
GDGT在全球气候变化研究中的应用进展*
2011-01-10 09:32于志刚赵美训
中国海洋大学学报(自然科学版) 2011年5期
姚 鹏,于志刚,赵美训
(1.中国海洋大学化学化工学院,海洋化学理论与工程技术教育部重点实验室,山东青岛266100;2.中国海洋大学化学化工学院,海洋有机地球化学研究所,山东青岛266100;3.英国约克大学化学系,约克YO10 5DD)
甘油双烷基甘油四醚(Glycerol Dialkyl Glycerol Tetraethers,GDGTs)是微生物细胞膜脂的主要成分[1]。从结构上来说,GDGTs可以分为两大类,即类异戊二烯GDGTs(Isop renoid GDGTs)和支链GDGTs(Branched GDGTs)(见图1)。类异戊二烯GDGTs分子中包含数目不等的环戊烷结构(见图1,Ⅰ~Ⅴ)[2]。其中泉古菌醇(Crenarchaeol,图1-Ⅵ)及其重构异构体(Regioisomer,见图1-Ⅵ′)是一类特殊的类异戊二烯GDGTs,分子中除了含有4个环戊烷结构外,还含有1个环己烷结构[3]。支链GDGTs则一般含有2~6个甲基支链和1~2个环戊烷结构[4](见图1,Ⅰa~Ⅲc)。
近几年来,基于这两类GDGTs分子比值的各种指标在海洋和陆地古温度重建等方面显示了巨大的应用潜力,已逐渐成为古海洋学、古湖沼学和古气候学研究的重要工具[1]。类异戊二烯GDGTs主要来自古菌门(A rchaea)的泉古菌集合Ⅰ(Group I Crenarchaeota)[3]。研究表明类异戊二烯GDGTs分子中的环结构数目和温度相关,根据不同结构的类异戊二烯GDGTs的相对分布所建立的TEX86(TetraEther indeX of 86 carbon atom s)古温度指标可以用于重建表层海水温度(Sea Surface Temperature,SST)[5]。支链GDGTs主要来自土壤中的厌氧细菌[4],在土壤侵蚀和河流搬运的作用下进入近海沉积物中[6]。根据代表海洋环境的泉古菌醇与代表陆地环境的支链GDGTs的相对比值建立的支链类异戊二烯四醚指标(Branched Isoprenoid Tetraether,B IT)可以用来定量估算陆源和海源有机质的相对丰度[6]。进一步的研究表明,支链GDGTs分子中环戊烷结构的数目与土壤p H相关,而甲基数目与平均大气温度(Mean Air Temperature,MA T)和土壤p H都相关[7]。根据不同结构的支链GDGTs的相对分布分别建立了反映土壤p H的CBT(Cyclization index of Branched Tetraethers)指标和反映平均大气温度及土壤p H的MBT(M ethylation index of Branched Tetraethers)指标,这2个指标与平均大气温度之间的函数关系可以用来重建陆地温度、土壤p H和海拔历史等[8-9]。本文从类异戊二烯GDGTs和支链GDGTs2个方面介绍与GDGTs相关的最新研究进展。
1 类异戊二烯GDGTs
泉古菌在开阔大洋中分布广泛,即使是在低生产力的区域[11],据估计它们可能占海洋超微型浮游生物(Picop lankton)生物量的20%[12]。泉古菌的细胞膜是由类异戊二烯GDGTs作为核心脂、以磷酸和糖苷等作为极性头基而形成的两面都是亲水基的单层膜脂,而真核生物和细菌的细胞膜脂一般是双层结构,并且类异戊二烯GDGTs分子中环戊烷结构数量的增加会使膜脂组装得更密实,这有效地避免了双层膜在高温下变性分开,并且保持了完整的内层疏水结构,使泉古菌适于在高温(60℃以上的热泉或热液喷口)和低p H等极端环境条件下生存[13-14]。越来越多的研究表明,在海洋、湖泊、土壤和泥炭中类异戊二烯GDGTs都广泛存在,说明无论是嗜热(Thermophilic)还是非嗜热(Nonthermophilic)古菌都能生产类异戊二烯GDGTs[3-15]。
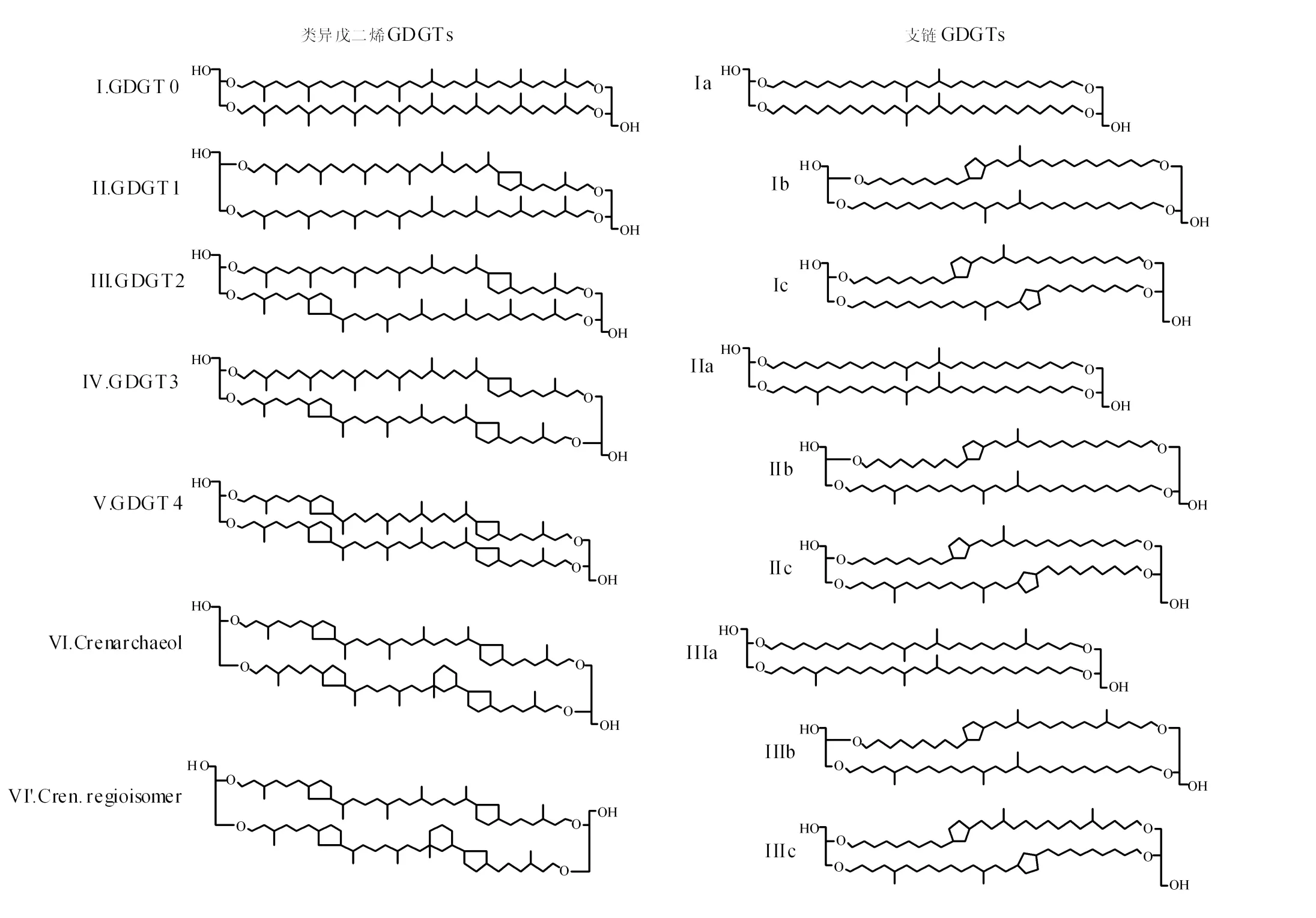
图1 GDGTs的分子结构式[10]Fig.1 Structures of GDGTs[10]
尽管醚类化合物稳定性稍次于烷烃,但其化学活性低,因此类异戊二烯GDGTs可在沉积物中较好的保存下来,即使是在1.12亿年前白垩纪的沉积物中也能检测的到[16]。对海洋表层沉积物中的类异戊二烯的GDGTs研究表明,GDGTs分子中环戊烷结构的数量与表层海水温度有很好的相关性,因而可应用描述这些GDGTs相对丰度的指标(TEX86)进一步定量描述年平均表层海水温度(SST)[5]。
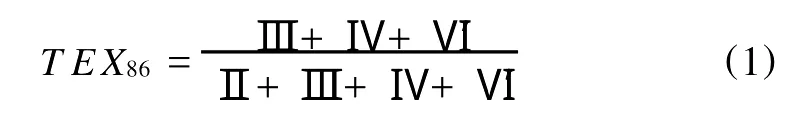
(罗马数字代表的GDGTs结构见图1)
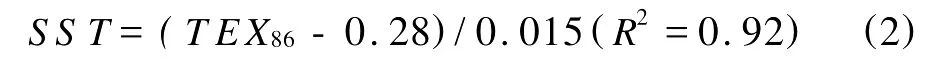
(该方程的适用温度范围为0~30℃)
TEX86提出之后,不同的研究者开展了多项工作验证其可靠性和适应性[1,17],并根据适用条件提出了多个校正方程[18]。W uchte等开展了围隔实验,通过分析不同温度和盐度条件下的泉古菌的类异戊二烯GDGTs,计算相应的TEX86[19],结果表明,TEX86的变化与盐度和营养盐无显著相关性,而是随着温度变化而发生显著变化,证实海洋古菌确实会根据温度调节类异戊二烯GDGTs的分布。Trommer等在高盐的红海的研究结果也证实盐度对TEX86的影响不大[20]。需要指出的是,p H也可能影响类异戊二烯GDGTs的分布,但是p H在海洋系统中的变化通常很小,所以其影响可以忽略[21]。
TEX86指标的特点之一是可以应用于高温区域和历史上高温期的温度重建。Schouten等把采集自热带印度洋海水中的海洋泉古菌在25~40℃条件下进行培养,结果显示泉古菌在40℃的高温环境中仍然生长良好,且TEX86在这个范围内都和SST线性相关,说明其可以用于地质历史高温时期的SST重建[15]。这比常用的只适合SST低于29℃的指标具有明显的优势[18]。在缺乏烯酮、碳酸岩和有孔虫的沉积区域和更加古老的地层中(早至中生代白垩纪),TEX86也得到了成功应用[16,22]。不过,TEX86在海洋系统的应用也存在一些限制[18,23],比如不能用于含高成熟度有机质的沉积物[24],不适用于类似于现代两极地区较冷水体和缺氧的富甲烷环境的古SST重建等[17]。目前TEX86的应用多集中于开阔海域,在边缘海的应用较少。研究的初步结果表明TEX86在我国陆架海可以应用,有可能弥补U37K指标在水深30 m之内基本不能用的不足(赵美训等,未发表数据)。另外,有研究指出TEX86指标除指示温度外,在反映古菌生态、营养盐含量和水文条件等方面也有很大潜力,但尚需更深入的研究论证[25]。
目前用于重建湖泊环境古温度的指标比较稀少,代表性的有硅藻[26]和摇蚊化石[27]沉积记录、碳酸盐氧同位素[28]、长链不饱和烯酮[29-31]等,这些指标大部分都显著地受到在湖泊环境中变化大的盐度和p H等参数的影响[1]。TEX86指标在海洋中的成功应用促使人们思考它是否也能应用于湖泊环境,即能否作为1个独立的重建陆地古温度的新方法。对不同气候类型的湖泊的研究表明,所有的沉积物中都含有大量的泉古菌GDGTs[7,32]。Pow ers研究了15个分布于全球各地的湖泊的TEX86指标,发现其与年平均湖泊表面温度(lake surface temperature,LST)之间存在线性关系,而且其斜率也与来自海洋的相似,表明这一古温度指标确实可以应用在湖泊环境中[33]。Powers等还由此重建了东非Malaw i湖末次冰盛期(Last Glacial Maximum)以来的年平均湖面温度,发现从末次冰盛期到现在年平均湖面水温升高了约3.5℃,在新仙女木事件(Younger Dryas,12.5 ka BP)期间和8.2 ka BP时则有约2℃的下降[34]。Tierney等运用TEX86指标研究表明,东非Tanganyika湖在全新世的年平均湖面温度大致在27~29℃之间变化,而在末次冰盛期时其温度则要低约5℃,这一结果和Powers等的结果相当吻合[35]。两项研究都表明,TEX86是一个有前途的研究陆地古气候的工具[1]。不过,随后有关47个欧洲较小湖泊的研究发现,TEX86在湖泊中的应用还存在一些困难,主要是附近流域土壤中的产甲烷古菌(Methanogenic)、甲烷氧化古菌(Methanotrophic)和嗜热的古菌也产生用于计算TEX86的GDGTs,它们的输入可能使TEX86指标不能准确反映湖泊温度[36],今后的工作需要考虑确定这些输入对TEX86指标反演湖泊温度影响程度以及如何消除这些输入的影响[37]。
我国湖泊众多,气候类型多样,许多湖泊由于所处的地理位置特殊,使它们成为地质历史时期区域气候、植被以及人类活动演化的良好载体。比如我国内陆最大的咸水湖——青海湖位于青藏高原东北部,处于对环境变化敏感的西部干旱区与东亚季风湿润区的过渡地带,受冬、夏季风和西风环流的影响,在全球气候变化研究中具有重要地位[38]。在湖泊表面温度重建方面,长链不饱和烯酮,即U37K指标的应用较多。Li等人曾在青海湖表层沉积物中检出了长链不饱和烯酮,进而提出了经过校正的U37K指标用于湖泊水体古温度的重建,但同时Li指出,除了温度之外,长链不饱和烯酮的不饱和类型还可能受到生物来源、盐度等诸多不确定因素变化的影响,进而影响温度估算结果的准确性[39]。付明义等研究了我国青海湖和柴达木盆地地区不同类型湖泊表层沉积物中的长链不饱和烯酮,并估算了两地区湖泊温度,发现估计的温度在青海湖地区(咸水湖)处于实际温度范围内,而在柴达木盆地(盐湖)低于实际温度,同样说明盐度对U37K指标的结果有较大的影响[40]。作为一种新的且已在国际研究中取得突出进展的湖泊表面温度替代指标,TEX86在我国湖泊领域的应用值得期待。
2 支链GDGTs
支链GDGTs的发现比类异戊二烯GDGTs的发现要早。2000年在德国的一个全新世泥炭沉积物中,Sinninghe Dam sté等发现了支链GDGTs并使用核磁共振确定其结构[41],随后的研究表明它们在土壤中也广泛存在[42]。支链GDGTs在结构上和类异戊二烯GDGTs不同,它们的碳骨架不是类异戊二烯结构,而是具有数目不等的甲基支链,环戊烷结构的数目(0~2个)也比类异戊二烯GDGTs少,而且没有环己烷结构(见图1)[4]。对支链GDGTs的甘油结构的立体结构分析表明,它们和细菌合成的甘油结构具有同一立体构型[4]。在嗜热细菌中还曾经发现支链双烷基甘油二醚(Dialkyl Glycerol Diethers)[43-45]。上述研究表明支链GDGTs可能是由细菌生产的,但是这些细菌的准确系统发育地位尚不明确[4]。
在一些近岸海洋沉积物中也发现了支链GDGTs,这可能是陆地土壤有机质通过河流输送所致[6,10,42,46-49]。支链GDGTs在近海沉积物中的丰度可被用来定量指示土壤有机质向海洋系统的输入,沉积物中支链GDGTs的含量与主要来自海洋古菌的类异戊二烯GDGTs泉古菌醇的含量之比称为支链和类异戊二烯四醚指标(Branched and Isop renoid Tetraether,B IT)[6]:
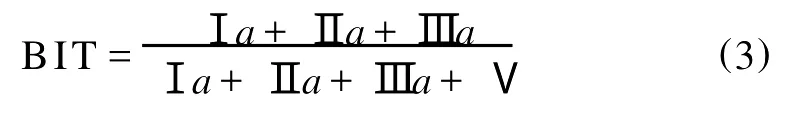
(罗马数字代表的GDGTs结构见图1)
BIT=1代表沉积物全部来自土壤有机质,而BIT=0代表有机质全部来自海洋。
对来自不同环境的大量样品的分析显示,B IT指标在海洋悬浮颗粒物和沉积物研究中有较大的应用潜力,所以BIT起初被认为可以用来定量估算近岸海洋环境中陆源有机质的来源和分布特征等[6,47-51]。但是,Walsh等利用δ13CTOC、木质素和B IT 3个参数研究了美国华盛顿州-加拿大温哥华外的边缘海沉积物有机物来源发现,B IT指标不能很好地反映陆源有机物输入,只能反映土壤和泥炭有机质的输入[49]。所以在古环境研究中B IT指标被用来定性追踪土壤有机质输入的历史变化[52-53],但如果结合传统指标还可以进一步区分沉积有机质来源。例如,Weijers等建立了基于δ13CTOC、C/N比和B IT指标的三端元模型,首先区分陆源和海源有机质的输入,然后将陆源有机质分成土壤和植物来源2部分,对土壤有机质进行了定量研究[54]。结果显示在过去的20 ka,土壤有机碳占了刚果河深海扇形地(Congo deep-sea fan)沉积物中有机碳的近一半(平均约45%),更占了陆源有机质输入的大部分;土壤有机碳的积累率从约17 ka BP开始增加,在新仙女木事件时期有所降低,峰值出现在早全新世,而低值在晚全新世,这一变化模式与中部非洲湿度和刚果河流量的历史变化相类似,表明中部非洲降雨变化规律主导了陆源有机碳在刚果河深海扇形地的沉积变化规律[54]。而Walsh等的研究却发现美国华盛顿洲-加拿大温哥华外的边缘海沉积物中的陆源有机质主要来自植被,土壤和泥炭有机质的贡献很低[49]。有研究提出,为了消除陆源物质的影响,可以将TEX86和B IT指标同时测定,从而将TEX86用于近海古温度重建[18],Rueda等的研究结果支持了这一设想[10]。另外,Blaga等对湖泊样品同时测定了BIT指标及GDGT-0/Crenarchaeol比值,发现对于BIT<0.4和GDGT-0/Crenarchaeol比值<2的9个湖泊,TEX86指标可以用来重建湖泊古温度[36]。所以,进行多参数测定并进行分析比较,是应用TEX86指标准确估算近海、湖泊古温度所必备的。
Weijers等分析了来自全球130个土壤样品的支链GDGTs,发现支链GDGTs在土壤中的相对分布主要受大气温度和土壤p H2个因素控制[55]。支链GDGTs分子中环戊烷结构和土壤p H相关,而甲基数目主要受土壤p H和平均气温(MA T)影响。根据这些关系,提出了2个指标定量描述支链GDGTs分布的变化。1个是反映土壤p H的CBT(Cyclization index of B ranched Tetraethers):
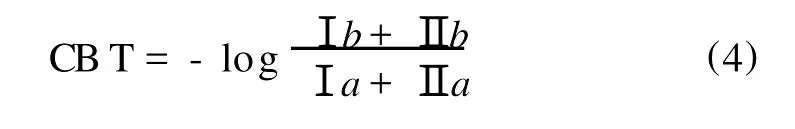
(罗马数字指示的结构请见图1)
并得出经验公式:
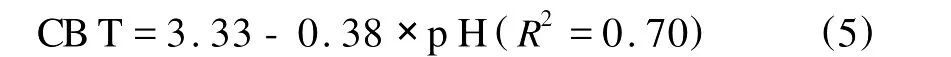
由此可见当p H上升时,CBT指标下降,表示GDGT分子系列中环戊烷数量增加。
另一个是反映大气温度和土壤p H的MBT(Methylation index of Branched Tetraethers):
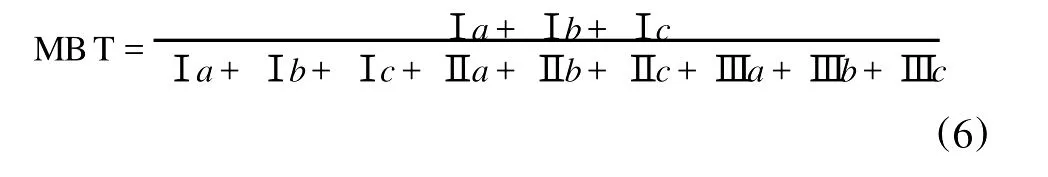
并得出经验公式:
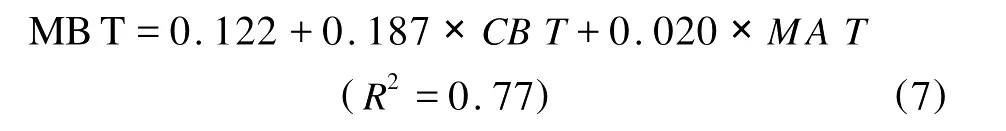
其中MA T是平均大气温度。由此可见当温度上升时,MBT指标增加,表示GDGT分子系列中甲基数减少。通过比较公式(5)和(7)可以进一步得出当p H上升时,MBT指标下降,表示GDGT分子系列中甲基数增加。
将公式(7)重新组合,即可在测定支链GDGTs分布的基础上重建陆地古温度:
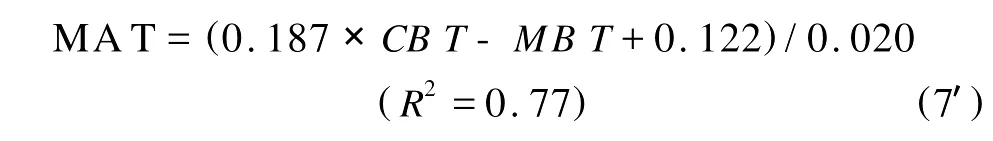
MBT/CBT指标已经在不同区域和地质年龄的海洋沉积物中得到应用[10,55-57]。在长时间尺度上,始新世-渐新世边界(Eocene-Oligocene boundary)是地球历史上气候转变的关键阶段之一,见证了东南极冰盖的首次重大扩张,但是对于气候变冷的程度,尤其是北半球高纬度内陆地区,却了解的很少[58]。Schouten等分析了采自格林兰盆地(Greenland Basin)的沉积物柱状样中的支链GDGTs,重建了格林兰大陆始新世末至渐新世早期的MA T,发现从E-O边界开始,有1个长期的逐渐变冷过程(降低3~5℃),与同一站点孢粉记录的结果非常吻合,与北半球中纬度陆地气候重建的结果也是一致的[57]。基于采自刚果河羽状三角洲的沉积物柱状样中的支链GDGTs的分布,Weijers等重建了过去25 000年热带非洲大陆气温的变化,发现末次冰盛期到全新世气温升高4℃,高于热带大西洋SST同期的增温[56]。在1个岩芯中通过重建同时得到MA T和SST使得Weijers等能够估算过去25 000年的陆地-海洋温度梯度,并指出温度梯度的变化决定了降雨变化[56]。Rueda等将采自挪威南部Skagerrak湾近岸的沉积物同时分析了类异戊二烯和支链GDGTs,利用TEX86、B IT和MBT/CBT指标重建了过去200年来SST和MA T的变化并将两者进行了比较,发现SST和M A T分别与历史记录的年平均表层海水温度和夏季气温吻合良好[10]。以上研究证明,分析近海沉积物中的生物标志物能够同时重建SST和MA T,这将无疑有利于比较海洋和陆地古气候变化的相同性和差异,明确气候变化机制。
古海拔是古地理、古气候和古生态重建中的一个至关重要的因素,定量研究古海拔的变化对阐述诸如青藏高原抬升历史等重大科学问题具有十分重要的意义,但是古海拔的定量重建一直是当前古植物学与古环境研究领域中的难点[59]。应用GDGT分子指标重建陆地气温随海拔变化是估算古海拔的一种新尝试。Peterse等分析了采自中国贡嘎山东麓海拔1 180~3 819 m的表层(0~5 cm)土壤样品,分析了其支链GDGTs的分布,并计算了各样品代表的温度,结果表明海拔每升高1 000 m,气温降低5.9℃,与气象站记录的温度下降速率(-6.0℃/1 000 m)很接近,但是数据点比较分散(R2为0.55),显示目前利用MBT/CBT指标重建古海拔还有很大的不确定性[9]。对此,作者建议将MBT/CBT指标与最近提出的利用不同海拔降水δD差异估算海拔高度的土壤正构烷烃δD指标相结合[60],以期获得更可靠的古海拔估算结果。
尽管已经取得了这些令人鼓舞的结果,但是关于MBT/CB T指标的运用仍然存在许多问题需要解决。首先,虽然MBT/CBT指标目前已被应用于各种环境,但只在热带区域的应用比较成功。因为在这些区域,大量的支链GDGTs从陆地经河流持续稳定地输运到海洋[6,55,61]。与潮湿的热带地区相对稳定的环境条件不同,高纬度区域平均大气温度M A T低于0℃,陆地常年被冰雪覆盖,生长季节很短,土壤形成率低,陆地土壤有机质向海洋中输送的主要机制是冰期和春季雪融的时期,这可能会影响MBT/CBT指标的有效运用,因此它在高纬度环境中的适应性还有待研究[8]。其次,最近在海洋沉积物中发现了支链GDGTs的现场生产,虽然数量相对较低,但是土壤中的支链GDGTs和海洋中现场生产的支链GDGTs分布有何异同尚不清楚,因此海洋中现场生产的支链GDGTs是否会对陆地气候信号的解析产生影响、会在多大程度上产生影响都还不清楚[8]。与此类似,支链GDGTs在湖泊沉积物中也很丰富,原来认为可能主要来自土壤的侵蚀[7,36],但是最近Tierney等的研究表明,土壤样品和水生样品中,支链GDGTs的甲基化和环化程度有显著的差异,说明在湖泊水生系统中可能也存在支链GDGTs的现场生产,这也势必使得MBT/CBT指标在湖泊沉积物中的应用变得复杂化[62]。因此,这些新的有机地化指标在海洋和湖泊沉积物中的应用都仍然需要进一步的验证[1]。
我国陆地环境受冬季季风和夏季季风控制,从东南到西北温度和降雨都有很大的梯度。同时我国陆地上的黄土-古土壤堆积、洞穴碳酸钙(石笋)和湖泊沉积物保留了连续、高分辨率的古环境-古生态记录。利用多参数指标,我国陆地环境重建做出了世界一流的成果,尤其是有关季风变化和植被演变规律的研究[63-70]。我国陆地古环境-古生态研究遇到的1个瓶颈问题是区分降雨与温度的影响,因为目前用于黄土和石笋研究的指标(比如黄土的磁化率和石笋的氧同位素)主要受季风所带来的降雨影响,而温度的影响还未能得到充分的研究。这是因为目前还没有1个比较成熟的陆地温度重建指标,难以获得如SST般定量化的古温度信息。我国陆地拥有大量的湖泊,Chu等分析了我国湖泊沉积物中的长链烯酮,发现在部分湖泊中U37K指标也可以用来重建平均气温[71]。从国际研究的结果可以看出GDGTs在我国海洋沉积物的分布应该比长链烯酮更广泛,所以GDGTs(尤其是支链GDGTs)有望成为一个可靠的重建陆地古温度的指标。利用分子指标重建我国陆地古温度,并结合其它记录研究我国陆地气候和生态演变机制极具潜力。
3 结语
GDGTs已经在古海洋学、古湖沼学和古气候学研究中显示了独特的优势和巨大的应用潜力。基于类异戊二烯GDGTs的TEX86指标的应用使得重建海洋和湖泊长时间尺度表层水温、特别是地质历史高温时期的表层海水温度SST成为可能,进一步结合B IT指标,则有望在近海SST重建中发挥作用。而基于支链GDGTs的MBT/CBT指标则提供了一个崭新的从海洋看陆地的视角,可以通过对近海沉积物的研究来反演流域的陆地气温变化。基于GDGTs的有机地球化学指标在近海和陆地的同时应用为比较大气温度和海水温度变化的相同性和差异提供了方便。虽然在应用中目前仍然存在一些需要解决的问题,但已经取得的研究结果是令人鼓舞的。国内在GDGTs古温度重建等方面的应用目前鲜有报道,应当及时开展有关研究。
[1] Sinninghe Damsté J S,Ossebaar J,Abbas B,et al.Fluxes and distribution of tetraether lipids in an equatorial African lake:constraints on the application of the TEX86palaeothermometer and BIT index in lacustrine settings[J].Geochimica et Cosmochimica Acta,2009,73:4232-4249.
[2] Sinninghe Dam st?J S,Rijpstra W IC,Hopmans E C,et al.Distribution of membrane lipids of planktonic crenarchaeota in the A-rabian Sea[J].App lied Environmental Microbiology,2002,68:2997-3002.
[3] Schouten S,Hopmans E C,Baas M.Intact membrane lipids of“Candidatus Nitrosopumilus maritimus”a cultivated representative of the cosmopolitan mesophilic Group I Crenarchaeota[J].Appllied Environmental Microbiology,2008,74:2433-2440.
[4] Weijers JW H,Schouten S,Hopmans EC,et al.Membrane lipids of mesophilic anaerobic bacteria thriving in peats have typical archaeal traits[J].Environmental Microbiology,2006,8:648-657.
[5] Schouten S,Hopmans E C,SchefuβE,et al.Distributional variations in marine crenarchaeotal membrane lipids:a new tool fo r reconstructing ancient sea water temperatures[J]?Earth and Planetary Science Letters,2002,204:265-274.
[6] Hopmans E C,Weijers JW H,SchefuβE,et al.A novel proxy for terrestrial organic matter in sediments based on branched and isoprenoid tetraether lipids[J].Earth and Planetary Science Letters,2004,224:107-116.
[7] Powers L A,Werne J P,Johnson T C,et al.Crenarchaeotal lipids in lake sediments:A new paleotem perature proxy for continental paleoclimate reconstruction[J]?Geology,2004,32:613-616.
[8] Peterse F,Kim J H,Schouten S,et al.Constraints on the application of the MBT/CBT palaeothermometer in high latitude environments(Svalbard,Norway)[J].O rganic Geochemistry,2009,40:692-699.
[9] Peterse F,van der Meer MT J,Schouten S,et al.Assessment of soil n-alkaneδD and branched tetraether membrane lipid distributions as tools for paleoelevation reconstruction[J].Biogeosciences Discuss,2009,6:8609-8631.
[10] Rueda G,Rosell-MeléA,Escala M,et al.Comparison of instrumental and GDGT based estimatesof sea surface and air temperatures from the Skagerak[J].Organic Geochemistry,2009,40:287-291.
[11] Wuchter C,Schouten S,Wakeham S G,et al.Temporal and spatial variation in tetraether membrane lipids of marine Crenarchaeota in particulate organic matter:implications for TEX86paleothermometry[J].Paleoceanography,2005,20:PA 3013.
[12] Karner M B,DeLong E F,Karl D M.A rchaeal dominance in the mesopelagic zone of the Pacific Ocean[J].Nature,2001,409:507-510.
[13] Sinninghe DamstéJ S,Hopmans EC,Schouten S,et al.Crenarchaeol:the characteristic co re glycerol dibiphytanyl glycerol tetraether membrane lipid of cosmopolitan pelagic crenachaeota[J].Journal of Lipid Research,2002,43:1641-1651.
[14] 姚鹏,于志刚.海洋沉积物中现存微生物化学标志物完整极性膜脂研究进展[J].地球科学进展,2010,25(5):474-483.
[15] Schouten S,Forster A,Panoto F E,et al.Towards the calibration of the TEX86paleothermometer for tropical sea surface temperatures in ancient greenhouse worlds[J].Organic Geochemistry,2007,38:1537-1546.
[16] Schouten S,Hopmans E C,Forster A,et al.Extremely high sea-surface temperaturesat low latitudes during themiddle Cretaceous as revealed by archaeal membrane lipids[J].Geology,2003,31:1069-1072.
[17] Kim J H,Schouten S,Hopmans E C,et al.Global sediment co re-top calibration of the TEX86paleothermo meter in the ocean[J].Geochimica et Cosmochimica Acta,2008,72:1154-1173.
[18] 赵美训,李大伟,邢磊.古菌生物标志物古海水温度指标TEX86研究进展[J].海洋地质与第四纪地质,2009,29:75-84.
[19] Wuchter C,Schouten S,Coolen M J L,et al.Temperature-dependent variation in the distribution of tetraether membrane lipids of marine Crenarchaeota:implications for TEX86 paleothermometry[J].Paleoceanography,2004,19:PA4028-1-10.
[20] Trommer G,Siccha M,van der Meer MT J,et al.Distribution of Crenarchaeota tetraether membrane lipids in surface sediments from the Red Sea[J].O rganic Geochem istry,2009,40:724-731.
[21] Pearson A,Pi YD,Zhao W D,et al.Factors controlling the distribution of archaeal tetraethers in terrestrial hot springs[J].Appllied Environmental Microbiology,2008,74:3523-3532.
[22] Hofmann P,St sser I,Wagner T,et al.Climate-ocean coup ling off North-West Af rica during the Low er A lbian:The Oceanic Anoxic Event 1b[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2008,262:157-165.
[23] 张晓林,沙金庚,刘静.四醚膜类脂物的古温标—TEX86[J].古生物学报,2008,47:498-505.
[24] Schouten S,Hopmans E C,Sinninghe DamstéJ S.The effect of maturity and depositional redox conditions on archaeal tetraether lipid palaeothermometry[J].Organic Geochemistry,2004,35:567-571.
[25] Turich C,Freeman K F,Bruns M A,et al.Lipidsof marine Archaea:Patterns and provenance in the water-column and sediments[J].Geochimica et Cosmochimica Acta,2007,71:3272-3291.
[26] Anderson N J.Diatom s,temperature and climatic change[J].European Journal of Phycology,2000,35:307-314.
[27] 陈建徽,陈发虎,赵艳,等.古温度定量重建的良好代用指标—湖泊沉积摇蚊化石记录研究进展[J].地球科学进展,2004,19:782-788.
[28] 曾承,安芷生,刘卫国,等.湖泊碳酸盐记录的古水温定量研究进展[J].盐湖研究,2008,16:42-50.
[29] Zink K G,Leythaeuser D,Melkonian M,et al.Temperature dependency of long-chain alkenone distributions in Recent to fossil limnic sediments and in lake waters[J].Geochimica et Cosmochimica Acta,2001,65:253-265.
[30] Liu Z H,Henderson A C G,Huang Y S.Alkenone-based reconstruction of late-Holocene surface temperature and salinity changes in Lake Qinghai,China[J].Geophys Res Lett,2006,33,L09707,doi:10.1029/2006GL026151.
[31] Pearson E J,Juggins S,Farrimond P.Distribution and significance of long-chain alkenones as salinity and temperature indicato rs in Spanish saline lake sediments[J].Geochimica et Cosmochim ica Acta,2008,72:4035-4046.
[32] Escala M,Rosell-Mele A,Masque P.Rapid screening of glycerol dialkyl glycerol tetraethers in continental Eurasia samp les using HPLC/APCI-ion trap mass spectrometry[J].Organic Geochemistry,2007,38:161-164.
[33] Powers L A.Calibration and application of a new palaeotemperature tool in lacustrine systems:TEX86for continental palaeoclimate reconstruction.Doctor Dissertation[D].Twin Cities:University of M innesota,2005.
[34] Powers L A,Johnson T C,Werne J P,et al.Large temperature variability in the southern African tropics since the Last Glacial Maximum[J].Geophysical Research Letters,2005,32:L08706-1-4.
[35] Tierney J E,Russel J M,Huang Y S,et al.Northern hemisphere controls on tropical Southeast African climate during the past 60,000 years[J].Science,2008,322:252-255
[36] Blaga C,Reichart GJ,HeiriO,et al.Tetraether membrane lipid distributions in water-column particulate matter and sediments:A study of 47 European lakes along a North-South transect[J].Journal of Paleolimnology,2009,doi 10.1007/s10933-008-9242-2.
[37] Powers L,Werne J P,Vanderwoude A J,et al.Applicability and calibration of the TEX86paleothermometer in lakes[J].Organic Geochemistry,2010,41:404-413.
[38] Yu J Q,Zhang L.Lake Qinghai:Paleoenvironment And Paleoclimate[M].Beijing:Science Press,2008.
[39] Li J,Philip R P,Pu F,et al.Long chain alkenones in Qing hai Lake sediments[J].Geochimica et Cosmochimica Acta,1996,60:235-241.
[40] 付明义,刘卫国,李祥忠,等.青海湖及柴达木盆地地区现代湖泊沉积物中长链烯酮的分布特征[J].湖泊科学,2008,20:285-290.
[41] Sinninghe DamstéJ S,Hopmans EC,Pancost RD,et al.New ly discovered non-isop renoid dialkyl diglycerol tetraether lipids in sediments[J].Journal of the Chemical Society,Chemical Communications,2000:1683-1684.
[42] Weijers JW H,Schouten S,Spaargaren O C,et al.Occurrence and distribution of tetraether membrane in soils:Imp lications for the use of the B IT index and the TEX86SST p roxy[J].O rganic Geochemistry,2006,37:1680-1693.
[43] Lang worthy T A,Holzer G,Zeikus J G,et al.Iso-branched and anteiso-branched glycerol diethers of the thermophilic anaerobe Thermodesul fotobacterium commune[J].Systematic and Applied Microbiology,1983,4:1-17.
[44] Huber R,Wilharm T,Huber D,et al.Aquifex pyrophilus gen.nov.sp.nov.rep resents a novel group of marine hyper thermophilic hydrogen-oxidizing bacteria[J].Systematic and Applied Microbiology,1992,15:340-351.
[45] Sinninghe Damst?J S,Rijpstra W I,Hopmans EC,et al.Structural characterization of diabolic acid-based tetraester,tetraether and mixed ether/ester,membrane-spanning lipids of bacteria from the order[J]Ther motogales.Archives of Microbiology,2007,188:629-641.
[46] Schouten S,Hopmans E C,Pancost R D,Sinninghe Dam st?J S.Widesp read occurrence of structurally diverse tetraether membrane lipids:evidence for the ubiquitous presence of low-temperature relatives of hyperthermophiles[J].Proceedings of the National Academy of Sciences,USA,2000,97:14421-14426.
[47] Herfort L,Schouten S,Boon J P,et al.Characterization of transport and deposition of terrestrial organic matter in the Southern North Sea using the BIT index[J].Limnology and O-ceanography,2006,51:2196-2205.
[48] Kim J H,Schouten S,Buscail R,et al.Origin and distribution of terrestrial organic matter in the NW Mediterranean(Gulf of Lions):Exp lo ring the newly developed B IT index[J].Geochemistry,Geophysics,Geosystems,2006,7:1-20.
[49] Walsh EM,Ingalls A E,Keil R G.Sources and transport of terrestrial organic matter in Vancouver Island fjords and the Vancouver-Washington Margin:a multiproxy approach usingδ13Corg,lignin phenols,and the ether lipid BIT index[J].Limnology and Oceanography,2008,53:1054-1063.
[50] Huguet C,Smittenberg R H,Boer W,et al.Twentieth century proxy records of temperature and soil organic matter input in the Drammensfjord,southern Norway[J].Organic Geochemistry,2007,38:1838-1849.
[51] Kim J H,Ludwig W,Schouten S.Impact of flood events on the transport of terrestrial organic matter to the ocean:a study of the Tt River(SW France)using the BIT index[J].Organic Geochemistry,2007,38:1593-1606.
[52] Sluijs A,Schouten S,Pagani M,et al.Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum[J].Nature,2006,441:610-613.
[53] Mnot G,Bard E,Rostek F,et al.Early reactivation of European rivers during the last deglaciation[J].Science,2006,313:1623-1625.
[54] Weijers JW H,Schouten S,SchefuβE,et al.Disentangling marine,soil and plant organic carbon contributions to continental margin sediments:A multi-proxy approach in a 20,000 year sediment record from the Congo deep-sea fan[J].Geochimica et Cosmochimica Acta,2009,73:119-132.
[55] Weijers JW H,Schouten S,van den Donker JC,et al.Environmental controls on bacterial tetraether membrane lipid distribution in soils[J].Geochimica et Cosmochimica Acta,2007,71:703-713.
[56] Weijers JW H,SchefuβE,Schouten S,et al.Coupled thermal and hydrological evolution of tropical Africa over the last deglaciation[J].Science,2007,315:1701-1704.
[57] Schouten S,Eldrett J,Greenwood D R,et al.Onset of longterm cooling of Greenland near the Eocene-Oligocene boundary as revealed by branched tetraether lipids[J].Geology,2008,36:147-150.
[58] Liu Z H,Pagani M,Zinniker D,et al.Global cooling during the Eocene-Oligocene climate transition[J].Science,2009,323:1187-1190.
[59] 孙启高.如何利用化石植物定量研究古海拔[J]?地质论评,2008,54:145-154.
[60] Jia GD,Wei K,Chen F J,et al.Soil n-alkaneδD vs.altitude gradients along Mount Gongga,China[J].Geochimica et Cosmochimica Acta,2008,72:5165-5174.
[61] Schouten S,Ossebaar J,Brummer G J,et al.Transport of terrestrial organic matter to the deep North Atlantic Ocean by ice rafting[J].Organic Geochemistry,2007,38:1161-1168.
[62] Tierney J E,Russel J M.Distributions of branched GDGTs in a tropical lake system:implications for lacustrine application of the MBT/CBT paleoproxy[J].Organic Geochemistry,2009,40:1032-1036.
[63] Guo Z T,Ruddiman W M,Hao Q Z.Onset of Asian desertification by 22 M yr ago inferred fromloess deposits in China[J].Nature,2002,416:159-163.
[64] Wang Y J,Cheng H,Edwards R L,et al.The Holocene Asian monsoon:Links to solar changes and North Atlantic climate[J].Science,2005,308:854-857.
[65] Wang Y J,Cheng H,Edwards R L,et al.Millennial-and orbit-al-scale changes in the East Asianmonsoon over the past 224,000 years[J].Nature,2008,451:1090-1093.
[66] Sun Y B,Clemens S C,An Z S,et al.Astronomical timescale and palaeoclimatic implication of stacked 3.6-M yr monsoon records from the Chinese Loess Plateau[J].Quaternary Science Review s,2006,25:33-48.
[67] Chen X L,Fang XM,An Z S,et al.An 8.1 Ma calcite record of Asian summer monsoon evolution on the Chinese central Loess Plateau[J].Science in China:Series D,2007,50:392-403.
[68] 刘敬华,张平中,程海,等.黄土高原西缘在AD 1875~2003期间石笋氧同位素记录的季风降水变化与海气系统的联系[J].科学通报,2008,53:2801-2808.
[69] 吴江滢,汪永进,程海,等.葫芦洞石笋记录的19.9~17.1 ka BP东亚夏季风增强事件[J].中国科学(D辑),2009,39:61-69.
[70] 周斌,沈承德,郑洪波,等.黄土高原中部晚第四纪以来植被演化的元素碳碳同位素记录[J].科学通报,2009,54:1262-1268.
[71] Chu G Q,Sun Q,Li SQ,et al.Long-chain alkenone distributions and temperature dependence in lacustrine surface sediments from China[J].Geochimica et Cosmochimica Acta,2005,69:4985-5003.
猜你喜欢
上海化工(2022年3期)2022-06-30
卫星应用(2022年3期)2022-05-23
农业机械学报(2021年10期)2021-11-09
中国化工贸易·上旬刊(2020年5期)2020-09-10
科学导报·学术(2020年12期)2020-04-14
小哥白尼(军事科学)(2019年7期)2019-11-16
分析化学(2015年3期)2015-04-20
北京航空航天大学学报(2014年1期)2014-12-19
中国机械工程(2012年15期)2012-07-25
中学生数理化·高二版(2008年2期)2008-10-19
