
运动、补糖或刺五加对大鼠糖原耗竭运动后肌细胞膜GLUT4转位的影响
2011-01-03 10:22杨则宜
天津体育学院学报 2011年4期
周 亮,杨则宜
●博士(生)论坛
运动、补糖或刺五加对大鼠糖原耗竭运动后肌细胞膜GLUT4转位的影响
周 亮1,杨则宜2
目的:研究大鼠运动后骨骼肌细胞膜葡萄糖转运体4(Glucose transporter 4,GLUT4)转位及其时相性变化,并探索补糖和刺五加对其影响。方法:128只SD大鼠大鼠随机分为训练对照、训练补糖、训练补刺五加皂甙和训练补刺五加皂甙和糖4组,在糖原消耗性运动前和运动后不同时间点(0 h,4 h,12 h)采样,共16小组(n=8)。采用Western blotting方法分析骨骼肌细胞质膜和细胞膜的GLUT4的相对蛋白含量。结果:(1)糖原耗竭性运动后骨骼肌细胞内质膜GLUT4相对蛋白量明显降低(105.66±10.54 vs 98.05±11.89)。补充刺五加提高了骨骼肌的骨骼肌细胞内质膜GLUT4蛋白含量。(2)糖原耗竭性运动后即刻骨骼肌细胞膜的GLUT4相对蛋白量明显升高(100.47±10.40 vs 188.14±24.31)。补糖和刺五加可显著升高运动后骨骼肌细胞膜GLUT4相对蛋白含量。结论:运动后骨骼肌细胞膜GLUT4转位增加,补糖或补刺五加均可以促使运动后GLUT4转位增加。
运动;GLUT4;补糖;刺五加;细胞膜
近来研究发现,肌糖原合成的主要限速反应是葡萄糖进入骨骼肌的过程。葡萄糖进入肌细胞不是简单的单纯扩散,而是借助于肌细胞膜表面葡萄糖载体蛋白(Glucose transporter,GLUT)的异化扩散过程。在骨骼肌组织中,葡萄糖转运的激活依赖含GLUT4的小泡从运动敏感的细胞内向浆膜和T管直接动员[1]。引起骨骼肌GLUT4转位的分子机制非常复杂[2]。现在已经很清楚骨骼肌内分别存在对胰岛素和运动做出反应的不同的“GUT4池”。胰岛素和运动诱导GLUT4转位并增加骨骼肌葡萄糖摄取的分子信号是不同的:胰岛素引起rab4蛋白分布的变化,而运动引起转铁蛋白受体的从新分布[3]。尽管这样,但是二者之间还是有相似性,运动和胰岛素都能引起胰岛素应答氨基肽酶和肌膜上GTP接合蛋白的浓度变化[4]。大量研究显示,运动后及时补糖是促进运动后糖原合成的重要营养学手段。同时,也有研究表明,运动后补充刺五加会促进骨骼肌的糖原合成。但是,其机制尚不明确,运动后补糖或刺五加对骨骼肌细胞的GLUT4转位有何影响尚未见报道。
1 研究对象与方法
1.1 实验动物与饲养条件
SPF/VAF级雄性SD大鼠,体重180~210 g,购自北京维通利华实验动物技术有限公司。质量合格证号为:NO.0049137。国家甲级动物饲养房内随机分笼饲养,每笼8只,自由进食和饮水。饲养房室温20~25℃,湿度40%~70%。每日光照时间为8∶00-20∶00。
1.2 实验动物筛选
实验大鼠共购买150只,所有大鼠安静饲养3天,第4天开始进行为期3天的游泳技能学习。第3天的游泳训练视为筛选测试。如果游泳过程中大鼠下沉超过10 s,捞出休息60 s,再次放入水中。如果连续3次均下沉超过10 s,则视为游泳能力太差而遭淘汰,经实验选择128只用于正式实验。
1.3 实验动物分组
128只大鼠,随机分为训练对照组(C组)、训练补糖组(G组)、训练补刺五加皂甙组(A组)和训练补糖加刺五加皂甙组(GA组)4组,各大组按照最后一天的糖原消耗性运动前和运动后不同的标本采集时间(0 h、4 h、12 h)又各分4小组,共16小组(n=8)。
1.4 实验动物训练方案
每日进行静水水池中负重递增的间歇高强度游泳训练和流动水池中时间递增的耐力游泳训练,每3天一个周期,共2个周期,第7天休息,第8天进行糖原消耗性运动。负重为尾根部系重物法,每3天测量一次体重以调整负重(见表1)。

表1 实验动物训练方案Tab.1 Project of animals training
1.5 消耗糖原运动方案
糖原消耗性运动参照魏守刚的方案修改[5],设计为流动水中进行间歇高强度负重游泳训练和耐力游泳训练结合。方法为:大鼠负重9%,流动水池游泳60 s,休息120 s,如此重复3次后,除去负重物,再将其放入上述流动水池中游泳90 min(见表1)。
1.6 药物补给方案
灌胃,每日训练后的即刻进行,灌胃量为2 mL/只。休息日在上午8:00—9:00点给药。刺五加皂甙的剂量为每天500 mg/ kg,补蔗糖的剂量为每天6 g/kg,同时补糖和补刺五加组剂量同上,按剂量要求和灌胃量配成溶液,对照组给予等量的蒸馏水。刺五加皂甙购自浙江霍夫曼德公司,食品级蔗糖购自超市。
1.7 标本采集
各组大鼠在相应时点宰杀,2%戊巴比妥钠(5 mg/100 g体重)腹腔麻醉后,取双侧股四头肌,液氮速冻后-80℃低温冰箱保存待测。
1.8 GLUT4检测方法[6]
1.8.1 大鼠骨骼肌细胞内质膜和外膜的分离 用改良Klips方法制备大鼠骨骼肌细胞内质膜和外膜。取大鼠后肢股四头肌,肌肉约为10 g,浸至0℃溶液A(250 mmol/L Sucrose,50 mmol/L Tris,0.2 mmol/L EDTA)中,剔除血管、脂肪及筋膜后,剪碎,匀浆。匀浆液以9 000 g高速离心10 min(4℃),保留上清,沉淀如上再匀浆、离心,共3次。3次上清以190 000 g超离心1 h(4℃),取沉淀铺于25%、30%、35%的蔗糖梯度上,150 000 g超离心16 h(4℃)。于25%蔗糖层得到骨骼肌细胞膜悬浊液,35%层上得到骨骼肌细胞内质膜悬浊液。膜蛋白浓度用蛋白定量试剂盒测定。
1.8.2 GLUT4的检测 取大鼠骨骼肌细胞内质膜和外膜各50μg,用缓冲液A稀至相同体积,加入等体积的2倍样品溶液〔0.1 mol/L Tris -HCL,PH6.8;Glycerol 20% (V/V);SDS 2% ;Bromophenolblue 0.005%;2-Mercaptoethanol10%(V/V)〕,以 9 000 g离心10 min,取相同体积上清液在10%聚丙烯酰胺的胶上进行SDS凝胶电泳,随后转移至硝酸纤维素(NC)膜上,NC膜浸于含5%脱脂奶粉的PBS中,在水浴摇床中温育60 min(37℃),然后加入GLUT4羧基端12肽的多克隆抗体过夜(4℃)。洗涤后与HRP标记的羊抗兔IgG水浴摇床温育1 h(37℃),充分洗涤后与ECL(增强化学发光剂,Amersham公司生产)反应1 min,即刻用Kodak底片爆光,洗片后进行扫描,定量。以对照组内膜的GLUT4蛋白含量为100作为标准,计算其他组的相对含量。
1.9 统计学分析
数据用平均数±标准差表示,统计处理采用Spss12.0软件完成,用SigmaPlot软件做图。采用广义线性模型(General Liner Model)进行单变量分析(Univaridfe),分析补糖、补药和运动3个因素对某一变量的主效应,两两之间交互效应及三者间的交互作用;采用单因素方差分析(One-Way ANOVA)由LSD多重比较同一时刻点C、G、A和GA各组间的AMPK蛋白量的差异。
2 实验结果
2.1 股四头肌骨骼肌细胞内质膜GLUT4相对蛋白量
2.1.1 各因素对骨骼肌细胞内质膜GLUT4蛋白量的主效应及其交互作用 方差分析表明,补药和运动对骨骼肌细胞内质膜GLUT4相对蛋白量的主效应有统计学显著意义,但是他们之间没有交互作用。补糖对骨骼肌细胞内质膜GLUT4相对蛋白量的主效应不明显(F=0.602,Sig=0.439),补糖和补药之间也没有明显的交互作用(F=3.391,Sig=0.068)。补药对骨骼肌细胞内质膜GLUT4相对蛋白量的变化影响最大(补药大鼠的骨骼肌细胞内质膜GLUT4相对蛋白量均数为105.93,不补药大鼠为94.31)。运动的影响主要体现为运动前骨骼肌细胞内质膜GLUT4相对蛋白量较高,糖原耗竭性运动后即刻明显降低,而且在糖原耗竭性运动后4 h内持续降低,随后有所恢复,在糖原耗竭性运动后12 h有所上升,但是并没有达到运动前水平(见图1)。
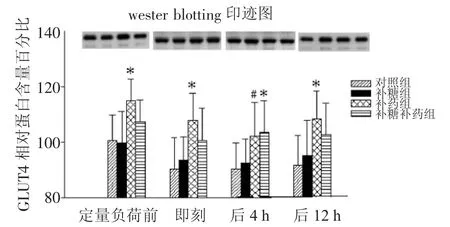
图1 各组大鼠运动后骨骼肌细胞内质膜GLUT4相对蛋白含量的变化Fig.1 Change of the relative quantity of GLUT4 on intracellular membrane in different groups of rats after exercise
2.1.2 各因素对骨骼肌细胞内质膜GLUT4相对蛋白量的影响安静状态时、糖原耗竭性运动后即刻和12 h,补药组的骨骼肌细胞内质膜GLUT4蛋白量相对较高,和对照组及补糖组相比较,差异有显著性。在糖原耗竭性运动后4 h,补药组和补糖补药组的骨骼肌细胞内质膜GLUT4相对蛋白量含量均较高,补药组相对对照组差异有显著性,而补药同时补糖组相对于对照组和单纯补糖组差异有显著性(见表2、表3)。
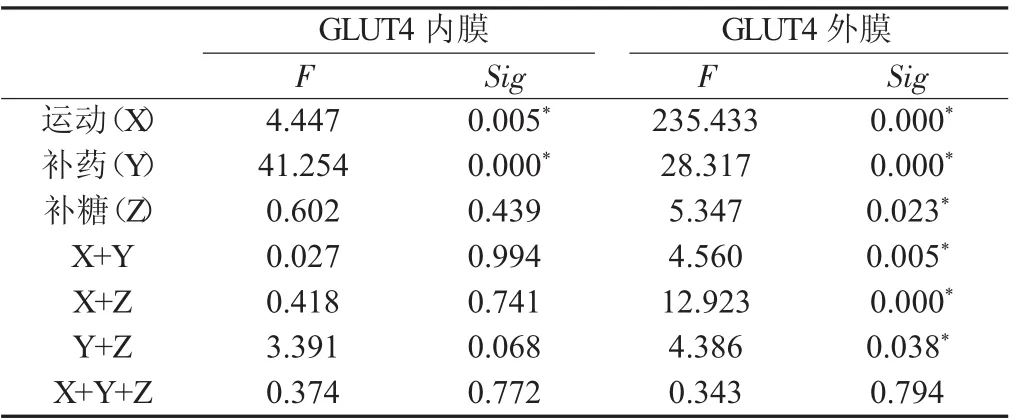
表2 运动、补糖和补药对各变量的主效应(Ⅳ)Tab.2 Main effects of the time course after exercise,glucose or Acanthopanax administration on the different variable(Ⅳ)

表3 各组大鼠骨骼肌细胞内质膜GLUT4相对蛋白含量比较Tab.3 Compare of the relative quantity of GLUT4 in intracellular membrane in different groups of rats
2.2 股四头肌细胞膜GLUT4蛋白量
2.2.1 各因素对骨骼肌细胞膜GLUT4蛋白量的主效应及其交互作用 方差分析表明,补糖、补药和运动对骨骼肌细胞膜GLUT4相对蛋白量的主效应都有统计学显著意义,并且两两间的交互作用明显。补糖组骨骼肌细胞膜GLUT4相对蛋白含量为(149.10±42.09),不补糖大鼠为(143.45±31.22)。补药对骨骼肌细胞膜GLUT4相对蛋白量的变化影响最大补药大鼠的骨骼肌细胞膜GLUT4相对蛋白量均数为152.78±39.94,不补药大鼠为(139.77±32.89)。运动的影响主要体现为运动前骨骼肌细胞膜GLUT4相对蛋白量较低(100.47±10.40),运动后即刻最高(188.14±24.31),在随后的恢复过程中逐渐降低(见图2)。
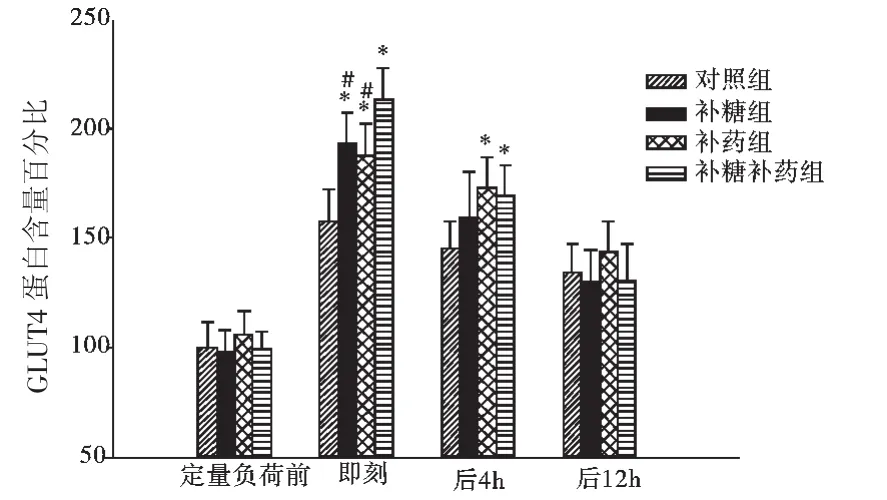
图2 各组大鼠运动后骨骼肌细胞膜GLUT4相对蛋白含量的变化Fig.2 Change of the relative quantity of GLUT4 in extracellular membrane in different groups of rats after exercise
2.2.2 各因素对骨骼肌细胞膜GLUT4相对蛋白量的影响 方差分析表明,在运动前安静状态时,各组的骨骼肌细胞膜GLUT4蛋白量基本一致,没有统计学显著性差异。在运动后即刻,各组的骨骼肌细胞膜GLUT4相对蛋白含量升高,组间也体现了显著性差异,表现为对照组相对其他各组的骨骼肌细胞膜GLUT4相对蛋白含量较低,补糖同时补药最高,相对其他各组都有统计学显著性意义。在随后的恢复过程中,各组大鼠的骨骼肌细胞膜GLUT4相对蛋白含量均降低,直至12 h依然高于安静状态时水平。在糖原耗竭性运动后4小时时刻点,体现为补药组和补药同时补糖组比其他两组较高,有统计学意义(见表4)。
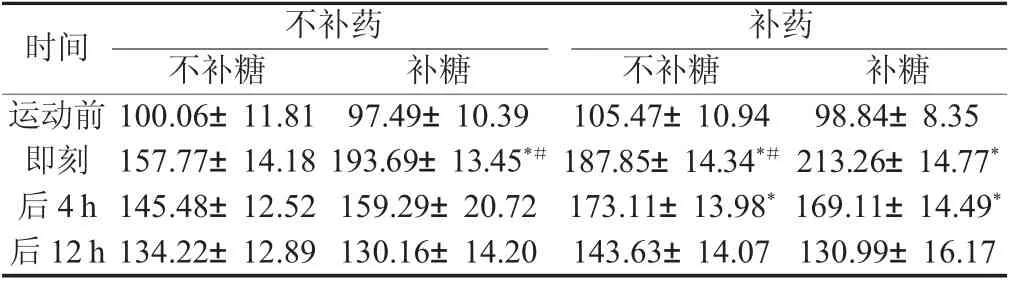
表4 各组大鼠骨骼肌细胞膜GLUT4相对蛋白含量比较Tab.4 Compare of the relative quantity of GLUT4 in extracellular membrane in different groups of rats
3 讨 论
血糖是运动后糖原恢复的重要底物来源,血糖进入肌细胞主要依赖葡萄糖载体的转运,GLUT4是存在肌细胞膜表面的主要葡萄糖转运体。运动是一种可调节骨骼肌细胞内GLUT4的生理因素,单纯的急性运动或电刺激离体的骨骼肌就能直接促进骨骼肌细胞内的GLUT4向骨骼肌细胞膜转位,并使其内在活性增加[7]。长期的体育锻炼可以提高骨骼肌细胞GLUT4的含量,这种升高与在分离的骨骼肌中观察到的胰岛素刺激下葡萄糖摄人升高以及胰岛素敏感性增加相符。运动可改善外周组织对葡萄糖的摄取和利用,不同方式的运动后,正常大鼠骨骼肌GLUT4蛋白含量上升30%~200%不等[7]。去神经骨骼肌收缩研究进一步证实,收缩运动是骨骼肌GLUT4基因表达的一种调节因素。Friedman等在对肥胖大鼠进行跑台运动训练后发现,运动训练可刺激骨骼肌细胞GLUT4蛋白含量的增加,从而提高骨骼肌细胞对胰岛素的敏感性[8]。Eriksson等研究了7例糖尿病患者,他们参加3个月的抗阻运动后胰岛素敏感性增加23%,无氧葡萄糖代谢增加27%[9]。
杨晓冰等研究发现,经过6周游泳训练的大鼠,与对照组大鼠相比,骨骼肌细胞内质膜GLUT4含量增加16.0%(P<0.01),骨骼肌细胞膜GLUT4含量增加71%(P<0.01),表明运动可促进骨骼肌细胞内质膜GLUT4向骨骼肌细胞膜转运,从而提高细胞对葡萄糖的摄取和利用[10]。另外,糖尿病运动组大鼠骨骼肌细胞内GLUT4 mRNA含量明显高于糖尿病非运动组大鼠,提示细胞内GLUT4基因转录增加,进一步导致细胞内GLUT4含量增加,这可能是运动改善糖尿病机体外周组织对胰岛素的敏感性,促进葡萄糖利用的机理之一。
Ren等研究表明,经过一次耐力训练大鼠骨骼肌中GLUT4 mRNA量增加约2倍,GLUT4增加50%,经过两次耐力训练(每天一次)GLUT4增加约2倍,而GLUT4 mRNA量未再增加,在胰岛素刺激情况下细胞膜上GLUT4含量较正常组高2倍[11]。Ren还观察到正常大鼠运动3 h后,骨骼肌细胞内GLUT4 mRNA和蛋白含量就有所增加,而且GLUT4蛋白含量的增加可持续到停止运动1周后,GLUT4含量的增加和骨骼肌对胰岛素敏感性的改变在时间上和程度上相吻合。Cox等也研究了短期运动训练对老年人骨骼肌细胞GLUT4的影响,经过7天的耐力运动,老年人骨骼肌细胞GLUT4的含量明显增加,同时胰岛素活性也明显增加[12]。Martineau等研究表明,运动训练不但能够增加老年大鼠骨骼肌细胞内的GLUT4含量,而且可使老年大鼠心肌细胞内GLUT4含量明显增加,促进骨骼肌细胞和心肌细胞对葡萄糖的摄取和利用,改善葡萄糖耐量[13]。
和运动刺激GLUT4转位一样,胰岛素也可以在大鼠的骨骼肌中引起同样的生物学效应。已经有一些研究清楚的显示,在细胞内存在着分别对运动和胰岛素敏感的两种不同的GLUT4池,运动和胰岛素通过不同途径作用于这两种特定的GLUT4池。
Hayashi等研究发现,运动对胰岛素和GLUT4的激活机制不同[14]。胰岛素是利用磷脂酰肌醇-3-激酶依赖机制,而运动信号由肌浆网的钙离子释放,导致其他中介信号激活,此外也与自分泌或旁分泌介导的运输激活有关。运动后胰岛素相关的葡萄糖摄取显著增加,这种骨骼肌运动时胰岛素依赖性葡萄糖摄取增加有重要的临床意义,特别是对于有胰岛素抵抗的2型糖尿病患者。
骨骼肌细胞内质膜是细胞器膜和核膜的统称,其作用是把原生质分成既互相隔离、互不干扰,又互相关联、彼此依存的多相微环境。由于内膜和外膜的密度不同,内膜的沉降系数大于外膜,可通过梯度离心方法将其分离。在静息状态下,绝大部分GLUT4蛋白位于骨骼肌细胞内质膜中,如微粒体、高尔基体及其他对胰岛素敏感的富含GLur4的囊泡样膜结构,只有2%位于骨骼肌细胞膜上。当胰岛素与受体相结合后,触发细胞的排粒过程,GLUT4从细胞内向骨骼肌细胞膜移动并与骨骼肌细胞膜融合,转位于骨骼肌细胞膜上。发挥其转运葡萄糖的生理作用。在图1中,各组大鼠GLUT4相对蛋白含量变化的结果均分别来自4次独立Western Blot实验,实验中每次Western Blot是上样9孔,因为实验的western比较多,所以提取图样的时候只是提取了每个时间点的4个不同组做对照的在论文中体现,而且Western Blot基本也是半定量的研究,所以采用了相对对比,实验设计尽可能的合理、实验条件控制尽量一致,以减少误差。我们的实验结果发现,在运动后肌骨骼肌细胞内质膜的GLUT4蛋白含量有所下降(总体约下降7%)而骨骼肌细胞膜的蛋白量有所增加(总体上升约87%),这和以上的大部分结果是一致的。进一步的分析发现,运动后骨骼肌细胞内质膜的GLUT4含量下降,而其最低值并非是运动后即刻出现,而是在运动后的4 h,而后有所回升。因为我们的时间点是有限的并不能就此推断运动后GLUT4的变化规律是运动后哪个时点最低,但是至少可以说明在运动后的初期,肌骨骼肌细胞内质膜GLUT4含量变化较小,或者还稍减少(变化不到1%)。但是,我们对肌骨骼肌细胞膜GLUT4蛋白量的观察,发现其最高点是出现在运动后的即刻,随后下降,而不是在运动后初期稳定或稍上升。这可能是外膜GLUT4在此过程有所降解或内陷所致。
我们的实验显示,补糖对安静状态时肌骨骼肌细胞内质膜的GLUT4含量没有太大的影响,而补药则显著提高了肌骨骼肌细胞内质膜的蛋白含量。各组间安静时的肌骨骼肌细胞膜GLUT4蛋白含量的差异不大,说明补药提高了机体的总的GLUT4蛋白含量,这可能是因为补药后促进了肌细胞的GLUT4的mRNA表达和翻译所致。通过比较,我们还发现,补药组大鼠在运动后即刻肌骨骼肌细胞内质膜的GLUT4蛋白含量仅仅下降了6%,而外膜GLUT4的增加则达到约80%;对照组在运动后即刻肌骨骼肌细胞内质膜的GLUT4蛋白含量下降了10%,而外膜GLUT4的增加则仅仅约57%。说明补药后不仅增加了GLUT4的转位,同时也提高了肌细胞的GLUT4蛋白总量。
补糖组也提高了机体的GLUT4转位。因为有研究显示,胰岛素是GLUT4转位的强刺激因子。我们已经发现补糖组的胰岛素水平显著高于其他组,可能这是其增加GLUT4转位的重要原因之一。而补药对胰岛素的影响不大,甚至稍降低了胰岛素浓度,所以补药肯定并非通过胰岛素刺激GLUT4的转位。而且补药和补糖有明显的协同效应,说明在刺激GLUT4的转位和蛋白量的表达上存在着不同的机制。补药可能主要刺激GLUT4的转录、翻译,增加其蛋白总量;而补糖可能主要刺激其由内膜向外膜的转位,但是对其蛋白的表达影响不大。
4 结论
运动后GLUT4转位增加,补糖或补刺五加都可以促进这种转位,两者同时补充对肌骨骼肌细胞膜的GLUT4含量有协同作用。
[1]刘传道,江钟立.GLUT4转位和活性的研究进展[J].中国康复医学杂志,2004,19(4):313-315.
[2]Lemieux K,Han X X,Dombrowski L,et al.The transferrin receptor defines two distinct contraction-responsive GLUT4 vesicle populations in skeletal muscle[J].Diabetes,2000,49(2):183-189.
[3]Wright D C,Hucker K A,Holloszy J O,et al.Ca2+and AMPK both mediate stimulation of glucosetransport by muscle contractions[J].Diabetes,2004,53(2):330-335.
[4]Coderre L,Kandror KV,Vallega G,et al.Identification and characterization of an exercisesensitive pool of glucose transporters in skeletal muscle[J].J Biol Chem,1995,270(46):27 584-27 588.
[5]魏守刚,杨则宜,贾弘堤,等.运动训练和营养补剂对大鼠骨骼肌糖原生成素基因表达的影响[J].中国运动医学杂志,2005,24(2):143-146.
[6]Douen A G T,Ramlal S,Rastogi P J,et al.Exercise induces recruitment of the"insulin-responsive glucose transporter"Evidence for distinct intracellular insulin-and exercise recruitable transporter pools in skeletal muscle[J].J Biol Chem,1990,265(23):13 427-13 430.
[7]Rodnick K J,Henriksen E J,James DE,et al.Exercise training glucose transporter and glucose transport in rat skeletal muscle[J].Am J Physiol,1992,262(1):c9-cl4.
[8]杨晓冰,吴毅,李云霞,等.运动对糖尿病大鼠骨骼肌细胞葡萄糖运载体4的影响[J].中华内分泌代谢杂志,1998,14(6):390-391.
[9]Eriksson J,Tuominen J,Valle T,et al.Aerobic endurance exercise or circuit-type resistance training for individuals with impaired glucose tolerance[J].Horm Metab Res,1998,30(1):37-4.
[10]杨晓冰,吴毅,李益明,等.运动促进大鼠骨骼肌细胞葡萄糖运载体4的转位[J].中国运动医学杂志,2000,19(1):37-38.
[11]Ren J M,Semenkovich E A,Gulve J,et al.Exercise induces rapid increasesin GLUT4 expression,glucose transportcapacity,and insulin-stimulated glycogen storage in muscle[J].J Biol Chem,1994,269(20):14 396-14 401.
[12]CoxJ H,Cortright R N,DohmGL,et al.Effect ofagingon response the exercise training in humans:skeletal muscle GLUT4 and insulin sensitivity[J].J Appl Physiol,1999,86(6):2 019-2 025.
[13]Martineau L C,Chadan S G,Parkhouse W S.Resistance of the ages myocardium to exercise-induced chronic changes in glucose transport related protein content[J].Mech Ageing Dev,1999,110(1):109-118.
[14]HayashiT,HirshmanMF,KurthEJ,etal.Evidencefor5'-AM-activated protein kinase mediation of the efect of muscle contraction on glucose transport[J].Diabetes,1998,47(8):1 369.
Influence of Exercise and Glucose or Acanthopanacis Senticosi Supplement on GLUT4 Transposition in Muscle Cell after Glycogen Exhaust Exercise in Rats
ZHOU Liang1,YANG Zeyi2
(1.Dept.of PE,Hunan University of Science and Technology,Xiangtan 100084,China;2.China Antidoping Center,Beijing 100029,China)
Object:The purpose of this research was to explore the transposition of glucose transport 4(GLUT4)and the recovery in different phases after exercise in muscle cell in rat.Moreover,we should to study the effect of glucose and/or acanthopanacis senticosi supplement on the GLUT4 transposition. Method:128 rats were divided into four groups including exercise control groups(C groups),exercise and glucose administration groups(G groups),exercise and acanthopanacis senticosi administration groups(A groups),exercise and glucose and acanthopanacis senticosi administration groups(GA groups).The glucose and acanthopanacis senticosi supplement should be completed by intragastric adminis tration in 0.5 h after exercise.They were divided into 16 groups(n=8).the value of GLUT4 of intracellular membrane and all rats should be killed in different designing time points before or after glycogen exhaust exercise(0 h,4 h and 12 h,respectively).Then,the content of GLUT4 protein in membrane of myocyte and endoplasm were analyzed by Western blotting.Results:The quantity of GLUT4 in intracellular membrane of myocyte was highest before exercise(105.66±10.54)and obviously declined after exercise(98.05±11.89),this trend lasted to 4 h after exercise(97.22±11.50)then gradually recovered to the level at rest.The quantity of GLUT4 in endoplasm membrane of A groups was significance higher than that of C groups and G groups.The quantity of GLUT4 in myocyte membrane was lower before exercise(100.47±10.40)and attained the peak(188.14±24.31)instantly after exercise.The quantity of GLUT4 of C groups(134.38±25.03)was lower than that of any other groups.The value of GA groups was significantly higher than the others.Conclusions:The transfer of muscle cell membrane glucose transport 4 was improved after exercise.Both glucose and acanthopanacis senticosi administration could encourage those effect.
exercise;GLUT4;glucose administration;acanthopanacis senticosi;cellular membrane
G 804.7
A
1005-0000(2011)04-0341-05
2011-01-18;
2011-06-15;录用日期:2011-06-20
周 亮(1975-),男,湖南岳阳人,博士,副教授,研究方向为运动营养和运动机能评定。
1.湖南科技大学体育学院,湖南湘潭411201;2.国家体育总局运动营养研究所,北京100029。
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
医学综述(2021年16期)2021-12-01
爱你·心灵读本(2018年5期)2018-08-04
运动(2018年14期)2018-07-16
爱你(2018年13期)2018-06-07
体育科学(2018年3期)2018-04-20
分析化学(2018年11期)2018-01-16
课程教育研究·新教师教学(2016年14期)2017-04-12
大众健康(2015年12期)2015-09-10
科学中国人(2015年13期)2015-02-28
