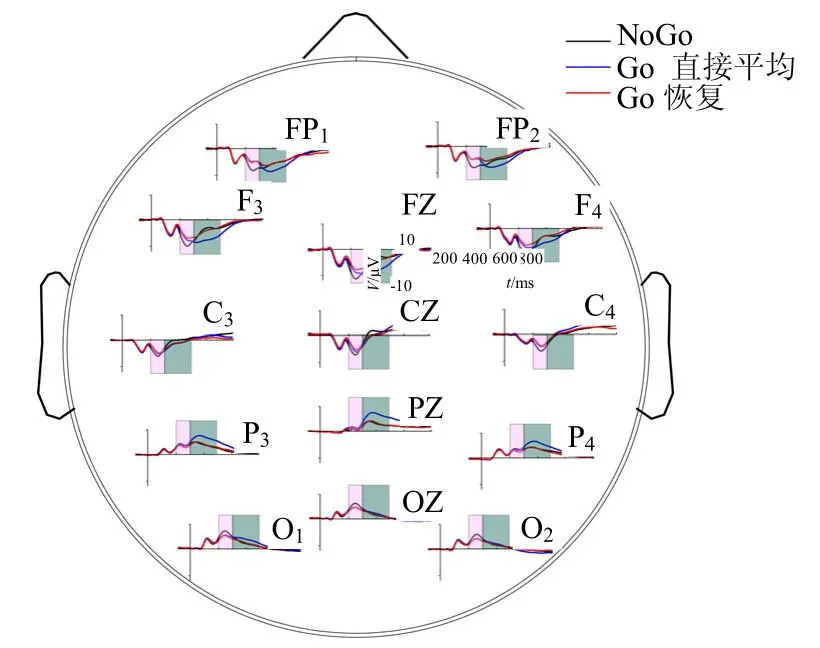
图1 14个被试者分别在Go和NoGo条件下的总平均ERP波形
3.1 N2成份
图2a显示了不同脑区N2时间段内的平均幅度,本文选择了不同的脑区的3个典型电极:FZ(前额叶区)、PZ(后顶区)、OZ(枕区)。对于FZ电极,配对T检验的统计结果显示NoGo任务下的N2相对于Go任务有更大的幅度(NoGovs.Go直接平均t=6.616,p<0.001;NoGovs.Go恢复t=5.845,p<0.001)。同时Go直接平均的N2幅度大于Go恢复的N2幅度(Go直接平均vs.Go恢复t=2.185,p=0.0478<0.05)。对于Pz电极,两种任务下的N2幅度没有显著性差异(NoGovs.Go直接平均t=1.036,p=0.319>0.05;NoGovs.Go恢复t=0.233,p=0.820>0.05)。对于OZ电极,NoGo任务相对于Go任务有更大的幅度(NoGovs.Go直接平均t=5.527,p<0.001;NoGovs.Go恢复t=4.428,p<0.001)。同时Go直接平均幅度和Go恢复的N2幅度没有显著性差异(Go直接平均vs.Go恢复t=0.85,p=0.41>0.05)。

图2 3个典型电极(FZ、PZ、OZ)上的N2成份和P3成份幅度的对比
3.2 P3成份
对于刺激后300~500 ms的时间段,计算3种情况下(NoGo、Go直接平均、Go恢复)ERPs的平均幅度并做配对T检验,结果如图2b所示。对于FZ电极,Go直接平均的P3相对于NoGo任务有更大的幅度(NoGo vs. Go直接平均t=4.262,p<0.001),Go恢复的P3幅度和NoGo任务下P3幅度无统计显著性差异(NoGo vs. Go恢复t=0.443,p>0.05)。同时,Go直接平均P3幅度和Go恢复P3的幅度有显著性差异(Go直接平均 vs.Go恢复t=4.588,p<0.001)。对于PZ电极,其结果和Fz电极上的结果类似,Go直接平均的P3幅度相对于NoGo和Go恢复均有更大的幅度,Go恢复的P3幅度和NoGo任务P3幅度无统计显著性差异(NoGovs.Go直接平均t=4.509,p<0.001;NoGo vs. Go恢复t=0.365,p=0.721>0.05;Go直接平均vs.Go 恢复t=6.552,p<0.001)。对于OZ电极,各情况之间均无统计显著性差异(N o G o v s.G o直接平均t=1.668 1.95,p=0.119>0.05;NoGo vs.Go恢复t=1.95,p=0.079>0.05;Go直接平均vs.Go恢复t=1.711,p=0.09>0.05)。
3.3 NoGo、Go差异成份
为了检测Go、NoGo任务之间ERP成份的差异,通常采用的方法是对两种任务下的ERP波形进行配对相减得到差异波。图3显示了NoGo和Go任务间的ERP差异波以及差异波在N2和P3时间段内的平均头表地形图,图中的阴影部分分别表示N2(200~300 ms)和P3(300~500 ms)时间段。
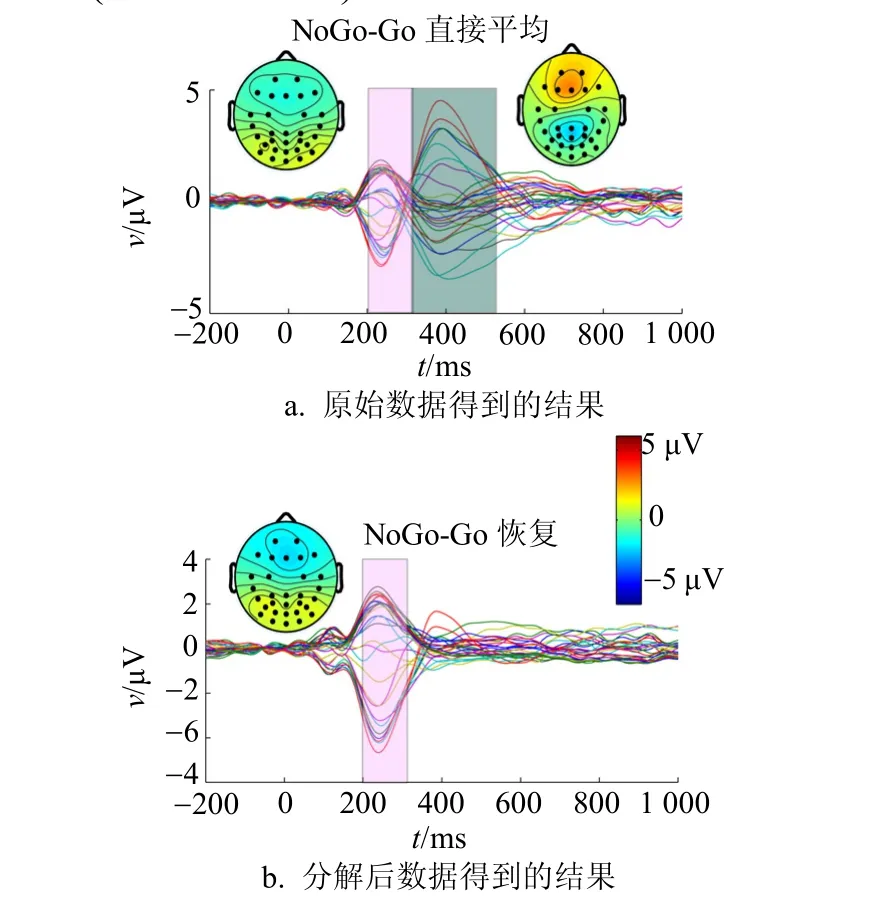
图3 NoGo和Go两种任务之间的ERP差异波以及差异波在N2和P3时间段的平均头表地形图
4 讨 论
传统的研究表明,Go/NoGo实验范式下,大脑前额叶区NoGo任务相对于Go任务有N2的负向偏移和P3的正向偏移(NoGo-N2、P3效应),因此文献[1-5]认为NoGo-N2、P3效应反映了反应抑制的过程。本文采用直接迭加平均ERP的对比结果表明NoGo-N2、P3效应和传统的研究结果是一致的。然而,关于NoGo-N2、P3效应是否反映反应抑制过程依然存在一些争论,争论的焦点是NoGo-N2、P3效应是否来自于Go任务下的运动相关成份。研究结果表明,从Go任务下直接叠加平均ERP中剥离了运动相关成份后NoGo-N2效应依然存在,而NoGo-P3效应则消失了,如图3b所示,这说明在直接叠加平均下得到的NoGo-P3效应是运动相关成份的叠加效果,很可能与反应抑制过程无关。下面分别讨论这两个效应。
4.1 Go/NoGo的N2效应
对于刺激后的大约200~300 ms前额区的N2成份,NoGo任务相对于Go任务(Go直接平均,Go恢复)有一个负向偏移。虽然对比NoGo和Go直接平均以及NoGo和Go恢复都得到似的NoGo-N2效应,但对比NoGo和Go恢复得到的NoGo-N2效应更为明显,同时图3中NoGo-N2效应的头表地形图表现在前额区更为聚集,这主要是因为在前额叶区靠近运动区,Go任务下运动相关电位的早期负向电位通过体传导叠加到刺激引发的ERP上,使得刺激引发的N2成份幅度增加,从而导致NoGo和Go任务之间的N2成份幅度差异减小。因此本文的结果增强了NoGo-N2效应。
4.2 Go/NoGo的P3效应
在Go/NoGo实验范式下,传统的研究结果表明在刺激后的大约300~500 ms前额叶区NoGo任务的ERP相对于Go任务的ERP有正向偏移,即NoGo-P3效应。对比直接叠加平均ERP,能得到很明显的NoGo-P3效应(如图2b和图3a所示)。同时NoGo-P3效应有很明显的前额叶区的分布(图3a),这与传统的研究结果相符合。当从Go任务下直接叠加平均ERP中剥离开运动相关成份以后,无论时间过程还是头表地形图NoGo-P3效应都消失了,这说明按传统方式得到的NoGo-P3效应的确来自运动相关成份的叠加效果。本文的研究结果也证实了文献[6]和文献[14]的推测:NoGo-P3效应并不反映在NoGo条件下的抑制过程,可能仅仅是Go条件下运动相关成份的叠加效果。在文献[15]的研究结果中,计数条件下NoGo任务并没有相对于Go任务下更大的P3幅度,特别是在前额区(作者并未对此现象做出讨论),这也从另外一个角度说明了在按键条件下运动相关成份对NoGo-P3效应有很大的影响。但文献[11]在对比按键和计数条件下的NoGo-P3效应时发现两种情况都观察到了前额区NoGo任务下P3的幅度大于Go任务下P3的幅度,这与文献[6]的研究结果相矛盾,同时文献[5]的研究结果也得出类似结论:按键和计数两种情况都观察到了前额区NoGo-P3效应,而计数条件下的NoGo-P3效应比按键条件弱,其原因是NoGo-P3效应有少部分是来自于Go任务下的运动相关成份。实际上造成上述研究结果的不一致以及对NoGo-P3效应解释的分歧主要有以下3个可能的因素:
1) 刺激概率,无论是Go任务还是NoGo任务下的P3幅度都和刺激概率成负相关的关系[15],从文献[15]的研究结果可以看出,当刺激概率为75%时NoGo-P3效应基本上消失了。文献[6]采用的靶刺激概率为15%,文献[5]采用的靶刺激概率为20%,文献[11]采用的是等概率50%刺激。
2) Go任务下反应时的差别。反应时的长短与实验任务的难度、刺激方式以及被试的精神状态等因素有关[16]。反应时长短以及反应时的分布,都将影响运动相关成份在刺激引发的ERP上的叠加效果。文献[6]的实验中,对于不同的刺激概率平均反应时是不一样的(25%:434 ms;50%:401 ms;75%:373 ms),文献[10]使用体感刺激时得到的平均反应时是216.7 ms,文献[5]使用听觉刺激得到的平均反应时分别是434和535 ms。可以看出不同实验条件下得到的反应时差别是比较大的。
3) 在计数条件下被试的计数策略的差别。在按键条件下要求被试尽可能最快和最准确地做出行为反应。和按键反应不一样,计数条件下被试没有时间压力,因此被试完成任务的策略也不一定是相同的,例如被试可以在靶刺激出现后立即计数,也可以等到下一个刺激来临之前计数。很明显这两种计数方式的心理过程是不一样的,因此会导致最终得到ERP波形的不一致。另外由于计数方式没有外部行为表现,在实际实验过程中不可能监测到被试的计数策略,因此不同的计数策略下得到的NoGo效应也可能是有差别的。
综上所述,出现对NoGo-P3效应解释的广泛争论是因为缺乏运动相关成份是否影响NoGo-P3效应最直接的证据。因此,只有从Go任务中恢复出真实的刺激引发的成份才能找到NoGo-P3效应是否反映反应抑制过程的直接证据。
另外,由于本文的实验中靶和非靶刺激的概率都是50%,被试在实验任务中对靶和非靶刺激投入的心理资源量是相等的,本文的研究从Go任务下直接叠加平均ERP中剥离运动成份后,Go任务和NoGo任务在后顶区的P3成份也没有差别如(图1和图3),该结果符合相关文献报道:经典P3主要分布在后顶区,它的幅度和刺激概率成负相关的关系,且P3的波幅与投入心理资源量成正比[17]。而直接迭加平均的结果显示大脑后顶区的Go任务下的P3幅度比NoGo任务下的P3幅度大,这说明运动相关成份的叠加效果在后顶区依然存在。因此本文实验采用刺激反应成份分解方法能有效地去除经典叠加平均方法中运动相关成份的干扰。
5 结 论
通过刺激、反应成份分解算法从Go任务下ERP数据中恢复出真实的刺激引发的ERP成份,结果表明,NoGo-N2效应反映了反应抑制过程,而NoGo-P3效应是来自运动相关成份的干扰,与反应抑制过程无关。本文的结果是在等概率视觉刺激实验条件下得到的,对于不同的刺激概率以及不同的刺激方式(听觉,体感等)是否有相同的结论还需要进一步的研究论证。
[1] KOK A. Effects of degradation of visual stimulation on components of the event-related potential (ERP) in go/nogo reaction tasks[J]. Biol Psychol, 1986, 23(1): 21-38.
[2] JODO E, K AYAMA Y. Relation of a negative ERP component to response inhibition in a Go/No-go task[J].Electroencephalography and Clinical Neurophysiology,1992, 82(6): 477-482.
[3] BEKKER E M, KENEMANS J L, VERBATEN M N.Electrophysiological correlates of attention, inhibition,sensitivity and bias in a continuous performance task[J].Clin Neurophysiol, 2004, 115(9): 2001-2013.
[4] SM ITH J L, JOHNSTONE S J, BARRY R J. Effects of pre-stimulus processing on subsequent events in a warned Go/NoGo paradigm: response preparation, execution and inhibition[J]. Int J Psychophysiol, 2006, 61(2): 121-133.
[5] SM ITH J L, JOHNSTONE S J, BARRY R J.Movement-related potentials in the Go/NoGo task : The P3 reflects both cognitive and motor inhibition[J]. Clini Neurophysiol, 2008, 119(3): 704-714.
[6] SALISBURY D F, GRIGGS C B. The NoGo P300‘anteriorization’ effect and response inhibition[J]. Clini Neurophysiol, 2004, 115(7): 1550-1558.
[7] BEKKER E M, KENEMANS J L, KOEKSMA M R, et al.The pure electrophysiology of stopping[J]. Int J Psychophysiol, 2005, 55(2): 191-198.
[8] DIMOSKA A, JOHNSTONE S J, BARRY R J. The auditory-evoked N2 and P3 components in the stop-signal task: indices of inhibition, responseconflict or error-detection?[J]. Brain Cogn, 2006, 62(2): 98-112.
[9] SM ITH J L, JOHNSTONE S J, BARRY R J. Response priming in the Go/NoGo task: the N2 reflects neither inhibition nor conflict[J]. Clin Neurophysiol, 2007, 118(2):343-355.
[10] HIROKI N, KOJI I, YOSHIAKI N, et al. Effects of a go/nogo task on event-related potentials follow ing somatosensory stimulation[J]. Clini Neurophysiol, 2004,115(2): 361-368.
[11] PFEFFERBAUM A, FORD J M, WELLER B J, et al.ERPs to response production and inhibition[J].Electroenceph Clin Neurophysiol, 1985, 60(5): 423-434.
[12] ZHANG J. Decomposing stimulus and response component waveforms in ERP[J]. Journal of Neuroscience Methods, 1998, 80(1): 49-63.
[13] YIN G, ZHANG J, TIAN Y, et al. A multi-component decomposition algorithm for event-related potentials[J].Journal of Neuroscience Methods, 2009,178(1): 219-227.
[14] CHEN C Y, TIEN Y M, JUAN C H, et al. Neural correlates of impulsive-violent behavior: an event-related potential study[J]. Neuroreport, 2005, 16(11): 1213-1216.
[15] BRUIN K J, WIJERS A A. Inhibition, response mode, and stimulus probability: a comparative event-related potential study[J]. Clin Neurophysiol, 2002, 113(7): 1172-1182.
[16] ROBERT J. KOSINSKI. A literature review on reaction time[J/OL]. [2009-08-11]. http://biae.clemson.edu/bpc/bp/Lab/110/ reaction.htm#Type%20of%20Stimulus.
[17] 魏景汉, 罗跃嘉. 认知事件相关脑电位教程[M]. 北京:经济日报出版社, 2002.
WEI Jing-han, LUO Yue-jia. The cognitive ERP textbook[M]. Beijing: Economic Daily Publishing Company, 2002.
编 辑 黄 莘
猜你喜欢
养生月刊(2020年10期)2020-10-14 02:06:26
宇航计测技术(2018年3期)2018-09-08 02:21:24
证券市场红周刊(2018年41期)2018-05-14 18:45:56
证券市场红周刊(2018年22期)2018-05-14 17:40:17
Transactions of Nanjing University of Aeronautics and Astronautics(2018年1期)2018-03-29 07:35:47
制造业自动化(2017年2期)2017-03-20 14:26:17
安徽医科大学学报(2016年12期)2017-01-15 14:21:49
中国洗涤用品工业(2015年9期)2015-02-28 19:03:05
股市动态分析(2014年27期)2014-07-29 19:40:43
体育教学(2012年10期)2012-04-29 13:44:44
